酵母肽體重控制代餐粉調節糖脂代謝功能評價
2019-09-10 07:22:44趙可心段盛林韓詩雯潘聰于偉厚夏凱
中國食物與營養 2019年5期
趙可心 段盛林 韓詩雯 潘聰 于偉厚 夏凱


摘 要:目的:評價酵母肽體重控制代餐粉對于調節糖脂代謝的作用。方法:用酵母肽以及多種谷物粉配制體重控制代餐粉,采用體外消化的方法測定其GI值。提取得代餐粉水提物,得率為42.97%。將所得的水提物作用于游離脂肪酸誘導的胰島素抵抗HepG2細胞,采用MTT實驗檢測其對細胞的毒性,觀察HepG2細胞上清葡萄糖、胞內糖原、胞內TG、胞內TC以及油紅O染色的變化情況。結果:代餐粉的血糖生成指數(Glycemic Index,GI)為44.23,為低GI食品;其水提物在100~500μg/mL時對細胞的增殖沒有影響;與對照組相比,模型組細胞內糖原的含量顯著降低,上清葡萄糖,胞內TG、TC的含量均顯著升高,代餐粉處理后胞內糖原的含量明顯上升,上清葡萄糖,胞內TG、TC的含量均顯著下降;油紅O染色結果說明,模型組細胞內出現了大量小油滴,說明模型組出現脂代謝異常,而二甲雙胍和代餐粉處理后,紅色區域明顯減少,且500μg/mL代餐粉處理組效果優于300μg/mL。結論:酵母肽體重控制代餐粉可以減緩葡萄糖的釋放,調節HepG2細胞的糖脂代謝,減輕HepG2胰島素抵抗。
關鍵詞:酵母肽;代餐粉;血糖生成指數;糖脂代謝
酵母肽是以優質鮮酵母為原料,利用多種酶降解酵母蛋白,破除酵母壁并從酶解體系中分離純化而制成的小分子多肽[1],其氨基酸種類齊全,比例更接近于聯合國糧農組織和世界衛生組織推薦的理想蛋白質的氨基酸組成比例,是優質的蛋白質來源[2]。代餐粉是一種由谷類、豆類、薯類食材等為主,其他植物的根、莖、果實等可食用部分為輔制成的一種單一或綜合性沖調粉劑產品。因此,以酵母肽為主,輔以谷物粉、紫薯粉制備而成的營養代餐粉,將為快節奏高強度生活下追求健康生活的人們提供更便利的飲食和均衡的營養。
低血糖生成指數(Glycemic Index,GI)食物進入腸道后停留的時間長,釋放緩慢,使得血液中葡萄糖峰值較低,引起餐后血糖反應較小,需要的胰島素也相應減少,避免了血糖的劇烈波動,所以能夠有效控制血糖,既可以預防高血糖也能夠預防低血糖[3-6]。同時低GI膳食能降低血脂,改善胰島素抵抗,其原因是血液游離脂肪酸水平在進食低GI膳食后被抑制,同時拮抗激素的反應,進而使外周組織對葡萄糖的攝取利用率增加[7]。因此,低GI膳食可以調節糖脂代謝,達到輕脂減重和改善胰島素抵抗的效果。
本研究中酵母肽體重控制代餐粉中含有高膳食纖維的紫甘薯粉和木耳粉,脂肪代謝促進劑左旋肉堿,優質蛋白酵母肽以及各種谷物粉。研究表明,血清高濃度左旋肉堿可改善胰島素抵抗,促進糖脂代謝,對體重控制起輔助作用[8]。從配方來講,該代餐粉不僅營養全面,飽腹感強,還能調節糖脂代謝,控制體重。本研究評價了酵母肽體重控制代餐粉對胰島素抵抗HepG2細胞糖脂代謝的影響。
1 材料與方法
1.1 材料與試劑
酵母肽體重控制代餐粉,中國食品發酵工業研究院自行研發;白面包,市場購買;人肝癌細胞株(HepG2),北京協和細胞中心;DMEM培養基、磷酸鹽緩沖液(PBS)、DMEM無糖培養基、小牛血清,美國Gibco;油酸(OA)、棕櫚酸(PA)、DCFH-DA、二甲基亞砜(DMSO)、噻唑蘭(MTT)、油紅O染色劑、胰蛋白酶、胃蛋白酶、淀粉轉葡萄糖苷酶和胰酶,美國Sigma化學公司;無FFA牛血清蛋白(BSA),日本WAKO公司;RIPA裂解液、TG試劑盒、葡萄糖試劑盒,碧云天生物技術研究所;乙腈、甲醇,均為色譜純;瓜爾豆膠,北京化工廠;95%乙醇、磷酸二氫鈉、磷酸氫二鈉、鹽酸、氫氧化鈉,國產分析純。
1.2 儀器與設備
高效液相色譜儀,日本島津公司;倒置生物顯微鏡,日本OLympus公司;生物安全柜,中國濟南鑫貝西生物技術有限公司;細胞培養箱,日本松下公司;Spectra Max i3酶標儀,美國Molecular Devices公司;分析天平,梅特勒-托利多儀器有限公司;超聲波清洗儀,昆山超聲儀器有限公司;pH計,上海雷磁儀器廠;GL-20G-Ⅱ型高速冷凍離心機,上海安亭科學儀器廠;DK-8D三溫三控水槽,上海博迅實業有限公司。
1.3 方法
1.3.1 體外模擬消化測定酵母肽體重控制代餐粉的GI值 (1)口腔模擬[9]:取含1 g可利用的碳水化合物的各樣品組放入一燒杯中,加入 3mL 0.1mol/L的磷酸緩沖液和1mL預先加熱到37℃的淀粉酶,用研缽棒輕輕上下敲打15次后(或15s)(保持在37℃的環境中)。(2)胃部模擬:即用4mL 0.1mol/L的磷酸緩沖液沖洗研缽棒,保證研缽棒上的樣品都轉移至燒杯中,加入6mL含有0.4g/L的NaCl、0.05g胃蛋白酶和0.05g瓜爾豆膠的0.1mol/L磷酸緩沖液,利用2mol/L HCl溶液調到pH1.5,放入3~5顆玻璃球,然后放在37℃搖床中保溫30min。(3)小腸模擬[10]:將10 mL磷酸緩沖液(pH 6.9,0.5 mol/L)加入到上述溶液中,利用50%的NaOH溶液調節pH為6.9;然后加入125 μL MgCl2-CaCl2溶液、125 μL 胰酶溶液、400 μL淀粉轉葡萄糖苷酶,補充蒸餾水至50 mL,在37℃搖床中保溫180 min。不同時間取1 mL樣品放入含有4 mL 95%的乙醇溶液中(預熱至60℃),沸水浴滅酶后自然冷卻,10 000 r/min離心10min,上清過 0.45 μm 微孔濾膜后進行高效液相色譜測定。(4)葡萄糖的測定:色譜柱:Hypersil NH2S氨基色譜柱(4.6×250 mm,5 μm)、柱溫40℃、流動相:乙腈∶水 70∶30(V/V)、流速1.0 mL/min、進樣量10 μL[11]。(5)計算方法:按照葡萄糖釋放量,計算碳水化合物水解率,繪制碳水化合物水解率和時間關系圖,模擬方程,計算曲線下面積。定義面包的水解率為100,從而計算樣品的水解率HI,按照GI與HI的關系式:GI=0.862HI+8.189計算樣品的GI[12-13]。
1.3.2 超聲法制備酵母肽體重控制代餐粉水提物 取酵母肽體重控制代餐粉10 g,置于燒杯中,加入250 mL 去離子水,料液比為1∶25,溫度為30 ℃;超聲波輔助提取1 h;提取液經真空抽濾得到澄清提取液①;在濾渣中繼續加入250 mL 去離子水,超聲波輔助提取1 h,真空抽濾得到澄清提取液②;合并提取液①和②,真空旋轉蒸發濃縮,濃縮后浸膏采用真空冷凍干燥,得到酵母肽體重控制代餐粉水提物[14],水提物得率為42.97%。
1.3.3 人肝癌細胞株HepG2的培養 人肝癌細胞株HepG2細胞培養于含有10%新生牛血清、青霉素100 U/mL和鏈霉素100 μg/mL的DMEM培養基中,于培養箱37℃、5% CO2培養條件下培養。選取對數生長期細胞進行實驗。將細胞以1×106個/mL濃度接種于96孔板,待細胞貼壁并長滿時,用PBS潤洗細胞1次,加入不同的處理液。實驗分組如下,對照組:25 mmol/L葡萄糖組(25G 1% BSA)、模型組:20 mmol/L葡萄糖+15 mmol/L果糖組(20G 15F 1 mmol/L FFA)組以及模型組中加入各濃度酵母肽體重控制代餐粉提取物[15]。
1.3.4 MTT法測定HepG2細胞存活率 取96孔培養板,每孔加100 μL細胞懸液,HepG2細胞濃度為1×105個/mL,37℃培養24 h后棄去培養液,用PBS清洗1次,每孔加入100μL100、200、300、400、500μg/mL的酵母肽體重控制代餐粉水提物干預24h后除去培養液,加入0.5 mg/mL MTT-DMEM于37℃避光孵育4 h,再加入100 μL DMSO,震蕩混勻以完全溶解出MTT紫色結晶產物。用酶標儀在490nm處測定吸光度值。以對照組細胞的細胞存活率為100%計算其余組別細胞存活率。
1.3.5 油紅O染色 油紅O是一種脂溶性偶氮染料,具有很強的脂溶性和染脂性,該染料可特異性的與組織內或細胞內的甘油三脂(TG)等中性脂肪結合成小滴狀而使脂類物質著色呈現橘紅色,同時又可以避免使細胞內的其他成分與油紅O發生反應而被其染色,以干擾實驗結果[16-18]。細胞加樣處理24 h后,棄掉舊培養基,用PBS緩沖液洗去殘余的細胞培養基,加入新鮮配制好的75%乙醇,室溫環境下固定細胞10min。再用PBS緩沖液洗去殘余的乙醇溶液,重復3次,然后加入新鮮配制的油紅O染液工作液,室溫下染色10 min。棄掉油紅O,用PBS緩沖液洗去殘余的油紅O,重復3次。將細胞培養皿置于倒置相差顯微鏡下觀察細胞中脂滴的形成。
1.3.6 糖代謝指標的檢測 細胞糖吸收測試:將對數期生長的細胞接于96孔板中,待細胞長滿后,棄去上清,PBS清洗1次,各組(每組設置3個平行)細胞加樣處理24 h后,小心棄去培養液,用PBS洗1次,換為12.5 mmol/L DMEM培養基,培養 24 h后取5μL上清液,用葡萄糖試劑盒測定葡萄糖含量。
細胞內糖原含量的檢測:將對數期生長的細胞接于12孔板中,待細胞長滿后,棄去上清,PBS清洗1次,各組(每組設置3個平行)細胞加樣處理24 h后,小心棄去培養液,PBS 洗1次,細胞用RIPA裂解液裂解,用糖原試劑盒測定胞內糖原含量。結果以模型組的百分比表示。
1.3.7 脂代謝指標的檢測 將對數期生長的細胞接于12孔板中,待細胞長滿后,棄去上清,PBS清洗1次,各組(每組設置3個平行)細胞加樣處理24 h后,小心棄去培養液,將板內待測定細胞用PBS潤洗2次后,加入RIPA裂解液。充分裂解后,用TG試劑盒測定TG含量,TC試劑盒測定TC含量。結果以模型組TG和TC含量的百分比表示(%)。
1.4 統計學處理
采用每組實驗重復3次,結果以X±SD表示。采用SPSS 23軟件進行單因素方差分析,來判斷顯著性差異,*P<0.05代表有顯著性差異,**P < 0.01代表有極顯著性差異。
2 結果與分析
2.1 酵母肽體重控制代餐粉的GI值
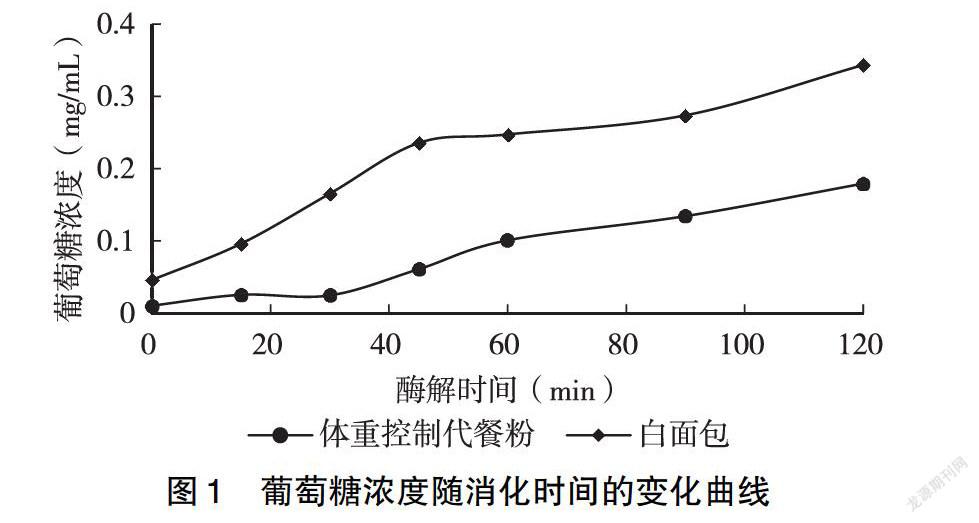
以葡萄糖濃度為橫坐標、峰面積為縱坐標,繪制葡萄糖標準曲線,進行線性回歸分析,得到回歸方程為Y=61 244X-183.7,r2=0.999(n=3),表明葡萄糖在0~2 mg/mL濃度范圍內線性良好。對體重控制代餐粉體外消化過程中葡萄糖的變化進行檢測。如圖1所示,隨著消化時間的延長,白面包和體重控制代餐粉酶解后的葡萄糖濃度都在不斷增加。相比白面包組來講,體重控制代餐粉酶解后葡萄糖濃度增加速度比較緩慢,對血糖濃度影響較小。經計算得知,白面包GI值為94.38,酵母肽體重控制代餐粉GI值為44.23,為低GI食品。結果表明,酵母肽體重控制代餐粉可以有效抑制因攝入可溶性碳水化合物而導致的血糖升高,有利于調節糖代謝。
2.2 MTT法測定HepG2細胞存活率
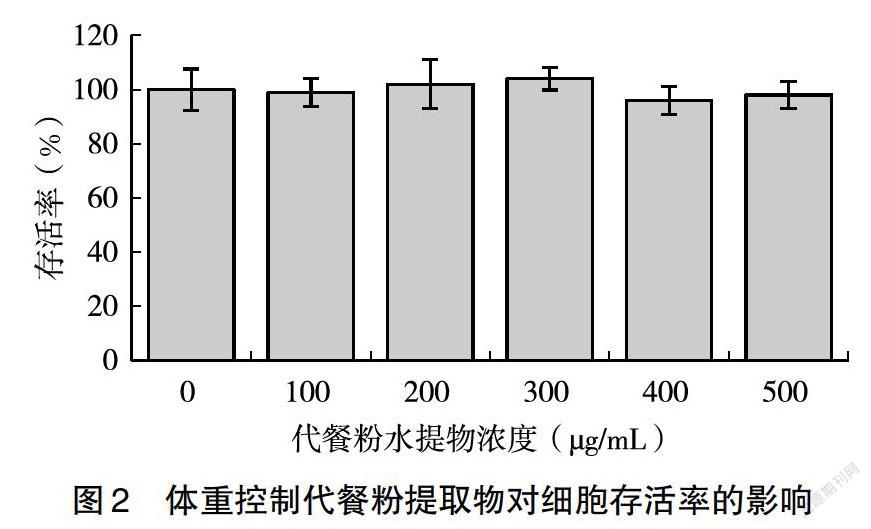
MTT法是一種檢測細胞存活和生長的方法,其檢測原理為活細胞線粒體中的琥珀酸脫氫酶能使外源性MTT還原為水不溶性的藍紫色結晶甲臜并沉積在細胞中,而死細胞無此功能[19]。二甲基亞砜(DMSO)能溶解細胞中的甲臜,用酶聯免疫檢測儀在540 nm波長處測定其光吸收值,可間接反映活細胞數量。酵母肽體重控制代餐粉水提物處理細胞24 h后,發現水提物濃度為0~500 μg/mL時對HepG2細胞增殖沒有明顯影響,說明該濃度的水提物對HepG2細胞無毒性,因此,后續細胞實驗可選用0~500 μg/mL的提取物對細胞進行處理(圖2)。
2.3 油紅O染色
油紅O染色結果表明,對照組細胞與模型組細胞的細胞形態出現明顯差異,大量模型組細胞被油紅O染成紅色,說明模型組細胞內發生脂質累積,出現小油滴,表明胰島素抵抗模型構建成功。陽性對照二雙胍組被染成紅色的區域較少,說明二甲雙胍具有明顯的促進脂代謝的作用,抑制脂質在細胞內積累。300 μg/mL和500 μg/mL的代餐粉提取物兩組的紅色油滴較少,500 μg/mL組紅色區域明顯少于300 μg/mL,且說明酵母肽體重控制代餐粉可以促進脂代謝,抑制脂質在細胞內積累且呈現劑量依賴性(圖3)。
2.4 酵母肽體重控制代餐粉對胰島素抵抗HepG2細胞糖代謝的影響
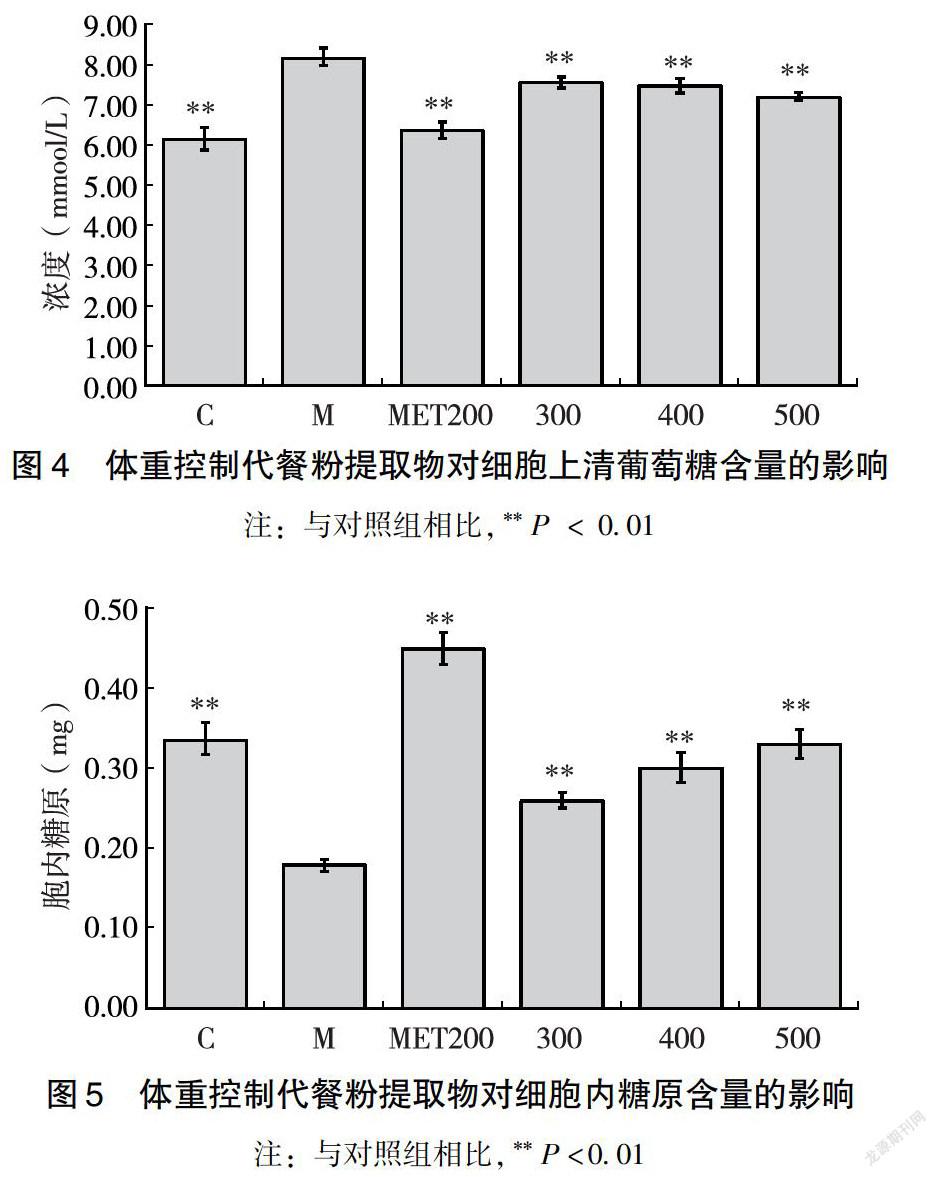
與對照組相比,模型組細胞上清葡萄糖的含量上升,細胞內糖原的含量下降,說明模型組細胞吸收葡萄糖的能力下降,糖原降解速度加快,肝糖異生明顯,說明FFA誘導后HepG2細胞內糖代謝異常。二甲雙胍處理后,細胞上清葡萄糖含量顯著下降,胞內糖原顯著上升(**P<0.01),說明二甲雙胍能夠促進細胞吸收葡萄糖,抑制細胞內糖原的降解。代餐粉水提物處理24 h后,上清葡萄糖含量明顯下降(**P <0.01),且與水提物濃度呈現負相關趨勢;胞內糖原的含量顯著上升(**P<0.01),且呈現劑量依賴性,表明體重控制代餐粉水提物可促進糖脂代謝紊亂HepG2細胞攝取葡萄糖,抑制細胞內肝糖原降解,從而改善胰島素抵抗。且代餐粉水提物改善胰島素抵抗HepG2細胞糖代謝的方式與陽性對照藥物二甲雙胍一致(圖4、圖5)。
2.5 酵母肽體重控制代餐粉對胰島素抵抗HepG2細胞脂代謝的影響
與對照組相比,模型組細胞內TG和TC的含量顯著上升(**P<0.01),說明FFAs誘導后,TG和TC在細胞內發生累積,細胞脂代謝異常。200μg/mL的二甲雙胍處理后,細胞內TG和TC的含量顯著低于模型組(**P<0.01),說明二甲雙胍能夠抑制胰島素抵抗HepG2細胞內TG和TC的積累,調節細胞的脂代謝。加樣處理24 h后,與模型組相比,隨著樣品濃度增加,胞內TG水平整體低于對照組,且400μg/mL組和500μg/mL組有顯著性差異(*P<0.05)。由圖7可知,加樣處理24 h后,與模型組相比,隨著樣品濃度增加,胞內TC水平整體顯著低于對照組(*P<0.05)。這表明體重控制代餐粉提取物可能通過促進胞內TG的釋放以及胞內脂肪代謝,從而減少了胞內TG的累積,其作用方式與陽性藥物二甲雙胍一致。且體重控制代餐粉提取物促進脂代謝的功能呈現一定的劑量依賴性(圖6、圖7)。
3 結論
本研究對Englyst[20]的體外消化方法進行完善,通過模擬食物的體內消化過程,確定了酵母肽體重控制代餐粉為低GI食品,代餐粉相對于白面包而言,葡萄糖釋放更加緩慢,不會導致人體血糖短時間迅速上升,從而起到控制糖代謝的作用。脂質過度蓄積(主要是TG)是造成肥胖、形成非酒精性脂肪肝、影響肝功能正常進行的先決因素[21]。與正常組細胞相比,模型組TG含量急劇上升且細胞形態發生變化,細胞輪廓不清晰,細胞內油脂滴多,細胞外有大量油圈,說明FFAs誘導后HepG2細胞已發生嚴重的細胞損傷且脂質蓄積過多。代餐粉水提物處理后,細胞內TG和總膽固醇(TC)的水平顯著降低,油紅O染色圖中代餐粉組小油滴的含量明顯減少,說明酵母肽體重控制代餐粉能夠抑制TG和TC在細胞內的積累從而維持脂代謝的平衡。
肝臟是糖代謝最主要器官之一,對維持血糖穩定有重要作用,主要表現在糖異生、糖原合成和攝取、利用及釋放葡萄糖。肝臟糖代謝異常產生胰島素抵抗,且為2型糖尿病基本病理生理現象[22-23]。因此,維持肝臟正常的糖代謝對機體具有重要意義。本研究發現,酵母肽體重控制代餐粉提取物能夠促進細胞吸收葡萄糖,減緩糖原的降解,從而維持正常的糖代謝平衡。
參考文獻
[1]梁天姣.殼寡糖的制備及其在酵母多肽分離純化上的應用研究[D]. 廣東湛江:廣東海洋大學,2015.
[2]江建梅,董連杰,陳冠青.酵母多肽在食品領域的應用及前景[J]. 食品安全導刊,2013(1):56-57.
[3]Jenkins DJ,Wolever TM,Taylor RH,et al.Glycemic index of foods:a physiological basis for carbohydrate exchange[J]. The American Journal of Clinical Nutrition,1981,34(3):362-366.
[4]The committee on food and nutrition of American diabetic association.Nutritional recommendations and principles for individuals with diabetes mellitus [J]. Diabetes Care,1991,14(25):20.
[5]Carbohydrates in human nutrition:Report of a Joint FAO/WHO Expert Consultation[M]. Food and Nutrition Paper,FAO 66.FAO Rome,1998.
[6]Granfeldt Y.Food factors affecting metabolic responses to cereal products[J]. University of Lund,1994.
[7]孫建琴,沈秀華,陳霞飛.食物血糖指數與糖尿病防治[J].腸外與腸內營養,2005,12(5):311-314.
[8]王詩琪,郭昆全.左旋肉毒堿與2型糖尿病[J]. 中國糖尿病雜志,2017(11):1051-1053.
[9]Englyst HN,Kingman SM,Cummings JH.Classification and measurement of nutritionally important starch fractions[J]. European Journal of Clinical Nutrition,1992(46):33-50.
[10]Granfeldt Y.Food factors affecting metabolic responses to cereal products[J]. University of Lund,1994.
[11]劉國玉.復合糖元模式研究及其在低GI特膳米中應用[D].中國食品發酵工業研究院,2016.
[12]楊月欣,崔紅梅,王巖,等.常見谷類和薯類的血糖生成指數[J]. 營養學報,2003,25(2):78-82.
[13]Foster-Powell K,Foster-Powell K,Miller JB.International tables of glycemic index[J]. The American Journal of Clinical Nutrition,1995,62(4):871-890.
[14]劉杰爾.松花粉成分分析及其輕身減肥功能初探[D].杭州:浙江大學,2008.
[15]柳嘉,郭孝萱,吳薇,等.高糖高脂誘導胰島素抵抗HepG2細胞模型的建立及活性成分的功能評價[J]. 食品科技,2012,37(3):73-78.
[16]鄭婧.肉桂多酚對 HepG2細胞脂質代謝的影響及其機制[J]. 山東醫藥,2017(1):38-40.
[17]歐露,麻燕妮,張彩平,等.煙酸通過下調 PCSK9的表達促進 HepG2細胞攝取LDL-C[J].中國藥理學通報,2017(2):243-248.
[18]李欣,王帥,周岳,等.羥基肉桂酸對HepG2和C2C12細胞糖脂代謝的調節作用[J]. 國際藥學研究雜志,2017(7):693-698.
[19]邊興艷.MTT比色法及其應用[J].國外醫學臨床生物化學與檢驗學分冊,1998,19(2):83-85.
[20]Englyst HN,Kingman SM,Cummings JH.Classification and measurement of nutritionally important starch fractions[J]. European Journal of Clinical Nutrition,1992(46):33-50.
[21]Cheung O,Sanyal A J.Abnormalities of lipid metabolism in nonalcoholic fatty liver disease[J]. Seminars in Liver Disease,2008,28(4):351-359.
[22]Stuart S,Andrzej W,Anil K J.Structural and functional organization of the animal fatty acid synthase[J]. Progress in Lipid Research,2003,42(4):289-317.
[23]Roncari D A.Abnormalities of adipose cells in massive obesity[J]. International Journal of Obesity,1990,14(3):187-192.
