大曲絲狀真菌的物種多樣性及其次級代謝產物的合成潛力
2019-08-26 03:24:50方程杜海徐巖
食品與發酵工業 2019年15期
關鍵詞:物種
方程,杜海,徐巖*
1(江南大學 生物工程學院,釀酒科學與酶技術中心,江蘇 無錫,214122) 2(工業生物技術教育部重點實驗室(江南大學),江蘇 無錫,214122)
大曲是中國白酒發酵所特有的糖化劑和發酵劑,它是在開放式環境中,通過自發的傳統固態發酵生產而來,并且其生產原料通常是未經滅菌處理的小麥、大麥和/或豌豆混合物[1],因此大曲中包含多種類型的微生物,如細菌、酵母和絲狀真菌[1-2]。其中絲狀真菌對白酒的發酵至關重要,因為絲狀真菌能夠分泌大量的水解酶類,可以將原料中的大分子物質水解成小分子的肽類、單糖等,從而為后續的酒精發酵提供能量物質[3]。
從分類學上來看,絲狀真菌是一類多樣性非常復雜的真核微生物,并且許多絲狀真菌的生長條件尚不明確,通過傳統培養的方法很難全面了解大曲中絲狀真菌的物種多樣性,因此需要借助未培養的方法。近年來興起的高通量測序技術可以對數百萬個DNA分子進行快速、準確地測序,使得更加深入研究微生物的多樣性成為可能[4],因此該技術也迅速在食品微生物領域得到廣泛的應用。例如,喬曉梅等[5]利用高通量測序對清香大曲真菌群落結構研究發現,在微生物物種水平上,中溫大曲包含米根霉(Rhizopusoryzae)、淀粉絲菌(Amylomycesrouxii)、微小根毛霉(Rhizomucorpusillus)、橫梗霉(Lichtheimia)和傘枝犁頭霉(Lichtheimiacorymbifera)、傘形卷霉等多種絲狀真菌[5]。夏玙等[6]同樣利用高通量測序發現,中高溫大曲中的優勢絲狀真菌屬主要為嗜熱子囊菌屬(Thermoascus)、根毛霉屬(Rhizomucor)和曲霉菌屬(Aspergillus)3種。除此以外,還觀察到了一些低豐度的Byssochlamys、Rhizopus、Monascus、Penicillium、Lichtheimia、Rasamsonia等絲狀真菌屬。最近,筆者通過高通量測序技術結合微生物溯源進行研究還發現,大曲中的部分絲狀真菌,如A.flavus和A.oryzae等,還能進入到白酒的酒精發酵過程[7-8],說明絲狀真菌除了產生各種水解酶類,可能還具有其他功能。
最近的研究表明,絲狀真菌具有強大的次級代謝產物合成潛力,并且許多次級代謝產物具有重要的藥理學活性[9],因而近年來許多研究者試圖從一些絲狀真菌豐富的棲息地(深海[10]、植物內生真菌[11]和海綿等[12])中研究這類微生物的物種多樣性,并挖掘具有生物活性的天然產物。且大曲中含有豐富的絲狀真菌資源,這些絲狀真菌可能也具有合成多種天然產物的潛力。但是,目前系統性地研究大曲中絲狀真菌的物種多樣性及其次級代謝產物的合成潛力較少。
本研究的目的有2個:(1)利用高通量測序的方法比較中溫、中高溫和高溫3種大曲中絲狀真菌的菌群結構差異;(2)結合純培養的方法研究大曲中絲狀真菌的物種多樣性及其次級代謝產物合成潛力。本研究為大曲來源的絲狀真菌次級代謝產物合成潛力研究奠定理論基礎,為與大曲相關的研究提供新的思路。
1 材料與方法
1.1 材料與儀器
1.1.1 大曲樣本采集
本研究使用的大曲樣本均為成品曲粉(表1),取樣時,每種樣本均使用無菌自封袋取500 g,并立即存放于-80 ℃,保存備用。

表1 本研究所使用的大曲樣本
1.1.2 試劑與設備
麥芽提取物瓊脂(malt extract agar,MEA)、馬鈴薯葡萄糖瓊脂(potato dextrose agar,PDA)、玉米粉瓊脂(corn meal agar,CMA)、沙氏葡萄糖瓊脂(sabouraud dextrose agar,SDA)和察氏瓊脂(czapek dox agar,CDA),青島海博生物技術有限公司;基因組DNA提取試劑盒(DP305)、1×TaqPCR Mastermix、DNA純化試劑盒(DP204)、瓊脂糖凝膠DNA回收試劑盒(DP209),天根生化科技(北京)有限公司;QuantiNova SYBR Green PCR試劑盒,德國QIAGEN公司。
Illumina Miseq PE300二代測序儀,美國Illumina公司;HWY-2112型恒溫培養搖床,上海智城分析儀器制造有限公司;LDZX-50KBS立式壓力滅菌鍋、SPX-250B型生化培養箱,上海博迅實業有限公司;SW-CJ-1D超凈工作臺,上海滬凈醫療器械有限公司;Nanodrop 2000/2000C 分光光度計,美國Thermo Fisher Scientific公司;PowerPac通用電泳儀電源、C1000 Touch PCR儀、CFX96 Touch實時定量PCR儀、ChemiDoc成像系統,美國Bio-Rad公司。
1.2 DNA提取
大曲微生物宏基因組的提取參考本研究室的前期研究[13-14]。
1.3 PCR擴增、Illumina Miseq測序和高通量數據分析
通過真菌內轉錄間隔區(ITS)1區研究真菌種群結構,利用ITS1F (5′-CTTGGTCATTTAGAGGAAGTAA-3′)和ITS2(5′-TGCGTTCTTCATCGATGC-3′)為引物對ITS1區序列進行擴增。PCR擴增程序:95 ℃ 5 min;95 ℃ 30 s,55 ℃ 30 s,72 ℃ 30 s,共25個循環;最后72 ℃ 10 min。擴增產物通過瓊脂糖凝膠回收、定量后,依據所需數據深度進行文庫制備,然后在Illumina Miseq PE300平臺上進行2×150 bp雙端測序。原始數據經Qiime (v1.8.0)處理后,再對序列進行拼接,并根據接頭序列將不同樣本進行分類。隨后依次去除序列上的接頭序列、標簽和引物序列,并去除低質量序列和嵌合體序列,得到優質數據(clean data)。再利用UCLUST將全部優質數據按照97%序列相似度聚類,進行操作分類單元(operational taxonomic units,OTU)劃分,提取代表序列,并得到OTU表[15]。將OTU代表序列與NCBI nt數據庫和UNITE數據庫(v 6.0)進行比對[16],獲得OTU物種分類信息。
“我這橫刀也不同意!”胖捕快走到兩排桌子之間,擺出了橫刀十三式的起手式,鐵氣激袖,刀光生寒,令他由怕老婆女兒的班頭,頓時變成威嚴肅殺、令人可畏的名捕。
1.4 熒光定量PCR
真菌18S rRNA基因的相對豐度定量參考ROUSK等[17]的研究。引物Fung(5′-ATTCCCCGTTACCCGTTG-3′)和NS1(5′-GTAGTCATATGCTTGTCTC-3′)用于擴增真菌18S rRNA基因部分序列[18]。通過擴增一系列濃度梯度的含有釀酒酵母18S rRNA基因全長拷貝的質粒建立標準曲線。PCR反應體系參考QuantiNova SYBR Green PCR試劑盒說明書。使用CFX96 Touch實時定量PCR儀進行所有qPCR反應,每個樣本重復3次。PCR擴增程序: 94 ℃、4 min,94 ℃、30 s,55 ℃、30 s,72 ℃、30 s,共40個循環;最后72 ℃、2 min。
1.5 絲狀真菌的純培養
取10 g曲粉加入到盛有90 mL無菌生理鹽水的250 mL錐形瓶中,并在搖床中以200 r/min振蕩1 h。然后將大曲生理鹽水混合物稀釋至終濃度為1 g/L,取200 μL均勻涂布于5種不同的真菌培養基上,并放置于28、37 和50 ℃培養。每種大曲在不同培養基和溫度條件下重復3次。
1.6 絲狀真菌鑒定與系統發育分析
使用引物ITS1(5′-TCCGTAGGTGAACCTGCGG-3′)和ITS4(5′-TCCTCCGCTTATTGATATGC-3′)對純培養獲得的絲狀真菌的ITS全長序列進行擴增[19]。50 μL的PCR反應體系包括:1×Taq PCR Mastermix、0.5 μmol/L引物、50 ng基因組DNA。PCR擴增程序:94 ℃、5 min;94 ℃、30 s,55 ℃、30 s,72 ℃、1 min,共35個循環;最后72 ℃、10 min。PCR產物使用通用DNA純化試劑盒純化后進行測序分析。利用NCBI BLAST工具(http://www.ncbi.nlm.nih.gov/blast)將獲得核酸序列進行手動比對,并使用MEGA (version 6.0)軟件對獲得的序列進行系統發育分析[20]。最后將獲得的系統發育樹通過iTOL進行可視化[21]。
1.7 次級代謝產物合成酶基因擴增
通過對次級代謝產物合成酶表達基因進行擴增,研究大曲絲狀真菌次級代謝產物合成能力。使用簡并引物(表2)擴增的次級代謝物合成基因包括非核糖體多肽類合成酶(non-ribosomal peptides synthase,NRPSs)、I型聚酮類合成酶(type I polyketide synthase,PKS I)、III型聚酮類合成酶(type III polyketide synthase,PKS III)、PKS-NRPS雜合類合成酶和萜烯類合成酶(terpene synthase,TPS)5種。25 μL的PCR反應體系包括:1×Taq PCR Mastermix、0.5 μmol/L引物、100 ng基因組DNA。每種基因的擴增包含1組陽性對照和2組陰性對照。陽性對照使用包含上述所有5種基因的A.parasiticus(CGMCC 3.6155)基因組;陰性對照除了使用無菌水作為擴增模板,還使用Saccharomycescerevisiae基因組作為擴增模板,因為該物種基因組不含任何上述5種次級代謝物合成基因[22]。PCR產物使用1%瓊脂糖凝膠電泳檢測,并使用Bio-Rad凝膠成像系統進行拍照。

表2 次級代謝產物合成基因及其簡并引物
注:簡并堿基符號:B=C/G/T;D=A/G/T;i=肌酐(inosine);K=G/T;M=A/C;R=A/G;S=C/G;Y=C/T。
2 結果與分析
2.1 高通量測序比較不同大曲中絲狀真菌的群落結構
通過利用高通量測序技術,在ZW、AGW和GW 3種大曲樣品中分別檢測到了217、235和207個真菌OTU。其中,絲狀真菌在3種大曲樣本中所占的比例穩定在25.6%~26.4%(表3),說明絲狀真菌的物種比例在不同類型的大曲中非常穩定,基本不受大曲類型的影響。
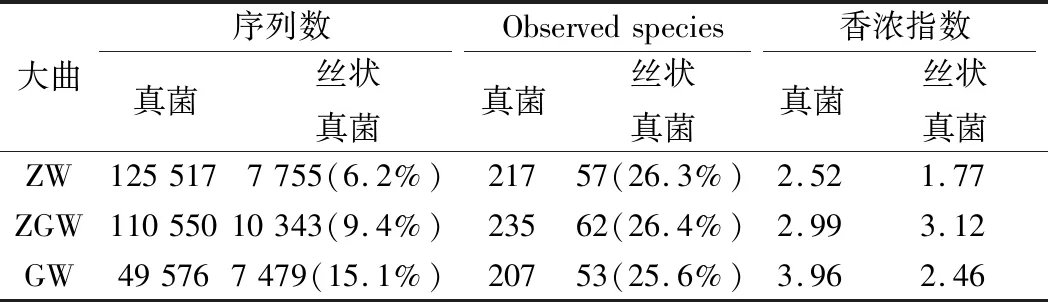
表3 大曲真菌及絲狀真菌多樣性分析
進一步將豐度高于1%的OTU序列使用NCBI BLAST進行手動比對,獲得OTU在種水平的物種分類信息。如圖1所示,不同樣本之間絲狀真菌的組成差異非常明顯。Aspergillus屬是絲狀真菌中主要的微生物屬,該屬在ZW、ZGW和GW樣本中比例分別為85%、58%和31%。Aspergillus屬是環境中廣泛存在的1種絲狀真菌,文獻報道該屬能高效產生葡糖淀粉酶(2 463±343 U/g)和α-淀粉酶(1 491±324U/g)[23]。
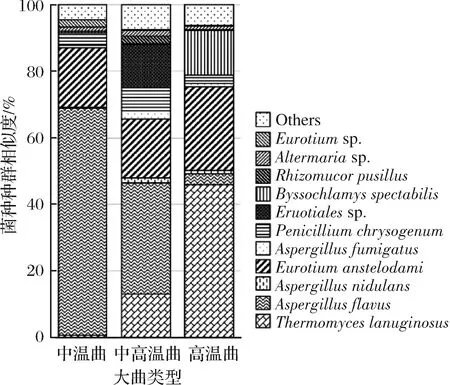
圖1 大曲中絲狀真菌種群結構(種水平)
Fig.1 Taxonomic classification of filamentous fungal reads at species level based on ITS1 region
2.2 大曲中絲狀真菌定量分析
大曲中絲狀真菌的數量通過實時熒光定量PCR(RT-PCR)技術結合高通量測序數據確定。首先利用一系列濃度梯度的含有釀酒酵母18S rRNA基因全長拷貝的質粒建立RT-PCR標準曲線。如圖2-A所示,良好的擴增效率(97%,通過E=10-1/斜率-1計算而來)和線性關系(R2=0.994 9)表明了標準曲線的準確性。然后根據絲狀真菌的序列數占真菌總序列數的比例,計算出絲狀真菌在不同大曲中的數量。從圖2-B可以看出,ZW樣本有最多數量的真菌(1.73×107個)和絲狀真菌(1.07×106個),GW樣本真菌(1.38×106個)和絲狀真菌(2.08×105個)最少,說明大曲中真菌和絲狀真菌的數量隨大曲發酵最高溫度的升高而減少,原因可能是因為較少的微生物可以在高溫環境下存活。

a-RT-PCR定量曲線;b-真菌與絲狀真菌18S rRNA基因拷貝數
圖2 RT-PCR結合高通量測序數據確定大曲中絲狀 真菌數量
Fig.2 Quantification of the filamentous fungi by using real- time PCR coupled with pyrosequencing
2.3 基于可培養的方法研究大曲中絲狀真菌的物種多樣性
本研究選擇GW樣本來研究絲狀真菌的物種多樣性,一方面,由于GW樣本中絲狀真菌序列數占真菌序列總數的比例最大,因而篩選到絲狀真菌的可能性更高;另一方面,來自極端環境的微生物產生天然產物的可能性更高。因為這些微生物可能是系統發育上全新的微生物,并且為了適應獨特的環境,還可能存在全新的天然產物合成途徑[25]。
由于不同類型的絲狀真菌對營養物質的需求可能不同,因此為了獲得盡可能多的絲狀真菌物種,使用5種不同類型的培養基進行篩選。在篩選過程中,對于具有相似形態和生長特征的絲狀真菌,僅保留1株。最終共篩選獲得50株絲狀真菌(表4),其中有45株是在28 ℃培養條件下獲得(PDA培養基24株,MEA培養基7株,CMA培養基3株,SDA培養基5株,CDA培養基6株),在37 ℃培養條件下并未獲得絲狀真菌單菌落,剩下5株在50 ℃培養條件下獲得(PDA培養基3株,MEA培養基2株)。
進一步對絲狀真菌的rDNA-ITS序列進行BLAST比對分析發現,50株分離株大部分與已報道的菌株具有99%以上的相似度,除了PDA-7和PDA-16分別只有98%和97%的相似度,說明PDA-7和PDA-16可能是分類上新的微生物種。這50株菌株由散囊菌目(Eurotiales)、毛霉目(Mucorales)、炭角菌目(Xylariales)和格孢菌目(Pleosporales)4個微生物目組成,共分為曲霉屬(Aspergillus)、紅曲霉屬(Monascus)、絲衣霉屬(Byssochlamys)、橫梗霉屬(Lichtheimia)、根毛霉屬(Rhizomucor)、毛霉屬(Mucor)、節菱孢屬(Arthrinium)、鏈格孢屬(Alternaria)、嗜熱真菌屬(Thermomyces)和Rasamsonia十個屬,其中Aspergillus屬絲狀真菌的種類最為豐富,共30株分離株屬于該微生物屬,隸屬于19個不同的種。
過去的研究曾采用單一培養基對高溫大曲中的絲狀真菌進行過篩選,但是僅能獲得有限種類的絲狀真菌(P.variotii、A.oryzae、P.namyslowskii、R.microsporus、Microascuscirrosus、Monascuspurpureus、P.chrysogenum和A.terreus[23])。而本研究中,同樣在高溫大曲中,篩選鑒定出至少28個不同的絲狀真菌種,說明大曲中絲狀真菌的物種豐富度可能被嚴重低估。
另外還發現,雖然大部分絲狀真菌分離株能在高通量測序結果中找到對應的微生物屬,但仍然有少數絲狀真菌僅能通過純培養獲得,例如Mucor和Arthrinium屬的微生物,其原因可能是用于高通量測序的通用引物的選擇性和覆蓋范圍限制了對所有物種的完整表征[26]。

表4 基于純培養獲得的絲狀真菌的物種多樣性和分類群信息
基于以上結果可以說明,大曲中絲狀真菌的物種豐富度可能被顯著低估,同時基于純培養的方法在目前微生物群落研究中仍然發揮著不可替代的作用。
2.4 大曲絲狀真菌來源的次級代謝產物合成基因多樣性分析
為了進一步探究大曲來源的絲狀真菌合成天然生物活性物質的遺傳潛力,利用簡并引物對篩選得到的50株絲狀真菌的NRPSs、TPS、PKS-NRPS、I型和III型PKSs 5種次級代謝產物合成基因進行了擴增。從圖3可以看出,50株分離株全部含有至少1個次級代謝產物合成基因。其中48%的絲狀真菌(24株)含有所有靶基因;32%(16株)含有4個靶基因;16%(8株)含有3個靶基因;2%(1株)含有2個靶基因;2%(1株)僅含有1個靶基因。結果表明,大曲來源的絲狀真菌具有合成多種次級代謝產物的遺傳潛力。
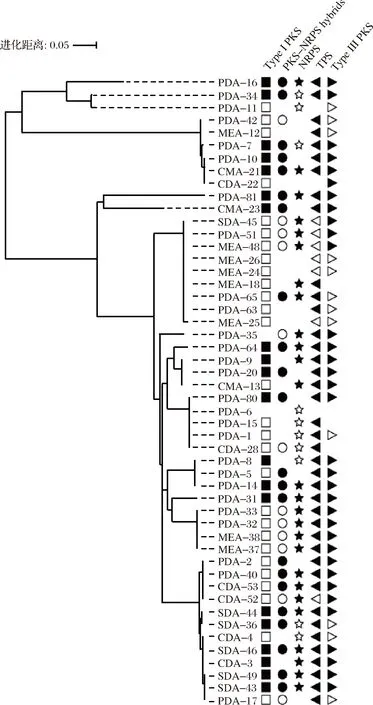
圖3 GW來源的絲狀真菌的系統發育樹及其次級代謝產物合成基因分布
Fig.3 Phylogenetic tree of filamentous fungi derived from GW coupled with presence of genes involved in synthesis of bioactive compounds
注:不同的形狀代表不同的次級代謝物合成基因;填充的形狀表示含有2個以上的SM合成基因,無填充的形狀表示僅含有1個SM合成基因,空缺代表沒有相應的SM合成基因。
另外,與前人研究發現真菌含有多種PKS基因的結果相一致[27],本研究幾乎在所有分離株中都觀察到至少1個I型和III型PKSs基因。文獻報道,PKS基因參與多種天然產物的合成,例如,I型PKS基因能夠合成大量具有藥理學活性的次級代謝產物,包括紅霉素、免疫抑制劑FK506以及抗寄生蟲的阿維菌素類化合物[28],因此近年來PKS基因受到了廣泛的關注[25, 27]。
相同微生物種的絲狀真菌也有可能含有不同的次級代謝產物合成基因,如Lichtheimiaramosa(PDA-10、CMA-21和CDA-22)和A.sydowii(CDA-3、CDA-4、PDA-17、SDA-43和MEA-49)。YU等[12]從海綿中篩到的8株Aspergillusterreus,也顯示出能夠產生不同的次級代謝產物的現象。造成這一現象的原因可能是真菌的次級代謝基因簇通過多次重排、重復或丟失而迅速進化[29]。大曲中的絲狀真菌在適應環境的過程中,可能丟失或者獲得了一些次級代謝基因。
為驗證真菌天然產物的合成,仍需通過模擬大曲發酵環境對分離株進行培養,并對產物進行化學鑒定和細胞毒性測試,以證實大曲來源的絲狀真菌是否能夠合成天然生物活性物質。
3 結論
通過高通量測序手段對3種不同類型的成品曲中的絲狀真菌的群落結構進行了分析,觀察到Thermomyces和Aspergillus屬的豐度在不同類型的大曲中差異非常明顯,能用于區分不同類型的大曲;另外利用純培養的手段對高溫大曲中絲狀真菌進行了篩選,共獲得了來自10個不同屬的50株絲狀真菌,表明大曲中絲狀真菌的物種豐富度被低估;進一步對這50株絲狀真菌基因組中參與次級代謝產物合成的基因進行了擴增,結果發現高溫大曲來源的絲狀真菌具有強大的次級代謝產物合成潛力。但是由于在大多數情況下,次級代謝產物合成基因是沉默的,因此仍需有針對性的檢測大曲以及白酒中的生物活性成分,并分析這些物質的微生物來源。綜合以上結論可以看出,大曲是一種優良的微生物庫和基因庫,包含豐富的微生物資源和功能基因資源。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
