間歇投喂及越冬對草魚幼魚肝臟抗氧化能力的影響
2019-09-18 03:15:22
長江大學學報(自科版) 2019年8期
關鍵詞:影響
(安徽農業大學動物科技學院,安徽 合肥 230036)
沐璟瑋
(安徽師范大學生命科學學院,安徽 蕪湖 241000)
李鵬程,姚峰,岳鼎鼎
(安徽農業大學動物科技學院,安徽 合肥 230036)
陳路
(農業農村部華南水產與畜禽飼料重點實驗室,廣東恒興飼料實業股份有限公司,廣東 湛江 524094)
楊嚴鷗
(安徽農業大學動物科技學院,安徽 合肥 230036)
在正常的生理代謝過程中,魚類的細胞內會產生少量活性氧自由基(reactive oxygen species,ROS),但當遇到各種脅迫時,活性氧自由基則會過量產生[1,2],這些自由基對生物機體產生一系列損傷,例如引起細胞膜發生脂質過氧化、改變生物膜結構和功能[1~3]等。為了抵抗這種損傷,生物體內進化出了有效的抗氧化系統[4,5],而營養不足會影響魚類的抗氧化指標。謝雨欣等[6]研究了饑餓和再投喂對黃顙魚幼魚抗氧化能力的影響;Dong等[7]研究了飼料蛋白質不足對黃顙魚幼魚抗氧化能力的影響,溫度變化也會影響魚類的抗氧化指標;郭勤單等[4]研究了溫度應激對褐牙鲆幼魚抗氧化能力的影響;彭婷等[5]研究了低溫脅迫對尼羅羅非魚免疫及抗氧化指標的影響;馮廣朋等[8]研究了溫度對中華鱘幼魚抗氧化酶活性的影響。但是魚類在生長過程中往往經歷不止一種環境脅迫,例如一些魚類經歷了一段時間的攝食不足后就進入越冬期,在這種情況下,其抗氧化指標如何變化還不太清楚,因此有必要進行研究,以更多地了解脅迫條件下魚類抗氧化能力的變化。為此,筆者以我國重要的經濟養殖魚類草魚(Ctenopharynodonidellus)為試驗魚對這一問題進行了探討。
1 材料與方法
1.1 試驗材料
試驗飼養設備為循環水養魚系統。水源為曝氣自來水,中央凈水系統主要使用沸石、活性炭等凈化水質。水族箱單個規格為60cm×60cm×60cm(長×寬×高),盛水180L,單箱循環水量1.00L/min;羅茨鼓風機接通風管再接散氣石對水箱充氧,每隔5min充氣2min。試驗期間水體總氨氮質量濃度低于(0.12±0.02)mg/L,溶氧量大于6.2mg/L,pH 7.2±0.1,電熱棒與空調控制水溫(25±1)℃,日光燈8:00~20:00照明。室內配備2臺抽濕機抽出水氣,維持空氣干燥。
試驗魚購自安徽合肥市漁場,為當年魚種,實驗室暫養7d以上。暫養期間所喂飼料與試驗飼料為同一種配合飼料,配方參見文獻[9],配制時將原料磨成粉狀物,混勻后經飼料機制成粒徑1.5mm顆粒,65℃烘干后于-20℃保存。
1.2 試驗方法
養殖試驗于10月至12月進行。試驗設4組,每組4個重復水族箱,共16個水族箱,先將魚饑餓24h再隨機取樣稱重分箱,每箱30尾魚。第1組每天投喂,第2、3、4組分別喂6、5、4d后再分別饑餓1、2、3d;4個組分別以S0(對照組)、S1/6、S2/5和S3/4表示。每天上、下午各投過量飼料1次,1.5h后回收殘餌。
養殖8周后,將魚饑餓24h,稱重每箱魚體重,取7尾魚置于冰盤取肝胰臟,等量混合后-80℃保存以測定抗氧化指標。水溫逐漸下降到室內自然水溫,經56d越冬(室內自然水溫4~9℃)后用上述方法取肝胰臟,保存以測定抗氧化指標。超氧化物歧化酶(SOD)活性、過氧化氫酶(CAT)活性、維生素E(VE)含量和丙二醛(MDA)含量采用南京建成生物工程研究所的試劑盒進行測定,具體測定方法按照試劑盒的說明進行。
1.3 數據分析
描述性統計值用平均值±標準差表示,單因素方差分析(ANOVA)后進行組間差異的多重比較(Duncan’s procedure)。顯著性水平設置為α=0.05,統計軟件為SPSS 16.0。
2 結果與分析
2.1 不同投喂模式對草魚終末濕重及干重的影響
試驗中無魚死亡。由表1可知,S1/6和S2/5的終末濕重和干重均與對照組無顯著差異(P>0.05),S3/4則顯著低于對照組(P<0.05)。

表1 不同投喂模式對草魚終末濕重及干重的影響(平均值±標準差)*
注:同列數據上標字母不同表示有顯著差異(P<0.05)。
2.2 不同投喂模式對草魚肝臟抗氧化指標的影響
由表2可知,在越冬前與越冬后,不同饑餓處理對SOD活性都沒有顯著影響,但越冬前有先下降后再上升的趨勢(P>0.05);CAT活性越冬前均顯著高于越冬后,隨著饑餓時間增加在不同時間段的變化趨勢也不同,越冬前饑餓時間越長則CAT活性越低(P<0.05),越冬后則CAT活性則顯著上升(P<0.05);MDA與VE含量變化趨勢一致,越冬前兩者的含量隨著饑餓時間延長顯著上升后再下降,越冬后則是顯著下降再上升(P<0.05);谷胱甘肽過氧化物酶(GPX)活性在越冬前隨著饑餓時間增加均顯著上升后再下降,越冬后則是顯著上升(P<0.05)。

表2 不同投喂模式對草魚肝臟抗氧化指標的影響
注:同行數據上標小寫字母不同表示不同處理組之間有顯著差異(P<0.05);大寫字母不同表示同一處理組越冬前后有顯著差異。
3 討論
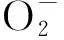
VE是重要的抗氧化劑,草魚飼料中添加VE能促進生長,增強機體抗氧化能力[15],機體內VE蓄積量的增加可抑制不飽和脂肪酸過氧化,使過氧化產物丙二醛含量降低[16]。該研究中,越冬前S1/6和S2/5組的VE含量上升,這可能表明一定的饑餓脅迫能激發VE含量,在CAT酶活性降低時能起到一定的抗氧化作用。
MDA是不飽和脂肪酸過氧化終產物之一,能與蛋白質的游離基作用,導致細胞損傷,其含量的高低能反映機體細胞受自由基攻擊的嚴重程度,因此被當作是一種細胞膜氧化損傷的指示物[17]。本研究中,間歇性饑餓處理導致越冬前3個試驗組的MDA含量均顯著高于對照組,這說明魚體處于一定的氧化脅迫狀態,對瓦氏黃顙魚[11]研究也表明,饑餓導致瓦氏黃顙魚肝臟的MDA含量顯著上升。越冬后,對照組、S1/6與S2/5組的MDA含量均顯著下降,這可能與魚體整體的代謝水平下降有關,但S3/4組的MDA顯著增加,原因可能是S3/4組的投喂方式對魚體抗氧化機能損傷過大,越冬導致這種后果表現出來,抗氧化系統不能更為有效地清除魚體產生的自由基,導致MDA含量上升。
GPX以谷胱甘肽為底物清除H2O2,是一類具有明確酶活性的硒蛋白,它通過催化谷胱甘肽(Glutathione,GSH)為氧化型谷胱甘肽(oxidized glutathione,GSSG),使有毒的過氧化氫及脂質過氧化物還原為無毒的羥基化合物,從而保護細胞膜結構及功能不受過氧化物損傷及干擾[18,19]。該研究中3個試驗組的GPX活性在越冬前后基本都顯著高于對照組,應該是環境脅迫的激發所致[19]。
該研究結果顯示,養殖投喂階段結束時,S1/6組和S2/5組的濕重與干重都與對照組無顯著差異,而饑餓時間最長S3/4組體重未能趕上對照組,這表明適當的間歇饑餓不會影響草魚的體重生長,但無論何種間歇饑餓都會導致草魚肝臟的抗氧化能力下降,經歷越冬后,與連續投喂的對照組相比,3個間歇投喂組仍然保持抗氧化應激狀態。
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
知識經濟·中國直銷(2016年3期)2016-02-27 16:15:49
現代檢驗醫學雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學低年級版)(2011年3期)2011-01-01 00:00:00
