穿山甲屬動(dòng)物的DNA條形碼及在穿山甲商品鑒定中的應(yīng)用△
2019-09-17 11:17:28李嬋謝雪那蔡炫張秋楠田恩偉晁志
中國(guó)現(xiàn)代中藥 2019年9期
關(guān)鍵詞:物種
李嬋,謝雪那,蔡炫,張秋楠,田恩偉,晁志
南方醫(yī)科大學(xué) 中醫(yī)藥學(xué)院,廣東 廣州 510515
穿山甲為鯪鯉科動(dòng)物穿山甲ManispentadactylaLinnaeus的鱗甲,始載于《名醫(yī)別錄》[1],具通經(jīng)下乳、消腫排膿、搜風(fēng)通絡(luò)之功,可用于經(jīng)閉癥瘕、乳汁不通、癰腫瘡毒、風(fēng)濕痹痛、中風(fēng)癱瘓、麻木拘攣[2]。現(xiàn)代藥理研究表明,穿山甲在降低血液黏度、升高白細(xì)胞、鎮(zhèn)痛、抗炎和促使炎癥修復(fù)等方面也有重要作用[3]。
穿山甲以白蟻為食,生活習(xí)性特殊,繁育困難,迄今未能成功實(shí)現(xiàn)人工養(yǎng)殖,供應(yīng)完全依賴野生資源。近二三十年來(lái),由于亂捕濫獵和棲息地的破壞,野生資源急劇減少,穿山甲藥材的供應(yīng)量大幅下降,價(jià)格日益增長(zhǎng),市場(chǎng)上出現(xiàn)從東南亞和非洲走私其他穿山甲屬物種的甲片替代入藥,以及多種其他摻假行為[4]。調(diào)查發(fā)現(xiàn),藥材市場(chǎng)上,中華穿山甲M.pentadactyla、馬來(lái)穿山甲M.javanica、印度穿山甲M.crassicaudata、南非穿山甲M.temminckii、大穿山甲M.gigantea、樹(shù)穿山甲M.tricuspis和長(zhǎng)尾穿山甲M.tetradactyla均有發(fā)現(xiàn)[5-6]。其他用于摻假的有豬、牛、羊的蹄甲等。為保證臨床用藥的安全有效,有必要對(duì)穿山甲的來(lái)源進(jìn)行準(zhǔn)確鑒定。
目前,過(guò)度捕獵和走私猖獗使穿山甲屬所有物種均瀕臨滅絕。世界自然保護(hù)聯(lián)盟《瀕危物種紅色名錄(2017)》將中華穿山甲和馬來(lái)穿山甲列為“極度瀕危”物種,將其他穿山甲物種列為“瀕危”或“易危”物種。2017年,8種穿山甲均被列入《瀕危野生動(dòng)植物種國(guó)際貿(mào)易公約》(CITES)附錄一。穿山甲的準(zhǔn)確鑒定也是保護(hù)穿山甲的必要條件。
目前,穿山甲常用的鑒別方法為性狀鑒別法,某些種類的穿山甲甲片的性狀極相似,即使有專業(yè)的技術(shù)與豐富的經(jīng)驗(yàn),也難以對(duì)其基原進(jìn)行準(zhǔn)確鑒定。另外有薄層色譜法、紅外光譜法、熒光分析法、離子色譜法、高效液相色譜法、高效毛細(xì)管電泳法等,這些方法多用在鑒別摻雜穿山甲、塑料制品、豬蹄甲等摻偽品[6-13]。與上述鑒別方法相比,DNA 分子技術(shù)不受樣品形態(tài)、數(shù)量等限制,對(duì)于其基原鑒定具有獨(dú)特的優(yōu)勢(shì),因此近年來(lái)有不少研究者開(kāi)始利用DNA分子鑒定法對(duì)穿山甲進(jìn)行研究,如尹艷等[14]建立了穿山甲特異性多聚酶鏈?zhǔn)椒磻?yīng)(PCR)鑒別方法,邢亞琳等[15]運(yùn)用cytb基因、12S rRNA基因、隨機(jī)擴(kuò)增多態(tài)性DNA標(biāo)記(RAPD)引物及微衛(wèi)星引物對(duì)穿山甲進(jìn)行鑒定等。
DNA條形碼是目前公認(rèn)可以對(duì)生物物種進(jìn)行準(zhǔn)確鑒定的技術(shù)。部分研究者應(yīng)用此方法對(duì)穿山甲進(jìn)行了研究,如邢亞琳[16]的DNA條形碼研究中對(duì)3種穿山甲進(jìn)行了分析;賈靜等[17]對(duì)中華穿山甲及混偽品豬、牛、羊進(jìn)行分析,但均缺乏對(duì)穿山甲屬所有種及混偽品的全面分析。鑒于我國(guó)藥材市場(chǎng)穿山甲來(lái)源復(fù)雜多樣,利用DNA條形碼技術(shù)鑒別穿山甲藥材的基原,了解其商品流通的情況,對(duì)加強(qiáng)其質(zhì)量控制具有一定的意義。因此,為進(jìn)一步充實(shí)穿山甲DNA條形碼鑒定的內(nèi)容,本研究基于COI序列探討DNA條形碼鑒定穿山甲屬及其他混偽品的可行性。
1 材料與方法
1.1 儀器
高速冷凍離心機(jī)(HC-3018R,安徽中科中佳科學(xué)儀器有限公司);電熱三用水箱(BOS2,北京市醫(yī)療設(shè)備廠);電泳儀(DYY-6C,北京市六一儀器廠);多功能暗箱式紫外分析儀(CBIO-UV6A1,北京賽百奧科技有限公司);分析天平(ALC-210.4,ACCULAB);自動(dòng)振蕩器(MX-S,SCILOGEX);PCR儀(2720,Applied Biosystem);Nanodrop分光光度計(jì)(ND2000C,Thermo Fisher Scientific);迷你筆型電磨筆(18 V,上海辰靈貿(mào)易有限公司)。
1.2 試劑
瓊脂糖(VetectM REAGENT grade Sigma-Aldrich,111860);無(wú)水乙醇(天津市大茂化學(xué)試劑廠,20180801);Proteinase K(TI ANGEN BIOTECH,R6830);血液/細(xì)胞/組織基因組DNA提取試劑盒(離心柱型)(TIANGEN BIOTECH,R6725);SuperRed/GelRed(Biosharp,68110160);D2000(TIANGEN BIOTECH,MD114);6×DNA loading buffer(TIANGEN BIOTECH,R6116);2×TaqPCR MasterMix(TIANGEN BIOTECH,R6525)。
1.3 材料
本研究收集了中華穿山甲及其混偽品共計(jì)11個(gè)物種38份樣品,并從NCBI上下載了37條序列(均經(jīng)質(zhì)量檢驗(yàn),以確定下載序列與物種對(duì)應(yīng)關(guān)系的準(zhǔn)確性[18-19]),具體信息詳見(jiàn)表1~2。中華穿山甲來(lái)自廣州市野生動(dòng)物救護(hù)中心。實(shí)驗(yàn)樣品均經(jīng)廣州市野生動(dòng)物救護(hù)中心鄒潔建工程師和作者鑒定。另外,從各地藥店及藥企收集了35份穿山甲生甲片商品,具體商品信息詳見(jiàn)表3。
1.4 方法
1.4.1 DNA提取 穿山甲甲片和蹄甲類偽品用無(wú)水乙醇浸泡12 h,每3 h將乙醇換一遍,同時(shí)通過(guò)振蕩器輕輕振蕩清洗;之后以去離子水重復(fù)以上步驟。在60 ℃烘干,在紫外燈下正反面分別各照射滅菌30 min。[16]然后用已滅菌的刀片刮去角質(zhì)層的外層污染,取內(nèi)層,用小型電磨機(jī)磨成細(xì)粉。取細(xì)粉約0.2 g,在裂解步驟中加500 μL GA、100 μL二硫蘇糖醇(DTT)[20]、50 μL蛋白酶K,56 ℃水浴過(guò)夜,12 000 r·min-1離心2 min后取上清液,其余步驟按照血液/細(xì)胞/組織基因組DNA提取試劑盒(TIANGEN)說(shuō)明書(shū)步驟提取樣品DNA。
舌頭及其他肌肉組織取約25 mg,按照血液/細(xì)胞/組織基因組DNA提取試劑盒(TIANGEN)說(shuō)明書(shū)步驟提取樣品DNA。

表1 穿山甲及其混偽品樣品信息
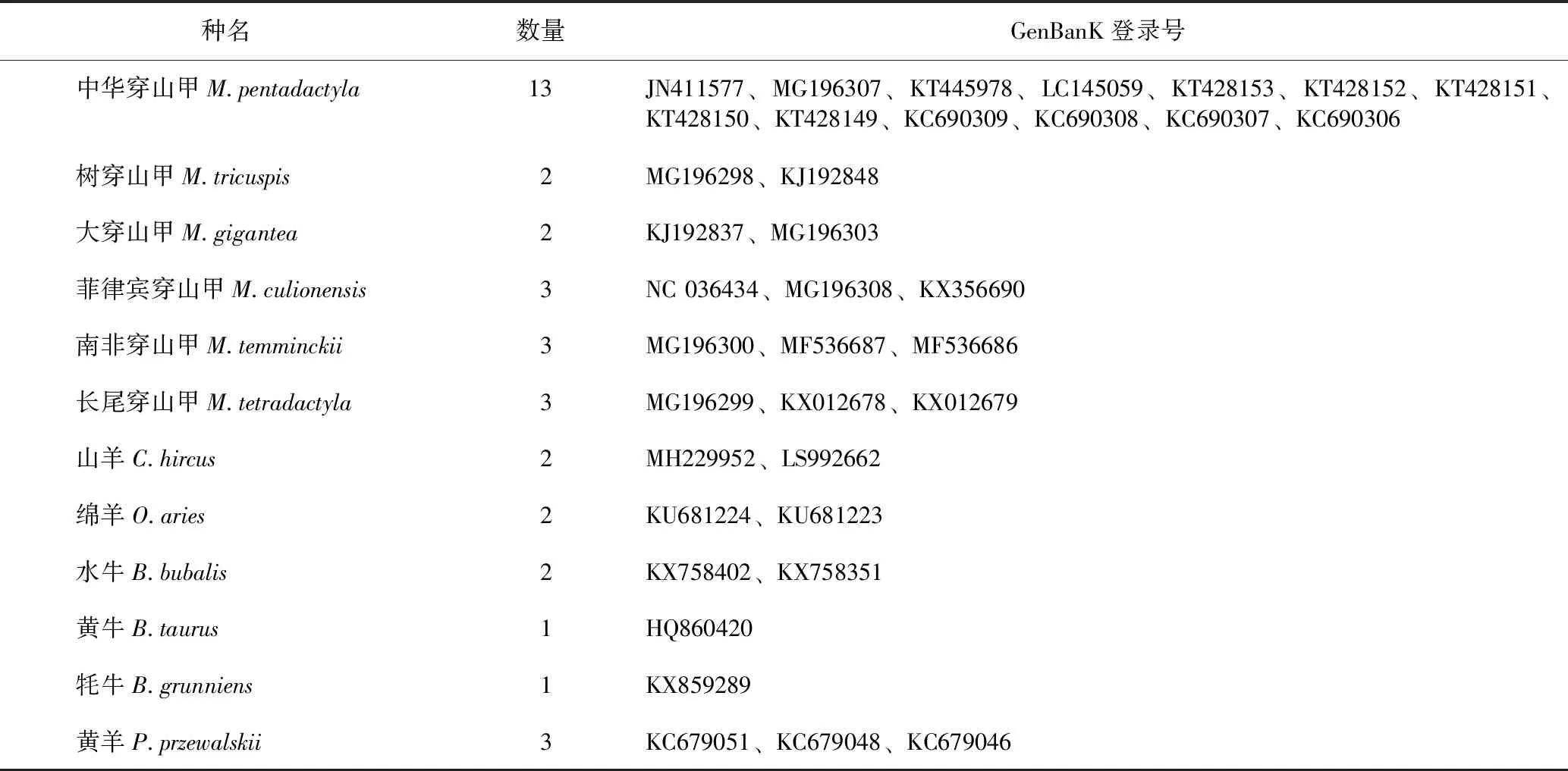
表2 GenBanK下載的穿山甲及其混偽品的序列信息

表3 穿山甲商品信息表
1.4.2 PCR擴(kuò)增及測(cè)序 采用COI序列通用引物MCOI-F(CAACAAATCACAAAGACATCGG)、MCOI-R(TTCTGGGTGTCCGAAAAATCA)[16],對(duì)提取的DNA進(jìn)行PCR擴(kuò)增。擴(kuò)增程序:95 ℃ 5 min;94 ℃ 30 s,50 ℃ 1 min,72 ℃ 1 min,35個(gè)循環(huán);72 ℃ 10 min。PCR反應(yīng)體積為15 μL,包含6 μL 2×TaqPCR Mix(TIANGEN),正反向引物各0.5 μL[生工生物工程(上海)股份有限公司],模板DNA 2 μL,ddH2O 6 μL補(bǔ)足體積至15 μL。實(shí)驗(yàn)中設(shè)置無(wú)DNA模板的空白對(duì)照組。將PCR擴(kuò)增產(chǎn)物送至上海美吉生物醫(yī)藥科技有限公司進(jìn)行雙向測(cè)序。
1.4.3 序列分析 用DNASTAR軟件中的Seqman對(duì)所得序列進(jìn)行拼接,去除引物區(qū),并對(duì)堿基進(jìn)行人工校對(duì)。將所有序列在MEGA(6.0版本)中對(duì)齊后,分析序列堿基組成和變異位點(diǎn)。
1.4.4 單倍型分析 用DNAsp v5軟件對(duì)COI序列進(jìn)行單倍型分析,并計(jì)算中華穿山甲的平均核苷酸多樣性(pi)[21],將中華穿山甲中數(shù)量最多的序列規(guī)定為核心單倍型,其他序列則為稀有單倍型。以正品核心單倍型為標(biāo)準(zhǔn)序列,以稀有單倍型和其之間的相似度為正品的相似度范圍,考察偽品所有單倍型與正品核心單倍型的相似度范圍,確定正品的相似度鑒別指標(biāo)。
1.4.5 種內(nèi)、種間遺傳距離分析及Barcode gap分析 將所有序列用MEGA軟件進(jìn)行K2P(Kimura 2-parameter)遺傳距離分析。并利用TAXON DNA/Species identifier軟件計(jì)算種內(nèi)及種間遺傳距離的分布頻率,并在Excel中畫(huà)出頻率分布直方圖,進(jìn)行條形碼間隙(barcode gap)分析[22]。
1.4.6 系統(tǒng)發(fā)育樹(shù)的聚類分析 在MEGA軟件中用鄰接法(neighbor joining method,NJ)構(gòu)建系統(tǒng)聚類樹(shù)。系統(tǒng)樹(shù)各分支的置信度用自舉檢驗(yàn)法(bootstrap test),檢驗(yàn)各分支的支持率,共進(jìn)行1000次循環(huán)。
1.4.7 ABGD劃分 ABGD(Automatic barcode gap discovery)基于遺傳距離對(duì)樣品進(jìn)行劃分,劃分在同一組的樣品被認(rèn)定為是一個(gè)種[23]。將樣品對(duì)齊的序列在線提交到ABGD網(wǎng)站(http://wwwabi.snv.jus-sieu.fr/public/abgd/abgdweb.htm),參數(shù)設(shè)置為:種內(nèi)差異先驗(yàn)值(prior intraspecific divergence)P為0.001到0.1,最小相對(duì)gap寬度值(Minimum relative gap width)X為1.0(K80)。
1.4.8 商品的鑒定 基于穿山甲及其混偽品的COI序列的單倍型信息及物種信息建立比對(duì)數(shù)據(jù)庫(kù),穿山甲商品序列可通過(guò)DNAMAN軟件與自建穿山甲條形碼數(shù)據(jù)庫(kù)中的序列進(jìn)行比對(duì),從而確定商品的物種。通過(guò)鄰接法與數(shù)據(jù)庫(kù)的序列構(gòu)建系統(tǒng)聚類樹(shù),系統(tǒng)樹(shù)各分支的置信度用自舉檢驗(yàn)法(bootstrap test),檢驗(yàn)各分支的支持率,共進(jìn)行1000次循環(huán)。并用ABGD劃分法對(duì)商品序列與數(shù)據(jù)庫(kù)序列一起進(jìn)行劃分,參數(shù)設(shè)置與1.2.7一致。
2 結(jié)果
2.1 DNA條形碼特征
2.1.1 COI序列堿基組成和變異位點(diǎn) 測(cè)序結(jié)果拼接且去除引物后得到658 bp的序列,所有序列對(duì)齊后長(zhǎng)度為600 bp,其中變異位點(diǎn)有232個(gè),堿基T、C、A、G平均含量分別為28.9%、27.6%、26.8%、16.7%。其中A+T含量(55.7%)明顯高于G+C含量(44.3%),核苷酸堿基構(gòu)成的偏倚是線粒體蛋白質(zhì)編碼基因的一個(gè)顯著特征。
2.1.2 單倍型分析 穿山甲及其混偽品的75條600 bp的COI序列,分為了45種單倍型。中華穿山甲不同來(lái)源樣品序列有16條,種內(nèi)變異位點(diǎn)有12個(gè),GC含量為0.44~0.45,共獲得6種單倍型(Hap1~Hap6)。中華穿山甲COI基因片段的平均核苷酸多樣性(pi)為0.003 78,單倍型多樣性(Hd)為0.743。COI基因片段中,正品的稀有單倍型與其核心單倍型之間的相似度為98.83%~99.67%,而偽品所有單倍型與正品核心單倍型的相似度為79.50%~95.17%。表4和圖1顯示了中華穿山甲的單倍型類型及核心單倍型的序列。

表4 中華穿山甲單倍型類型

圖1 中華穿山甲核心單倍型(Hap2)序列
2.1.3 種內(nèi)及種間遺傳距離分析 穿山甲及其混偽品的種內(nèi)遺傳距離為0~0.012,種內(nèi)平均遺傳距離為0.003 7,最大值出現(xiàn)在南非穿山甲Manistemminckii。種間遺傳距離值為0.044~0.334,種間差異凈值的最小值出現(xiàn)在馬來(lái)穿山甲M.javanica與菲律賓穿山甲M.culionensis之間,為0.044;最大值出現(xiàn)在豬Susscrofadomesticus與黃羊Procapraprzewalskii之間,達(dá)到了0.334。種間平均遺傳距離為0.255,遠(yuǎn)大于Hebert所推薦的物種鑒定最小種間遺傳距離0.020[24-25]。種間平均遺傳距離約為種內(nèi)平均遺傳距離的69倍,說(shuō)明COI序列能夠?qū)Υ┥郊准捌浠靷纹愤M(jìn)行有效的鑒定。
2.1.4 Barcode Gap檢驗(yàn) Barcode gap檢驗(yàn)結(jié)果(見(jiàn)圖2)顯示,穿山甲及其混偽品的種內(nèi)遺傳距離為0%~2%,種間遺傳距離>3%,存在GAP區(qū),即種內(nèi)遺傳距離最大值<種間遺傳距離最小值,說(shuō)明穿山甲及其混偽品的COI序列具有較高的種內(nèi)穩(wěn)定性和種間差異性,具有一定的遺傳間隔。
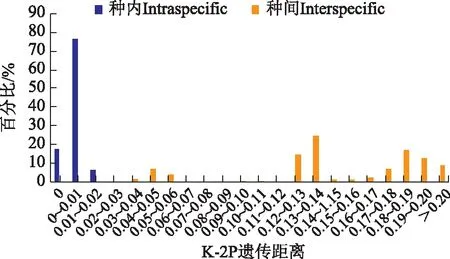
圖2 種內(nèi)和種間遺傳距離的分布
2.1.5 系統(tǒng)發(fā)育樹(shù)的聚類分析 由鄰接法構(gòu)建的系統(tǒng)聚類樹(shù)(見(jiàn)圖3)可知,中華穿山甲與其他混偽品均單獨(dú)聚為一支,支持率均大于95%,各物種可以區(qū)分開(kāi)。其中,這75條序列分成了2大支系,為穿山甲屬和其他混偽品。穿山甲屬又分成了2個(gè)支系:第一個(gè)支系為產(chǎn)于亞洲的種類,即馬來(lái)穿山甲、菲律賓穿山甲、印度穿山甲、中華穿山甲,這個(gè)支系中又分為2個(gè)支系,馬來(lái)穿山甲與菲律賓穿山甲聚為一支,印度穿山甲與中華穿山甲聚為一支。第二個(gè)支系為產(chǎn)于非洲的大穿山甲、南非穿山甲、樹(shù)穿山甲、長(zhǎng)尾穿山甲,在這個(gè)支系中也出現(xiàn)2個(gè)分支,大穿山甲、南非穿山甲為一分支,樹(shù)穿山甲、長(zhǎng)尾穿山甲為另一分支。
2.1.6 基于距離的分類分析 用ABGD對(duì)75個(gè)樣品進(jìn)行劃分,結(jié)果包括初始劃分和遞歸劃分2種情況(見(jiàn)圖4):初始劃分較穩(wěn)定,75個(gè)樣品被分成12組。遞歸劃分把75個(gè)樣品分成1~31組;P值介于0.01~0.03時(shí),75個(gè)樣品被穩(wěn)定地劃分成15個(gè)組;這15個(gè)分組分別代表了研究的15個(gè)物種,與聚類分析結(jié)果呈一一對(duì)應(yīng)關(guān)系(見(jiàn)圖3)。
2.2 穿山甲商品的鑒定
35批穿山甲甲片商品COI序列與自建的穿山甲條形碼數(shù)據(jù)庫(kù)比對(duì)結(jié)果見(jiàn)表5。在商品序列與自建數(shù)據(jù)庫(kù)的序列通過(guò)鄰接法構(gòu)建的系統(tǒng)聚類樹(shù)中,A114、B103與中華穿山甲聚為一支;A101~A113、A115、A116、A130與印度穿山甲聚為一支;A117~A128、B101、B104與馬來(lái)穿山甲聚為一支;A129、B102與大穿山甲聚為一支;A131與樹(shù)穿山甲聚為一支。同樣地,ABGD遞歸劃分將35個(gè)樣品分別劃分進(jìn)中華穿山甲組(A114、B103)、馬來(lái)穿山甲組(A117~A128、B101、B104)、印度穿山甲組(A101~A113、A115、A116、A130)、大穿山甲組(A129、B102)和樹(shù)穿山甲組(A131)。3種方法結(jié)果一致。

圖3 基于COI序列構(gòu)建中華穿山甲及其混偽品的NJ樹(shù)及ABGD遞歸劃分結(jié)果
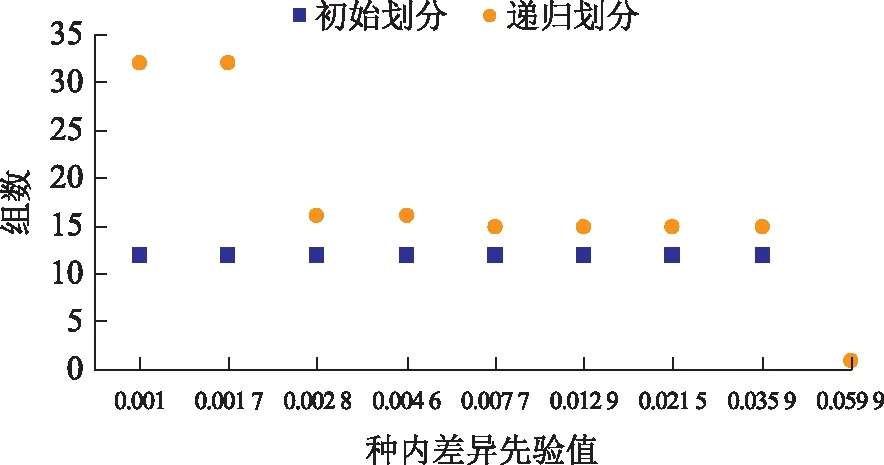
圖4 75個(gè)樣品的ABGD劃分結(jié)果
綜合以上結(jié)果可知,35份穿山甲商品中,僅有A114與B103為中華穿山甲(5.7%),其他樣品均為偽品。其中,以印度穿山甲(45.7%)、馬來(lái)穿山甲(40%)為最多。
2.3 印度穿山甲甲片性狀特征
印度穿山甲甲片(見(jiàn)圖5)因生長(zhǎng)部位不同,形狀大小也不一致。一般多呈扇面形,長(zhǎng)1.8~6.2 cm,寬2.1~6 cm,厚1.76~4.66 mm。甲片外表呈褐色。背面寬端較厚,窄端較薄,中央較厚,邊緣較薄。邊緣圓滑整齊或被磨損,有的有皮肉殘留。背面有多數(shù)縱向排列的線紋及在靠近頂端邊緣的地方會(huì)有數(shù)條橫線紋,背面的窄端較光滑,常有零亂與砂石摩擦的痕跡。內(nèi)表面色較淺,中部有一條明顯突起的弓形橫向棱線,其下方有數(shù)條與棱線相平行的細(xì)紋,偶有殘留皮脂飛邊或毛,縱向凸起紋理的面向上隆起呈弧形。角質(zhì)質(zhì)地較輕(平均密度為1.57 g·cm-3),半透明。氣微腥,味淡。

表5 35批穿山甲甲片商品比對(duì)結(jié)果

圖5 印度穿山甲甲片
3 討論
成功DNA提取是分子鑒定的基礎(chǔ)。穿山甲甲片及混偽品蹄甲為高度角質(zhì)化的材料,一般方法難以提取DNA。在本研究中,曾嘗試用結(jié)合酶消化的酚-三氯甲烷法提取[24],耗時(shí)近一周;而用DTT結(jié)合試劑盒[26]的方法提取,僅需1 d即可提取成功。前者提取的DNA濃度高于后者,但因后者效率更高且擴(kuò)增效率也好,最終選擇DTT結(jié)合試劑盒的方法進(jìn)行提取。在DNA提取之前,甲片及蹄甲應(yīng)用無(wú)水乙醇浸泡消毒,再用無(wú)菌水浸泡,充分烘干后在紫外燈下照射消毒。在取樣時(shí),需刮去甲片及蹄甲表層,取內(nèi)層,否則容易出現(xiàn)外源DNA的污染的情況,影響測(cè)序結(jié)果甚至出現(xiàn)擴(kuò)增出菌的現(xiàn)象。
本研究利用COI通用引物對(duì)穿山甲及其混偽品的DNA進(jìn)行擴(kuò)增,均得到了100%的擴(kuò)增效率與測(cè)序效率。采用Kimura-2-parameter模型計(jì)算遺傳距離,發(fā)現(xiàn)種間平均遺傳距離為種內(nèi)平均遺傳距離的69倍,滿足種間差異是種內(nèi)差異的10倍的條件,并且種間最小遺傳距離大于種內(nèi)最大遺傳距離。Barcoding gap檢驗(yàn)表明,穿山甲及其混偽品的COI序列具有一定的遺傳間隔。
從構(gòu)建的NJ樹(shù)可以看出,15個(gè)物種75份樣品均得到了較好的區(qū)分,各物種形成相對(duì)獨(dú)立的支。其中,產(chǎn)于亞洲的穿山甲物種和產(chǎn)于非洲的穿山甲物種形成兩個(gè)明顯的分支,說(shuō)明亞洲產(chǎn)穿山甲和非洲產(chǎn)穿山甲有明顯分化,親緣關(guān)系較遠(yuǎn),將其區(qū)分成不同的屬是合適的。亞洲支和非洲支又分別形成兩個(gè)分支,即中華穿山甲與印度穿山甲、馬來(lái)穿山甲與菲律賓穿山甲、大穿山甲與南非穿山甲、樹(shù)穿山甲與長(zhǎng)尾穿山甲。在新的分類系統(tǒng)中,穿山甲屬僅包括4種亞洲產(chǎn)穿山甲。其中中華穿山甲與印度穿山甲能爬樹(shù),但主要營(yíng)地棲生活,有觀點(diǎn)將兩者列為指名亞屬M(fèi)anis;馬來(lái)穿山甲和菲律賓穿山甲均傾向樹(shù)棲,后者以前一直作為前者的亞種,2005年才上升為獨(dú)立的種;兩者為另一亞屬Paramanis。大穿山甲和南非穿山甲形體大,地棲,幾乎不上樹(shù),在新的分類系統(tǒng)中歸屬于地穿山甲亞屬,或另立地穿山甲屬Smutsia。樹(shù)穿山甲和長(zhǎng)尾穿山甲則為樹(shù)棲,基本不下地,分別代表樹(shù)穿山甲亞屬Phataginus和長(zhǎng)尾穿山甲亞屬Uromanis,或組成樹(shù)穿山甲屬Phataginus[26]。
ABGD法是一種基于barcode gap對(duì)序列進(jìn)行分類的方法,它的優(yōu)點(diǎn)是快速、簡(jiǎn)單,即便出現(xiàn)種內(nèi)和種間遺傳距離分布重疊的現(xiàn)象,也可以對(duì)數(shù)據(jù)集進(jìn)行有效的劃分[23]。在本研究中將ABGD法作為對(duì)數(shù)據(jù)分類的一種補(bǔ)充方法,與聚類分析結(jié)合,可為分類結(jié)果的正確性提供支持,使得研究更加完善。用ABGD法進(jìn)行劃分,初始劃分只能對(duì)80%(12/15)的物種進(jìn)行解析。在遞歸劃分中,P值介于0.01~0.03時(shí),為較可靠的分組[22];本研究中,15個(gè)物種在P值為0.01~0.03時(shí),得到了穩(wěn)定的劃分,結(jié)果與形態(tài)鑒定結(jié)果、聚類分析結(jié)果呈對(duì)應(yīng)關(guān)系。
綜合以上結(jié)果可知,基于COI序列的DNA條形碼是穿山甲屬物種鑒定的有效手段,能夠?yàn)榇┥郊椎目焖勹b定提供可靠依據(jù)。
近年來(lái)已有穿山甲COI條形碼研究的報(bào)道[14-17]。本研究中,除增加了大穿山甲實(shí)體樣本外,更為重要的是建立的數(shù)據(jù)庫(kù)中包含了目前全球穿山甲屬物種所有的COI單倍型序列。另外,采取了序列相似性分析、系統(tǒng)發(fā)育樹(shù)構(gòu)建和ABGD遞歸劃分3種方法進(jìn)行綜合鑒定。全面的數(shù)據(jù)庫(kù)和綜合鑒定手段,提高了穿山甲的鑒定效率和準(zhǔn)確性。
應(yīng)用DNA條形碼的方法在穿山甲商品的鑒定中發(fā)現(xiàn)正品穿山甲甚少,主流商品為馬來(lái)穿山甲和印度穿山甲。印度穿山甲以前較少報(bào)道,目前市場(chǎng)上逐漸增多。我們對(duì)印度穿山甲的性狀特征進(jìn)行了詳細(xì)描述,發(fā)現(xiàn)其與中華穿山甲、馬來(lái)穿山甲在形態(tài)上極為相似(樹(shù)穿山甲和大穿山甲有比較明確的性狀鑒別特征[6])。我們又對(duì)3種穿山甲甲片密度進(jìn)行了測(cè)定,發(fā)現(xiàn)馬來(lái)穿山甲、印度穿山甲、中華穿山甲的平均密度分別為1.49、1.57、1.60 g·cm-3;馬來(lái)穿山甲質(zhì)地較輕,而印度穿山甲與中華穿山甲的質(zhì)地接近。雖然三者從性狀上難以區(qū)分,但用DNA條形碼可準(zhǔn)確鑒定其基原。
基于COI序列的DNA條形碼鑒別方法,可從分子水平實(shí)現(xiàn)對(duì)穿山甲的有效鑒定,為穿山甲的市場(chǎng)監(jiān)管、打擊非法貿(mào)易及保護(hù)穿山甲資源提供技術(shù)支持,具有重要的實(shí)用價(jià)值。目前,這種方法在市售鹿茸粉、僵蠶、海馬等藥材中也取得了成功,說(shuō)明DNA條形碼技術(shù)在動(dòng)物藥商品鑒定中是行之有效的,值得推廣應(yīng)用[27-29]。
猜你喜歡
課堂內(nèi)外·初中版(科學(xué)少年)(2025年1期)2025-02-28 00:00:00
課堂內(nèi)外·初中版(科學(xué)少年)(2025年2期)2025-02-28 00:00:00
課堂內(nèi)外·初中版(科學(xué)少年)(2024年12期)2024-12-02 00:00:00
英語(yǔ)世界(2023年10期)2023-11-17 09:18:18
中學(xué)生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(shuō)(2021年8期)2021-11-03 10:55:16
學(xué)苑創(chuàng)造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(shù)(快樂(lè)歷史地理)(2020年9期)2020-03-19 05:10:56
科學(xué)大眾(中學(xué))(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
