根癌農桿菌介導山葡萄VaCBF3基因轉化歐美楊111的研究
2019-08-30 07:07:52馮連榮矯麗曼王占斌趙繼梅宋立志梁德軍
西南林業大學學報 2019年5期
馮連榮 矯麗曼 王占斌 趙繼梅 尹 杰 宋立志 梁德軍
( 1. 遼寧省楊樹研究所,遼寧 營口 115213;2. 東北林業大學林學院,黑龍江 哈爾濱 150040)
楊樹是遼寧省平原地區的主要造林樹種[1],近年來受異常氣候(極端干旱、寒潮等)影響,遼寧楊樹接連發生大面積災害,其中2006年、2009年、2013年楊樹災害發生規模較大,損失嚴重,一些含有南方型美洲黑楊基因的楊樹品種受災較重,其中歐美楊107(Populus×euramericana cv.‘ Neva’)、 歐 美 楊 108( P.×euramericana cv.‘Guariento’)幾乎全軍覆沒[2]。除氣候因素外,樹種抗逆性差是楊樹發生災害的主要內在因素,所以培育耐低溫楊樹品種、提高楊樹抗寒性是楊樹生產迫切需求。目前楊樹抗寒育種主要依靠人工雜交和天然雜種選育,不僅周期長而且對提高植物抗寒性的作用不大。
近年來,大量的抗逆基因被分離,作為林木基因工程研究的模式植物[3],楊樹轉抗逆基因研究取得了重大進展,但關于抗寒方面的研究仍處于探索階段,開展了一些導入單一的功能基因如擬南芥(Arabidopsis thaliana)Fe-SOD基因[4]、胡蘿卜(Daucus carota)抗凍蛋白基因[5]、毛白楊(P. tomentosa)油酸去飽和酶基因FAD基因[6]和轉錄因子基因如擬南芥CBF1(CRT/DRE binding factor) 基 因[7]、 銀 腺 楊 MBF1(multiprotein bridging factor)基因[8]的研究,這些研究均獲得了陽性轉基因植株,并在一定程度上提高了轉基因植株的抗寒性。CBF(CRT/DRE binding factor)轉錄因子是一類受低溫誘導的反式作用因子[9],屬于AP2/ERF家族,可以與COR(cold-regulated)基因啟動子中的CRT/DRE(C-repeat/dehydration responsive element)等順式作用元件特異性結合[10],激活低溫和脫水響應基因的表達,促進脯氨酸、可溶性糖類等物質的積累[11],從而提高植物的抗寒性。目前已經在陸地棉(Gossypium hirsutum)[12]、哈密大棗(Zizyphus jujuba)[13]、番茄(Lycopersicon esculentum)[14]、黃瓜(Chcumis satires)[15]、 茄 子 ( Solanum melongena)[16]、 苜蓿(Medicago sativa)[17]等植物中得到了應用。CBF轉錄因子基因在楊樹轉化上的報道還比較少,目前僅可見毛果楊CBF/DREB1基因家族生物信息分析[18]、胡楊CBF基因家族的鑒定及表達特性分析[19]及大青楊PuCBF轉化煙草[20]等方面的少數報道。筆者利用RT-PCR方法克隆得到山葡萄(Vitis amurensis)CBF3(VaCBF3)基因[21]并分別構建了由組成型啟動子CaMV 35S啟動子和擬南芥rd29A基因啟動子驅動的植物表達載體[22],確定了轉化過程中抗生素使用濃度[23]。本研究通過農桿菌介導的葉盤法建立了歐美楊111(P. euramericana ‘Bellotto’)高效遺傳轉化體系,旨在為歐美楊111號抗寒育種及應用提供參考及擴繁材料。
1 材料與方法
1.1 材料來源
1.1.1 受體材料
歐美楊111 當年生枝條采自遼寧省楊樹研究所基因庫,經室內消毒、外植體接種獲得無菌組培苗。組培苗培養于(25±1)℃條件下培養,光周期12 h/d,光照強度2 000~2 500 lx。
1.1.2 菌株和質粒
工程菌為根癌農桿菌(Agrobacterium tumiefaciens)LBA4404,攜帶有擬南芥rd29A基因啟動子調控的VaCBF3基因植物表達載體質粒。質粒大小約為11.9 kb,其上攜帶卡那霉素抗性標記基因,見圖1。
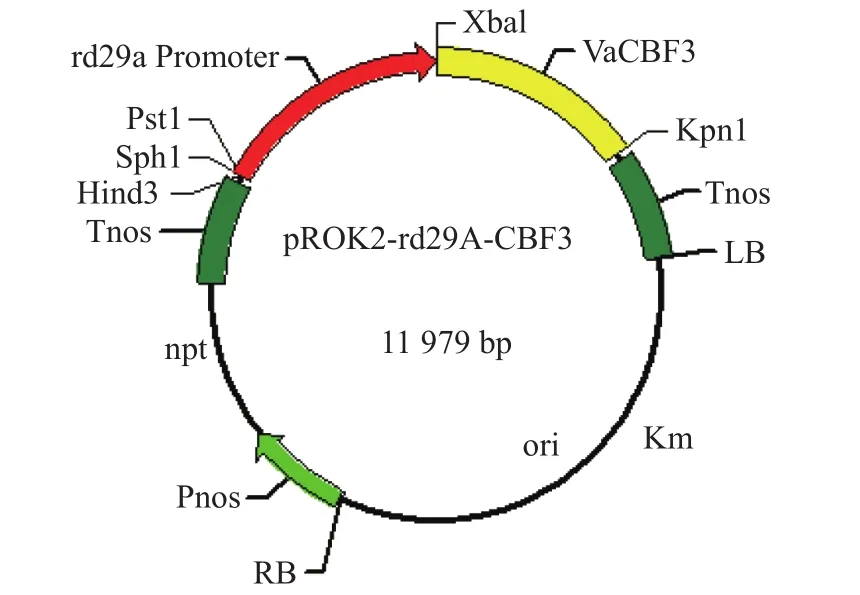
圖 1 質粒圖譜Fig. 1 The plasmid profile
1.1.3 培養基
遺傳轉化用培養基:
Ⅰ歐美楊111葉片分化培養基:MS+6-BA 0.5 mg/L+NAA 0.5 mg/L;
Ⅱ歐美楊111莖段生根培養基:1/2MS+IBA 0.4 mg/L;
Ⅲ脫菌分化培養基:MS+6-BA 0.5 mg/L+NAA 0.5 mg/L+Kan 10 mg/L;
Ⅳ選擇生根培養基:1/2MS+IBA0.4 mg/L+Kan 20 mg/L+Cef 50 mg/L;
每升培養基中加蔗糖20 g,瓊脂6 g,用1 mol/L NaOH調節pH至6.0~6.5,121 ℃ 0.1 MPa滅菌20 min備用。
LB液體培養基:胰蛋白胨10 g/L,酵母提取物5 g/L,NaCl 10 g/L,用5 mol/L NaOH調pH至7.0,121 ℃ 0.1 MPa滅菌20 min備用。
1.1.4 酶及生化試劑
Taq DNA Polymerase購自Thermo公司;TIANGEN DNAsecure新型植物基因組DNA提取試劑盒購自北京天根生化科技有限公司;卡那霉素購自Sigma公司;其他生化試劑均為國產分析純。
引物序列為:
RD1 為:5′-GCAAGCTTA ACGCATGATTTGATGGAGGA-3′;
RD2 為:5′-GCTCTAGACTTTCCAATAGAAGTAATC-3′
C1 為:5′-GCGGATCTAGAATGGAATCGGAGCGT-3′;
C2 為:5′-GGCGGTACCTCAAGCTATGCATCCAAC-3′。
RD1/RD2擴增擬南芥rd29A基因啟動子,片段長度945 bp;C1/C2擴增山葡萄CBF3(VaCBF3)基因,片段長度854 bp;RD1/C2擴增rd29A基因啟動子+VaCBF3基因,片段長度1 799 bp。
1.2 實驗方法
1.2.1 農桿菌介導目的基因遺傳轉化及抗性植株的獲得
以歐美楊111無菌組培苗葉片為外植體,采用農桿菌介導法[24]轉化山葡萄VaCBF3基因。選擇完全展開、深綠色的葉片,用手術剪刀剪2~3下,剪斷主脈,葉面朝上接種至培養基Ⅰ上進行預培養。于超凈工作臺上,將工程菌菌液(OD600≈0.5)轉入至50 mL離心管中,3 000 r/min,4 ℃離心5 min,去除上清液,將收集到的菌液用LB液體培養基稀釋至終濃度為OD600≈0.5,然后將預培養的葉片放入到稀釋好的菌液中,侵染2~10 min,侵染期間不停的搖動,使菌液與葉片充分接觸。侵染過的葉片取出后置于無菌干燥的濾紙上,吸附葉片其表面的殘余菌液,然后接種到培養基Ⅰ上,25 ℃黑暗條件下共培養,當葉片邊緣隱約可見菌斑時,共培養結束。將經過共培養的外植體轉移到加有選擇壓的脫菌分化培養基Ⅲ上,25 ℃條件下進行選擇培養。如果獲得抗性不定芽,將抗性芽剪下繼代擴繁培養。選擇培養3~4周后,將抗性芽轉入相應的附加選擇壓的分化培養基中令其生長或誘導分化。將獲得的長至2~3 cm的抗性芽切下,接種到選擇生根培養基Ⅳ中,進行生根篩選,培養條件同上,2周后可長出不定根。
1.2.2 不同因素對歐美楊111轉化的影響
采用單因素實驗設計,研究預培養時間、菌液濃度、侵染時間、共培養時間對轉化的影響。轉化基本條件為:外植體預培養時間為3 d,菌液濃度為OD600=0.2,侵染時間為4 min,共培養時間為3 d。以轉化基本條件為基礎,預培養時間設置為:1、2、3、4、5 d;菌液濃度為:0.1、0.2、0.3、0.4、0.5;侵染時間:2、4、6、8、10 min;共培養時間:1、2、3、4、5 d。每個處理侵染葉片20~31個,3次重復,使用Excel、DPS 15.0軟件對數據進行整理分析。
1.2.3 抗性植株分子檢測
采用天根快捷型植物DNA提取試劑盒提取抗性植株和對照植株葉片總DNA,操作步驟按說明書進行,DNA于-20 ℃下儲存備用。
以提取的總DNA為模板,未轉化對照植株DNA為陰性對照,質粒DNA為陽性對照,RD1/RD2和C1/C2及RD1/C2為引物進行PCR擴增。反應體系(25 μL):10×PCR Buffer 2.5 μL、dNTP(2.5 mmol/L) 2.0 μL、上游引物(10 μmol/L)0.5 μL、下游引物(10 μmol/L)0.5 μL、模板 1.0 μL、Taq 酶 0.25 μL、ddH2O 加 至 25 μL。 反 應 條 件為:94 ℃ 預變性 5 min,94 ℃ 變性 30 s,53 ℃退火30 s,72 ℃延伸45 s,72 ℃延伸10 min,反應30個循環,4 ℃終止反應。反應結束后,取6 μL反應產物經0.8%瓊脂糖凝膠電泳進行檢測。
2 結果與分析
2.1 不同因素對歐美楊111轉化的影響
2.1.1 預培養時間
預培養時間對歐美楊111葉片分化和轉化的影響,結果見圖2a:隨著預培養時間的增加,抗性芽率逐漸提高,預培養1 d時抗性芽率為0,可能是由于葉片的傷口還未來得及修復,農桿菌的毒害作用使葉片從傷口處迅速褐化死亡。當預培養時間為3 d時,抗性芽率達到最高12.34%,隨后呈現下降趨勢。原因可能是隨著預培養時間的延長,葉片邊緣細胞已經出現芽的分化,葉片切口快要愈合,創傷產生的酚類等vir基因誘導物減少,不利于農桿菌從傷口侵入,降低轉化率。因此,在進行歐美楊111遺傳轉化時,預培養時間以3 d為宜。
2.1.2 菌液濃度
農桿菌的濃度和侵染時間直接影響T-DNA向植物細胞的轉移幾率[23]。農桿菌一般菌液濃度范圍是OD600=0.1~0.5。本研究將菌液濃度分別稀釋至0.1、0.2、0.3、0.4和0.5,結果見圖2b:隨著菌液濃度的增加,歐美楊111葉片產生的抗性芽率隨之提高。當菌液濃度為0.3~0.4時,抗性率達到較大值9.68%~11.36%,隨后又轉為下降趨勢,菌液濃度過高,菌體本身易相互凝結,不利于在外植體附著,從而降低轉化率,而濃度過低時不利于基因的轉入。因此,適宜于歐美楊111轉化的農桿菌菌液濃度為0.3~0.4。
2.1.3 侵染時間
侵染時間的長短對影響轉化效率具有一定的影響。本研究將侵染時間設置為2、4、6、8 min和10 min,結果表明:侵染時間在2~4 min之間時,歐美楊111葉片抗性芽得率較高,分別達到12.12%和19.05%,之后隨著侵染時間的延長,抗性芽率逐漸降低,見圖2c,可能是由于侵染時間過長,農桿菌過度繁殖、毒性過強而使外植體缺氧軟腐。因此,適宜于歐美楊111轉化的侵染時間為2~4 min。
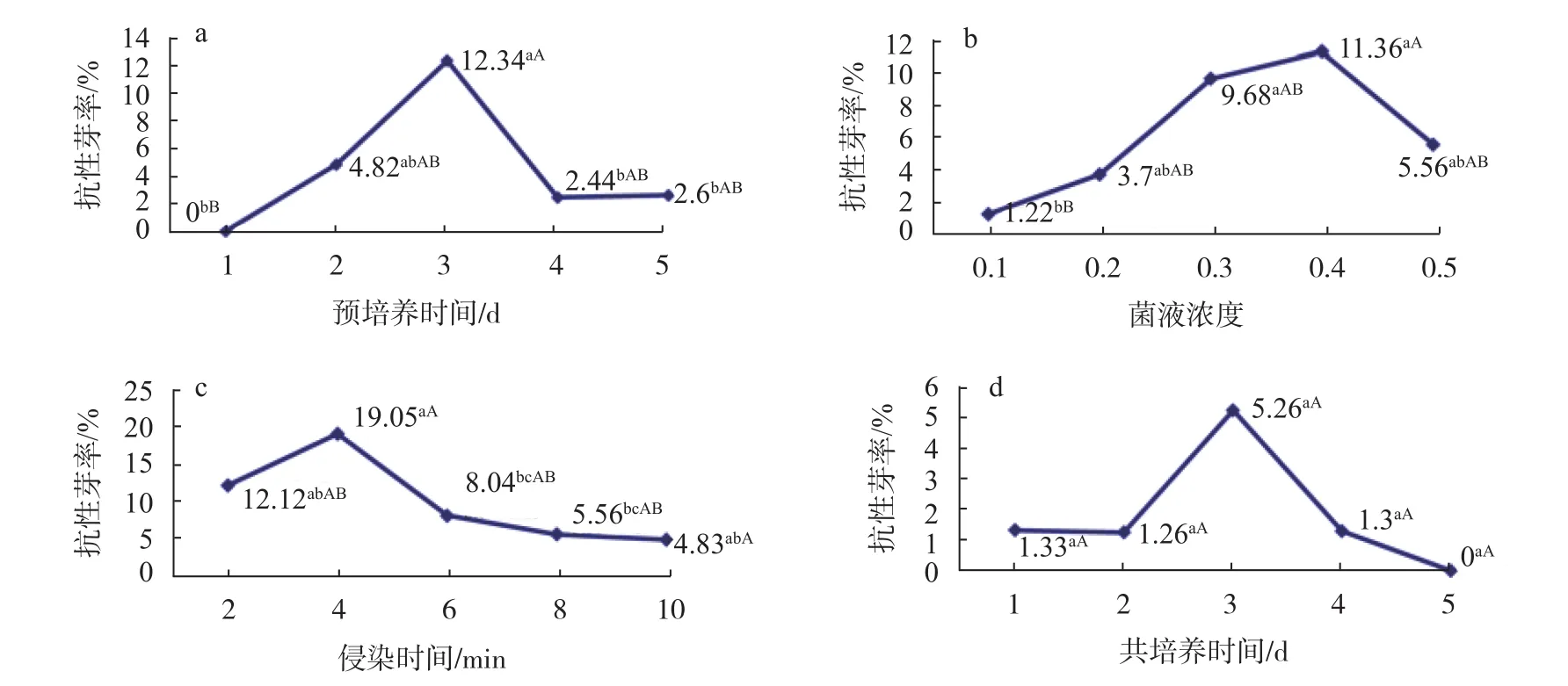
圖 2 不同因素與歐美楊111葉片抗性芽率的關系Fig. 2 The relationship between different factors and the rate of leaves resistant bud of P. euramericana 'Bellotto'
2.1.4 共培養時間
農桿菌與受傷部位共存16 h后才能誘發受體材料產生腫瘤,完成T-DNA完成轉移和整合[24]。因此,對歐美楊111的遺傳轉化共培養時間必須長于16 h。本研究將預培養時間設置為1、2、3、4、5 d,分析共培養時間對歐美楊111轉化的影響,結果見圖2d:隨著共培養時間的延長,抗性芽率增加,當共培養時間為3 d時,抗性芽率達到最大值5.26%,隨后抗性芽率降低,共培養時間為5 d時,抗性芽率更是降低為0。共培養時間過長時,長時間黑暗培養導致產生白化芽,白化芽轉入到選擇性培養基后,葉片和芽很快死亡,無法獲得抗性芽。因此,對于歐美楊111葉片的遺傳轉化,共培養時間在3 d時比較合適。
2.2 抗性植株的分子檢測
2.2.1 抗性植株總DNA提取
提取 66株抗性植株的DNA,DNA取5 μL DNA溶液混合1 μL上樣緩沖液加入到凝膠點樣孔,在約5 V/cm電場中電泳,使用凝膠成像系統觀察、拍照,1~21號抗性植株DNA電泳結果見圖3:電泳結果顯示提取的歐美楊111總DNA質量較好,條帶清晰,雜質較少,雖有DNA降解,但也可以作為PCR反應的模板。
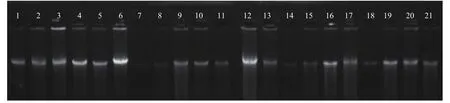
圖 3 歐美楊111 DNA電泳Fig. 3 The DNA electrophoretogram of P. euramericana 'Bellotto'
2.2.2 抗性植株的PCR檢測
歐美楊111抗性植株葉片總DNA經RD1/RD2、C1/C2和RD1/C2 3對引物進行PCR測,其中有4株陽性植株(圖4)分別擴增出目的條帶,片段大小分別約為:950、850 bp和1 800 bp。其中的1號、2號植株PCR產物檢測見圖5,并對1號,2號PCR擴增產物進行了測序,測序結果經過BLAST比對,與目的基因片段進行序列比對(表1),片段一致性達到了99%~100%,可以初步判斷目的基因片段導入到受體材料中。

圖 4 轉VaCBF3基因歐美楊111抗性芽及陽性植株Fig. 4 The resistant buds and positive plants of P. euramericana 'Bellotto' with transforming VaCBF3 gene
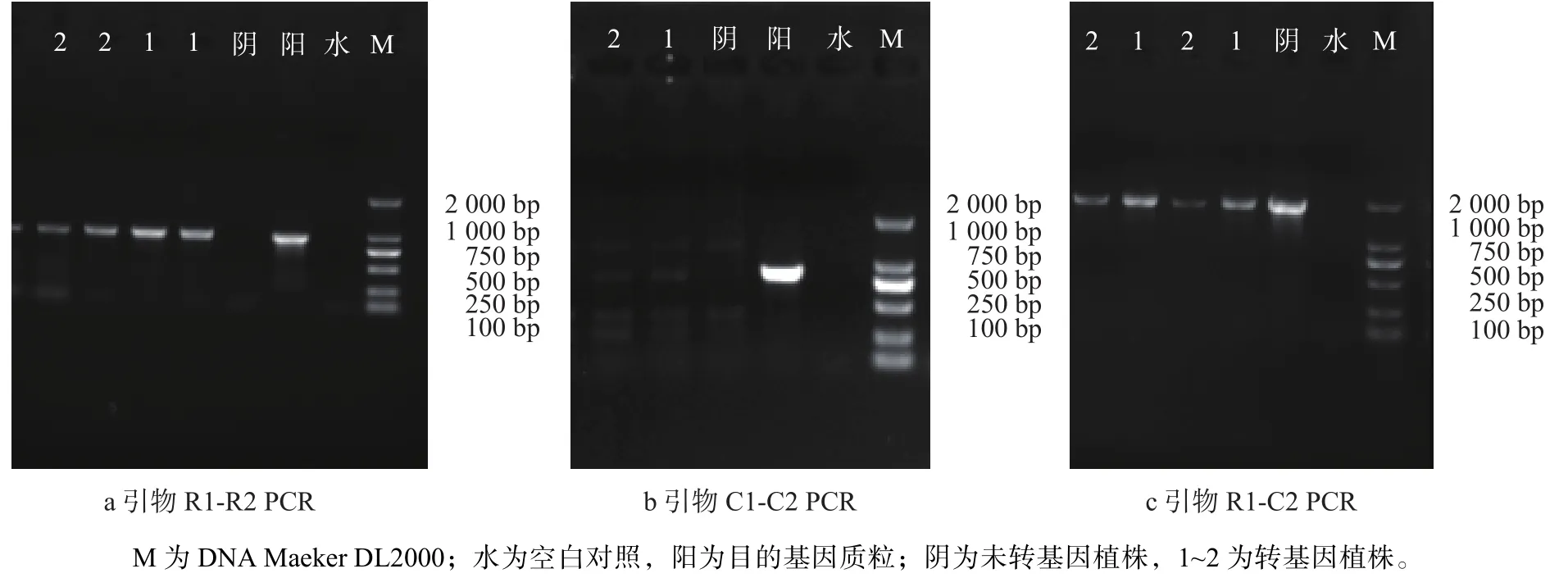
圖 5 PCR產物檢測Fig. 5 Detection of PCR products
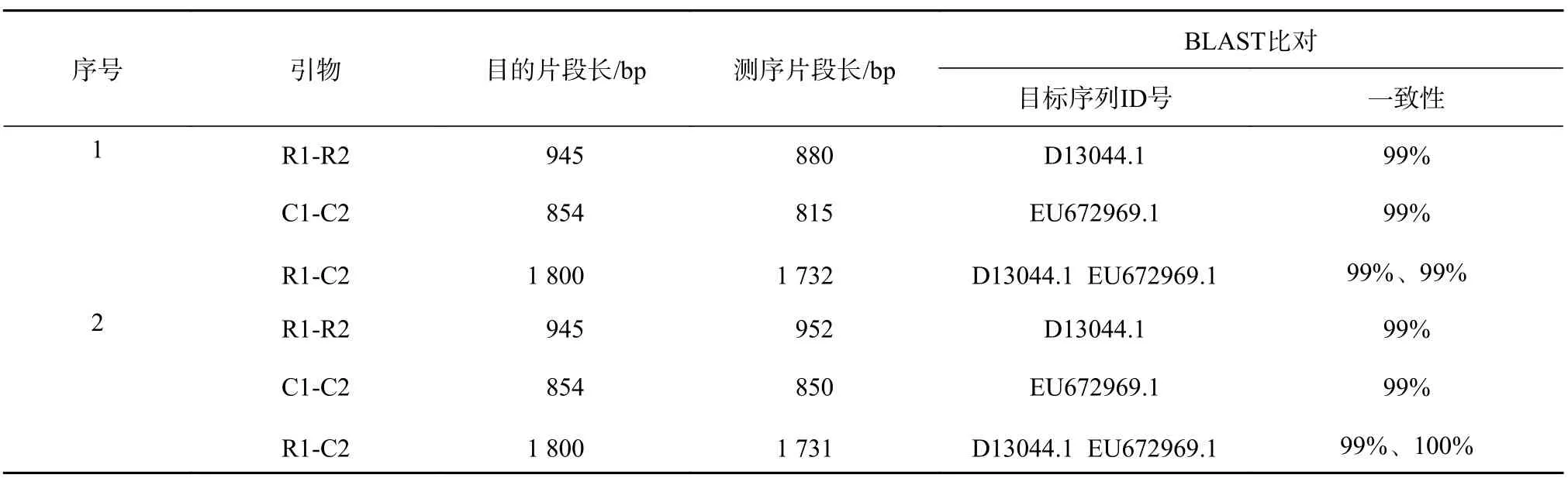
表 1 轉基因1、2號植株PCR產物測序結果Table 1 The sequencing results of PCR products in transgenic plants 1 and 2
3 結論與討論
根癌農桿菌介導的遺傳轉化系統,是當今楊樹遺傳轉化的主要方法[25]。轉化成功與否受楊樹基因型、外植體類型、培養條件、抗生素等因素影響。建立高效的遺傳轉化體系,是提高轉化效率的重要環節,除了確定受體材料對抗生素的敏感性,還需要篩選轉化過程中適宜的預培養時間、菌液濃度、侵染時間和共培養時間等。甄成[26]研究毛果楊(P. trichocarpa)莖段最佳侵染條件為菌液濃度OD600=0.4,侵染時間為20 min,共培養時間為2 d,在侵染及共培養過程中添加80 μmol/L的乙酰丁香酮,轉化率為26.7%。朱偉康[27]確定轉Bt基因歐洲黑楊(P. nigra)遺傳轉化最佳條件為:菌液濃度OD600=0.3,侵染時間10 min,共培養時間2 d,轉化率為18.0%。趙芳方[28]確定毛白楊(P. tomentosa)最佳遺傳轉化體系為菌液濃度OD600=1.0,侵染時間為9 min,共培養時間為25 h。孫偉博等[29]采用正交實驗設計研究‘南林895楊’(P. delotides×P. euramericana cv. ‘Nanlin895’)合適的遺傳轉化體系為:預培養3 d,菌液濃度OD600=0.8,侵染8 min,共培養3 d。本研究采用單因素實驗篩選出歐美楊111遺傳轉化過程中適宜的預培養時間為3 d,適宜菌液濃度為OD600≈0.3~0.4之間,適宜侵染時間為2~4 min,適宜共培養時間為3 d,轉化效率為5.26%~19.05%。但本研究只是采用單因素梯度實驗方法來探索預培養時間等影響轉化效率的4個關鍵因素的最佳水平,不能確定各因素間的交互作用,還有待于進一步的優化。另外提高轉化效率的另一種有效手段是在農桿菌培養、農桿菌和外植體共培養的培養基中添加vir基因誘導物,如乙酰丁香酮、羥基乙酰丁香酮等[21],其中以乙酰丁香酮效果最佳,使用較為廣泛,使用濃度一般為20~200 μmol/L[20],有待于在下一步優化過程中進行嘗試。
本研究利用農桿菌介導法將山葡萄VaCBF3基因導入到歐美楊111中,獲得4株抗性植株,初步證明VaCBF3基因已經整合到歐美楊111基因組中。下一步需要進一步在RNA水平、蛋白水平進行分子檢測,并進一步驗證VaCBF3的表達水平、表達穩定性,同時通過測試電導率、丙二醛、可溶性糖等抗寒生理指標,在生理上明確VaCBF3基因對提高楊樹抗寒性的作用。CBF轉錄因子是一個冷適應過程中的關鍵性調控因子,CBF的發現為基因工程改良植物耐逆性提供了一種全新的技術途徑。在南方植物北移,換季節種植等基因改良品種方面具有重要應用價值。
[ 參 考 文 獻 ]
[1]梁德軍. 遼寧省楊樹主要造林品種[M]. 沈陽: 遼寧大學出版社, 2017.
[2]劉巍, 蔄勝軍, 紀純陽, 等. 遼寧省楊樹災害發生的原因及對策 [J]. 防護林科技, 2014(6): 70-73.
[3]丁莉萍, 王宏芝, 魏建華. 楊樹轉基因研究進展及展望 [J]. 林業科學研究, 2016, 29(1): 124-132.
[4]Arisi A C M, Cornic G, Jouanin L, et al. Overexpression of iron superoxide dismutase in transformed poplar modifies the regulation of photosynthesis at low CO2partial pressures or following exposure to the prooxidant herbicide methyl viologen [J]. Plant Physiology, 1998, 117(2): 565-574.
[5]李春霞. 抗凍蛋白基因對山楊等植物遺傳轉化的研究[D]. 哈爾濱: 東北林業大學, 2003.
[6]Benedict C, Skinner J S, Meng R G, et al. The CBF1-dependent low temperature signalling pathway, regulon and increase in freeze tolerance are conserved in Populus spp. [J]. Plant, Cell and Environment, 2006,29(7): 1259-1272.
[7]周洲. 轉脂肪酸去飽和酶基因PtFAD2和PtFAD3銀腺楊84K的抗寒性研究[D]. 北京: 中國林業科學研究院, 2007.
[8]許東. 楊樹抗寒轉錄因子PlMBF1b的克隆及功能分析[D]. 泰安: 山東農業大學, 2016.
[9]Stockinger E J, Gilmour S J, Thomashow M F. Arabidopsis thaliana CBF1 encodes an AP2 domain-containing transcriptional activator that binds to the C-repeat/DRE, a cis-acting DNA regulatory element that stimulates transcription in response to low temperature and water deficit [J]. Proceedings of the National Academy of Sciences, 1997, 94(3): 1035-1040.
[10]李健, 王雅晴, 劉洋, 等. CBF轉錄因子在植物抗逆和生長發育中的重要功能 [J]. 植物生理學報, 2017,53(12): 2045-2056.
[11]Gilmour S J, Sebolt A M, Salazar M P, et al. Overexpression of the Arabidopsis CBF3Transcriptional activator mimics multiple biochemical changes associated with cold acclimation [J]. Plant Physiology, 2000,124(4): 1854-1865.
[12]馬劉峰, 陳蕓, 任羽, 等. 棉花CBF2基因克隆和超表達CBF2棉花增強抗冷性 [J]. 植物生理學報, 2018,54(2): 255-264.
[13]高啟明, 王斌, 賽買提·吐爾遜, 等. 農桿菌介導的CBF基因轉化哈密大棗 [J]. 北方園藝, 2016(18):94-98.
[14]王沛文, 朱文哲, 劉陽, 等. 多毛番茄冷誘導轉錄因子CBF1轉化番茄的研究 [J]. 江蘇農業科學, 2015,43(4): 30-35.
[15]譚克, 趙福順, 吳慧杰, 等. 冷誘導基因轉錄因子CBF1轉入黃瓜的研究 [J]. 北方園藝, 2015(9): 79-82.
[16]孟平紅, 萬發香, 王永清, 等. 冷誘導轉錄因子CBF3轉化茄子的初步研究 [J]. 中國蔬菜, 2013(10): 36-43.
[17]徐春波, 王勇, 趙海霞, 等. 冷誘導轉錄因子AtCBF1轉化紫花苜蓿的研究 [J]. 草業學報, 2012, 21(4):168-174.
[18]丁咚, 陳亞娟, 崔進榮, 等. 毛果楊 CBF/DREB1基因家族生物信息學分析 [J]. 西南農業學報, 2018, 31(3):457-461.
[19]賈會霞, 李建波, 孫佩, 等. 胡楊CBF基因家族的鑒定及表達特性分析 [J]. 分子植物育種, 2017, 15(2):492-500.
[20]姜洋. 大青楊PuCBF遺傳轉化基礎及功能驗證[D].哈爾濱: 東北林業大學, 2017.
[21]Wang Z B, Feng L R, Wang J J, et al. Vitis amuerensis CBF3gene isolation, sequence analysis and expression [J]. Agricultural Sciences in China, 2010,9(8): 1127-1132.
[22]馮連榮, 王占斌, 宋立志. 2種啟動子驅動下的VaCBF3基因植物表達載體構建 [J]. 沈陽農業大學學報, 2011, 42(5): 559-564.
[23]馮連榮, 張興芬, 尹杰, 等. 楊樹農桿菌介導遺傳轉化中抗生素濃度的篩選 [J]. 西南林業大學學報, 2014,34(4): 31-35.
[24]王關林, 方宏筠. 植物基因工程 [M]. 2 版. 北京: 科學出版社, 2002
[25]馮連榮, 宋立志, 張妍, 等. 根癌農桿菌介導楊樹遺傳轉化的影響因素 [J]. 西北林學院學報, 2015, 30(3):120-126.
[26]甄成. 毛果楊組培再生及遺傳轉化體系研究[D]. 哈爾濱: 東北林業大學, 2016.
[27]朱偉康. 轉Bt基因歐洲黑楊組織培養再生體系優化及其TA29-Barnase基因遺傳轉化[D]. 楊凌: 西北農林科技大學, 2016
[28]趙芳方. 毛白楊再生體系的建立及Barnase基因轉化的初步研究[D]. 鄭州: 鄭州大學, 2014.
[29]孫偉博, 于娟, 潘惠新, 等. ‘南林895楊’遺傳轉化體系的優化 [J]. 林業科技開發, 2013, 27(6): 85-88.
