
Cu2+脅迫對狹葉香蒲種子萌發與生長及抗氧化物活性的影響
2019-09-05 13:33:19黃振燕賀柯翔鄢夢玲楊賢均
現代農業科技 2019年5期
黃振燕 賀柯翔 鄢夢玲 楊賢均
摘要以狹葉香蒲種子及幼苗為材料,研究了0、50、100、150、200、300μmol/LCu2+脅迫下狹葉香蒲種子的萌發特性,以及0、250、500、750、1000μmol/LCu2脅迫下狹葉香蒲幼苗的生理響應。結果表明,當Cu2+濃度增加時,狹葉香蒲種子的萌發率、萌發勢和活性逐漸減弱;隨著Cut濃度增加,葉片和根中的POD活性上升,且在750~1000μmol/LCu2+脅迫下葉片的POD活性顯著高于對照組;在Cu2*影響下幼苗的子葉以及根中AsA和MDA顯著高于對照組。當Cu2+濃度增加時,狹葉香蒲葉片和根部的MAD呈先上升后趨于穩定的趨勢,且與對照組無顯著差異。可見,狹葉香蒲幼苗葉片對Cu2+脅迫的敏感性比根部強,其種子可在輕度(0~300μmol/)污染的水體中萌發,狹葉香蒲幼苗對Cu2*存在著可耐性,可用于Cu2+輕度污染和中度污染的治理和修復。
關鍵詞 狹葉香蒲;Cut脅迫;種子萌發;抗氧化物活性
中圖分類號 S567.23+9
文獻標識碼 A
文章編號 1007-5739(2019)05-0157-03
隨著社會的進步與經濟的快速發展,水體受重金屬的影響日益嚴重。這種影響通過整條食物鏈的集聚,直接或間接地威脅人類的生存環境"。在處理水質被重金屬污染的修復技術中,采用水生植物進行修復的方法越來越受到業內同行的認可,其具有事半功倍的環境治理效益、不會對水體構成二次污染治理成本價格較低、無需中間運送費用等優勢12-3)。在眾多重金屬污染問題中,其中污染比較嚴重的是Cu2+,其一直是惡性循環體,亟待人們提出相應的治理方案。通過種植轉化Cu2+毒害能力比較好的水生植物,將Cu2+對生態系統的污染導人良性循環,是保護水體良性水質的有效方法之一。
狹葉香蒲(Typhaangustifolia?Linn.)是香蒲科香蒲屬植物,多年生水生或沼生草本,根部呈淡黃色、黃灰色,發達部分呈白色,生長在湖水、小溪、水池岸邊,濕地水渠經常可見,即使地面出現龜裂也能繼續生長,也可生長在濕地里。陳桂珠等、陽承勝等研究發現,香蒲屬植物對化學工廠產生的污染物(其中含有Cu、Zn.Cd等重金屬元素)可以進行治理,有改善水體污染的功能。張開明等阿劉素純等間Zhangs等嘟對重金屬污染對植物的影響進行了研究。關于Cu2+對綠色植物生長機理的影響研究有很多18-10但對Cu2+脅迫下狹葉香蒲種子的萌發以及幼苗生長的影響還鮮見報道。本研究以狹葉香蒲為材料,探討不同程度Cu2+脅迫下狹葉香蒲種子萌發及其幼苗生長機理,以期為狹葉香蒲凈化Cu2污染水體提供參考。
1材料與方法
1.1試驗材料
試驗材料為狹葉香蒲的蒲棒(種子)以及當年生幼苗,分別于2016年9月20日和2017年3月22日采集于湖南省邵陽市大祥區池塘。
1.2試驗準備
1.2.1實地采取種子。采用水泡選法、鼓風機選法在收集好的狹葉香蒲堆中選出肥大、營養器官健全的種子,并去掉種皮上的附著物。
1.2.2狹葉香蒲種子萌發的最適溫度。本次課題以研究不同Cu2+脅迫濃度對狹葉香蒲種子萌發特性的影響為主,且本研究在狹葉香蒲種子溫度試驗結果的基礎之上進行。因此,先進行溫度試驗,即設置10、15、20、25、30C共5個溫度梯度,取消毒后的培養皿盛放大小一致的飽滿狹葉香蒲種子5份,每份50顆,加入蒸餾水至種子1/3處,再放人培養箱中在上述溫度梯度條件下開始試驗。在試驗階段,每隔24h觀察培養皿的水分情況并記錄狹葉香蒲種子的萌發情況,在試驗開始后的第7天,首先觀察到25C條件下種子萌發,直至試驗結束,5個溫度梯度共萌發162粒狹葉香蒲種子,多組研究顯示25C條件下狹葉香蒲種子的萌芽程度最高。為精確其最佳發芽溫度,試驗組在23~28C之間再進行一次溫度試驗,最終得到狹葉香蒲的最佳萌發溫度為24C。此后試驗都在最佳溫度基礎上進行。
1.3試驗方法
1.3.1Cu2+脅迫對狹葉香蒲種子萌芽率的影響。將處理后的種子用20μmol/LNaCl0溶液浸泡6min,再用干凈自來水過濾,然后將其收集放人分好類的培養皿中,在培養皿中墊人濾紙,共分7組,分別加入0(CK)、50、100、150、200、300μmol/LCuSO4溶液,種子要被溶液完全覆蓋,其中對照組(CK)為不添加CuSO4溶液的等量蒸餾水。4次重復,等量放入100粒種子,然后將培養皿放人24C光照培養箱中,提供24h恒溫和12h光照時間。計算狹葉香蒲種子的發芽率,觀察幼苗生長響應總趨勢,發芽率計算公式為:發芽率(%)=(研究完成后健康萌發并生長正常的種子數/研究開始前培養皿中的種子數量)x100;種子活力(%)=(種子發芽20d生長健康的幼苗/研究中已經發芽的培養皿中的種子數量)10013。1.3.2狹葉香蒲幼苗在Cu2+脅迫條件下的生理響應。將正常發芽的狹葉香蒲幼苗用已消毒的鑷子夾人至生長箱中,記錄處理前后的根和葉的生長狀況,注入適量蒸餾水清洗9d(每3d為一個周期進行替換),然后轉至保溫箱中,用植物生長營養液進行培養,每24h更換1次培養液,待幼苗根長達到一定數量后,挑選出生長狀況相似的幼苗,等量放到0、250、500、750、1000μmol/LCu(NO3)2營養溶液中,每個梯度設4次重復。經過15d培養液浸泡后,用鑷子取出并將其均分成上部組織和根部組織,用蒸餾水洗凈后進行生理響應指標變化的記錄。
1.4數據分析
采用電子設備合并數據進行分析,對數據及差異性進行分析。
2結果與分析
2.1狹葉香蒲種子誘發癥狀在Cu2t脅迫下的響應
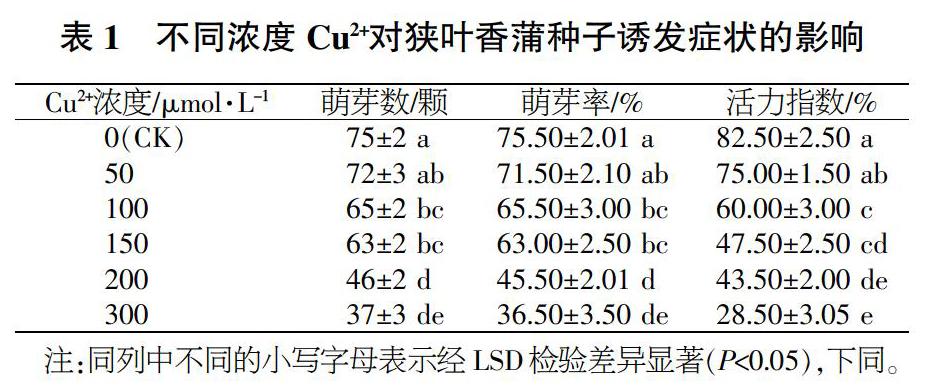
不同濃度Cu2脅迫對狹葉香蒲種子的誘發情況影響如表1所示。可以看出,在Cu2脅迫狀態下,隨著Cu2+含量不斷提高,狹葉香蒲種子的萌發情況、萌發趨勢、萌發癥狀和生命活性都逐漸下降4。在50μumol/LCu2+的脅迫條件下,種子的萌芽率、萌芽數.種子活性系數與對照組不存在差異;當Cu2+)濃度達150μmol/L時,種子的萌芽率、萌芽數、種子活性系數均明顯低于對照,其值分別較對照降低了16.6%、16.0%和42.4%,說明在150μumol/LCu2脅迫條件下狹葉香蒲種子的萌發受到了一定的抑制;當Cu2+濃度達到300μmol/L時,種子的萌芽率、萌芽數、種子活性系數很明顯低于對照組且降低幅度大,分別比對照降低了51.7%、50.7%、65.5%。綜上所述,隨著Cu2+濃度逐漸增高,對狹葉香蒲種子萌發的抑制作用越大。
2.2Cu2+脅迫對狹葉香蒲幼苗生理特性的影響
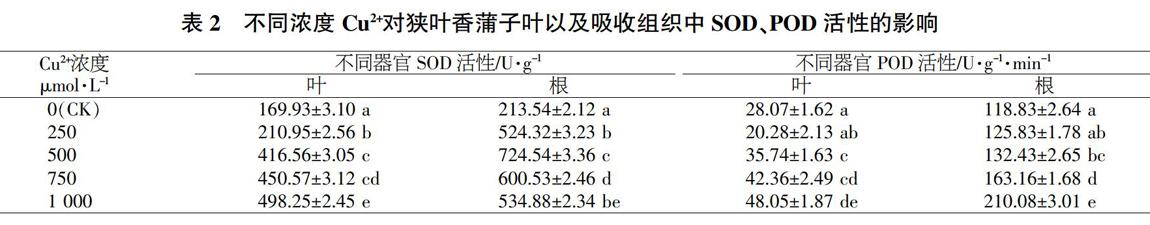
2.2.1對狹葉香蒲幼苗葉片以及根中SOD、POD活性的影響。在不同濃度Cu2+脅迫條件下,狹葉香蒲幼苗中葉片、根中SOD與POD活性的響應見表2。可以看出,在不同濃度Cu2+脅迫條件下,狹葉香蒲葉片、根中SOD活性均顯著高于對照且差異顯著,但兩者變化趨勢各不相同。葉片SOD活性隨著Cu2+濃度變大波動,但整體呈上升的趨勢,其中在250~500pumol/L濃度下波動較大,在500~1000μmol/L濃度下增幅較小,在1000μmol/LCu2脅迫條件下達到最高,為對照的2.93倍。根中SOD活性則隨著Cu2+濃度變大表現出先上升后下降的趨式,Cu2+脅迫下波動最高的濃度是500μmol/L,SOD活性達到724.54U/g,為對照的3.39倍;在1000μmol/LCu2+脅迫條件下,SOD活性為534.88U/g,為對照的2.50倍。
不同濃度Cu2+脅迫條件下,狹葉香蒲的葉片和根中POD活性隨著Cu2+濃度的增加而,上升。在250μmol/LCu2+脅迫條件~下葉片POD活性略低于對照,在500μmol/LCu2+脅迫條件下與對照有顯著差異;而在750、1000μmol/LCu2+脅迫條件下葉片POD活性顯著高于對照組,分別為對照組的1.51倍和1.71倍。
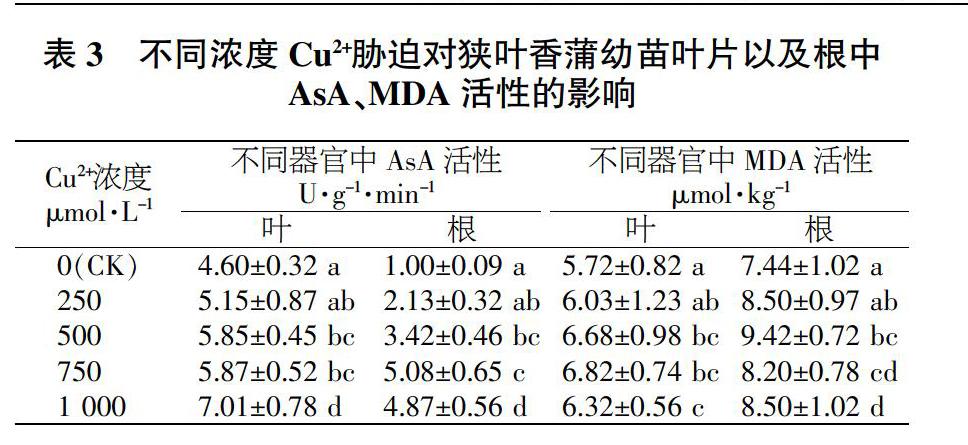
2.2.2對狹葉香蒲幼苗葉片以及根中AsA、MDA的影響。在不同濃度Cu2脅迫下,狹葉香蒲幼苗子葉以及根中的AsA和MDA的響應情況如表3表示。可以看出,不同濃度Cu2+脅迫情況下狹葉香蒲子葉以及根中的AsA顯著高于對照,葉片中的AsA活性都表現出上升趨勢,但在250μmol/LCu2+脅迫條件下與對照組差異不明顯;而在1000μmol/LCu2+脅迫條件下,葉片中的AsA活性與對照組存在著很明顯的差異,為對照組的1.52倍。根中AsA的活性與對照組存在明顯差異,并隨著Cu2+濃度的上升呈先上升而后趨于平穩的趨勢,根中AsA的活性在750μmol/LCu2+脅迫條件下,為對照組的5.08倍。
在Cu2+脅迫影響下,狹葉香蒲幼苗子葉和根中MDA活性與對照組差異不明顯,僅僅只是波動變化和波動趨勢存在微小差異。葉片中MDA活性隨著Cu2+濃度變大表現出先上升后稍有下降的波動趨式,在750μmol/LCu2+脅迫條件下最高。研究發現,葉片中的MDA與對照組差異不顯著,說明不同濃度Cu2+脅迫對狹葉香蒲幼苗葉片的質膜基本不存在過氧化破壞。根中的MDA活性隨著Cu2+含量上升呈現高一低一高的波動變化,在500μmol/LCu2時活性最大,是對照組的1.27倍;但是其他處理根中的MDA活性與對照組沒有很明顯的差別,很有可能是較低濃度Cu2+脅迫時狹葉香蒲幼苗產生的抗逆性而表現出MDA活性上升。
3結論與討論
試驗結果表明,隨著Cu2+濃度逐漸上升,狹葉香蒲種子的萌發率、萌發趨勢、萌發癥狀和生命活性均呈下降趨勢,這與試驗之前所收集到的文獻結論基本一致。該試驗中,在Cu2脅迫濃度較低(50~150μmol/L)的情況下,狹葉香蒲種子的發芽癥狀以及小苗的生理響應比對照組低,顯然,濃度偏低的Cu2脅迫對狹葉香蒲種子的萌發及其幼苗的生理響應均存在著不同程度的影響,但其發芽率仍然達到了63%,這也說明狹葉香蒲能在較低的Cu2+脅迫環境下萌發、生長;在相對高濃度Cu2+(150~300μmol/L)時,狹葉香蒲種子的發芽率和幼苗的生理響應情況都明顯低于對照組,在濃度高達300μmol/L情況下發芽率僅有36.5%,比對照組降低了51.7%;萌發的種子活力也只有28.05%。由此可見,濃度高的Cu2+脅迫對狹葉香蒲種子的發芽率和幼苗生理響應具抑制作用,這與王穗子等對Cu脅迫條件下AMF對海洲香薷光合色素含量、抗氧化能力和膜脂過氧化的影響的相關試驗結果一致。
超氧化物歧化酶(SOD)和過氧化物酶(POD)是綠色植物體內抗氧化組織的重要組成部分,其具有可抑制活性氧生成、消滅超氧陰離子自由基限制脂質過氧化和減低細胞膜系統受傷等功能15-17。在不同濃度Cu2+脅迫影響下,狹葉香蒲幼苗子葉以及吸收組織SOD活性都表現出比空白對照組高的現象,這充分表明Cu2+脅迫可以增加狹葉香蒲體內細胞消除超氧陰離子自由基的功效,植物通過自我修復能在一定程度上降低細胞被毒害的程度,加強植株對Cu2+的抵抗力。不同濃度Cu2+脅迫對狹葉香蒲根部和葉SOD活性的影響研究表明,狹葉香蒲根部對Cu2+的敏感度強于葉。在750、I000umol/LCu2t離子脅迫影響下,狹葉香蒲葉片的POD分別是對照組的1.51倍和1.71倍,而狹葉香蒲根部的POD分別是對照組的1.37、1.77倍。可見濃度較高的Cu2+脅迫可以引起狹葉香蒲植株體內POD活性的上升且根系對Cut脅迫的敏感度強于葉。
抗壞血酸(AsA)是植物細胞中的重要抗氧化劑。隨Cu2+脅迫濃度的不斷增高,狹葉香蒲植株中葉內AsA呈現增高趨勢,但只有在1000μumol/L濃度下增高顯著高于對照,根內AsA更為顯著地表現出增高趨勢。這表明低濃度Cu2對植株內細胞存在著一定的危害,最大的傷害是對根部的傷害,葉對Cu2脅迫的敏感程度小于根系。
丙二醛(MDA)是環境污染情況下植物中膜脂過氧化作用的產物,其在植物體中的沉積含量可體現出膜脂過氧化。當Cu2脅迫濃度逐漸變大時,狹葉香蒲植株中葉內MDA含量有升高趨勢但并不顯著。根部呈現了高一低一高的趨勢,但是只有在500umol/LCu2脅迫下顯著高于對照組,由此推測原因可能是低濃度的條件下根系有應激反應,以致影響了MDA含量上升。
綜上所述,在低濃度Cu2+脅迫條件下狹葉香蒲仍然具備相對較高的發芽率,說明該植株可以在輕度或中度的Cu2+污染水域中萌發和生長;在同一梯度Cu2*脅迫下,狹葉香蒲植株葉內的SODPOD活性以及MADAsA比根部小,這可能是由于根部是水體和植株直接接觸的位置,屬直接的受害器官叫。試驗結果表明,狹葉香蒲能夠通過根部和葉吸收部分Cu2+,以減少水體中Cu2t的含量,進而起到凈化水體的作用,可用于治理和修復輕度中度水體污染。
4參考文獻
[1]呂篤康,李海冰,巴音山,等.重金屬鉻及鹽脅迫對小香蒲種子萌發的影響[J].2013(2):653-761.
[2]張曉斌植物修復在水環境污染治理中的研究[D].杭州:浙江師范大學,2007.
[3]袁宜如,李曉云,高光林.狹葉香蒲及其開發利用研究進展[J]現代農業科技2012(9):12-113.
[4]中國科學院中國植物志編輯委員會中國植物志([M].北京:科學出版社,2004.
[5]張開明,佟海英,黃蘇珍,等.Cu脅迫對黃菖蒲和馬藺Cu富集及其他營養元素吸收的影響[J].植物資源與環境學報,2007(1):18-22.
[6]劉素純,蕭浪濤,廖柏寒,等鉛脅迫對黃瓜幼苗抗氧化酶活性及同工酶的影響[J].應用生態學報,2006,17(2):300-304.
[7] ZHANG S, HU J, CHENZ H, et al. Effecta of Pb pollution on seed vigor of
three rice cultivars[J]. Rice Science, 2005(3): 197-202.
[8]趙艷,徐迎春,吳曉麗,等.Cu脅迫對狹葉香蒲體內元素吸收分配的影響[J].生態學雜志,2009(4);65-670.
9]李鋒民,熊志廷,胡洪營海洲香薷對銅的蓄積及銅的毒性效應[J].環境科學,2003,24(3):30-34.
[10]廖斌,鄧冬梅,楊兵,等.鴨跖草(Commelina communis)對銅的耐性和積累研究[J].中山大學學報(自然科學版),2004,43(2):72-75.
[11]張弛,袁亞光,欽佩,等.香蒲對重金屬鎘的耐性及吸收途徑研究[J].南京大學學報(自然科學版),2013,49(4):506-511.
[12]趙艷,徐迎春,柴翠翠,等銅脅迫對狹葉香蒲生長及生理特性的影響[J]廣西植物,2010,30(3):367-372.
[13]孟煥,王雪宏,佟守正,等濕地土壤深埋對蘆葦.香蒲種子萌發的影響[J]生態學雜志,2013,32(9):2877-2957.
[14]徐金波,徐迎春,趙慧,等.Pb脅迫條件下狹葉香蒲種子的萌發特征及其幼苗的生理響應[J].植物資源與環境學報,2014,23(3):3302-3343.
[15]吳學明,蘇旭,劉玉萍.青海湖畔兩種植物葉片中超氧化物歧化酶(SOD)和過氧化物酶(POD)活性的研究[J].青海草業,2005,14(2):7-1.
[16]謝傳俊,楊集輝,周守標,等鉛遞進脅迫對假儉草和結縷草生理特性的影響[J].草業學報,2008,17(4):65-70.
[17]周希琴,莫燦坤.植物重金屬脅迫及其抗氧化系統[J].新疆教育學院學報,2003(2):103-108.
[18]孫天國,沙偉,劉巖.復合重金屬脅迫對兩種蘚類植物生理特性的影響[J].生態學報,2010,30(9):2332-2339.
[19]姚鵬現代生物技術在環境污染治理中的應用探討0].黑龍江科技信息,2017(16):169-170.
[20]王小玲,劉騰云,幸學俊,等硅對Cd.Pb.Cu.Zn正交脅迫下水稻丙二醛含量的影響J]湖北農業科學,2016,55(22):5771-5776.
[21]馬春花,馬鋒旺,李明軍,等.外源抗壞血酸對離體蘋果葉片衰老的影響[D.武漢植物研究,2006,33(6):5480-5498.
