毛果蕓香堿對不同光照下斑馬魚幼魚自發運動和焦慮性行為的影響
2019-10-09 01:01:48劉秀云張穎藍李婷婷
復旦學報(醫學版) 2019年5期
簡 靜 林 佳 劉秀云 張穎藍 李 斐 王 鳳 李婷婷 郭 寧 李 強
(1復旦大學附屬兒科醫院兒科研究所-復旦大學兒童發育與疾病轉化醫學研究中心-上海市出生缺陷防治重點實驗室 上海 201102; 2上海中醫藥大學中醫方證與系統生物學研究中心 上海 201203)
斑馬魚是科學研究廣泛使用的一種重要脊椎動物模型[1],目前已被廣泛用于遺傳、毒理[2-3]和發育生物[4]等學科的研究。Rihel等[5]證明斑馬魚可以作為神經系統相關藥物研究的一個快速、可靠且成本較低的工具, 在神經疾病的病因學方面與嚙齒動物和人類有許多共同性[6]。4~5天的斑馬魚幼魚即可表現出狩獵,回避和趨觸等廣泛的行為學[7-9]變化,在人類認識疾病的發病機制和新藥的研發過程中發揮著重要的作用[10-11]。斑馬魚的凍結、旋轉[12]、漫淡、聚集[13]、沉底、回避和趨觸[14]等行為都可以反映斑馬魚幼魚的神經性行為,且趨觸性已經被作為衡量斑馬魚幼魚壓力/焦慮水平的最重要指標,可直接在多孔板中進行。
趨觸性行為是指動物在所處區域邊界附近停留或移動的偏好。將多孔板中心占整個孔面積50%的圓形區域定義為斑馬魚的中心活動區域。用兩個相互關聯的參數來研究斑馬魚幼魚的趨觸性行為:第一個為斑馬魚幼魚在中心區域移動距離的百分比(后文公式1),第二個為斑馬魚幼魚在中心區域花費時間的百分比(后文公式2)。
毛果蕓香堿是常用的膽堿能激動劑,研究人員常通過毛果蕓香堿誘導大鼠的慢性癲癇持續狀態對慢性癲癇生理學和治療手段進行探索[15]。 有研究者用毛果蕓香堿誘導的小鼠癲癇模型具有顯著的焦慮樣狀態[16-17],但對用抗癲癇藥物進行干預時卻只發現其抑郁狀態顯著降低而焦慮未被抑制[17-18],Liu和 Duarte等[19-20]的研究結果表明毛果蕓香堿對大鼠具有致焦慮的特性。以上結果表明毛果蕓香堿具有誘發嚙齒類動物產生焦慮的作用,但很少有研究者在斑馬魚上進行毛果蕓香堿的研究。探索毛果蕓香堿對斑馬魚幼魚自發運動和焦慮相關因素的影響可以為斑馬魚焦慮模型的建造和焦慮癥狀的治療提供新思路新方向。
光照-黑暗交替的視覺反應測試可以引起斑馬魚幼魚的超運動行為,在光照-黑暗轉變期間斑馬魚幼魚趨觸性行為增加[21]。 因此,我們研究了毛果蕓香堿在連續光照和光照-黑暗交替條件下對5天齡(5dpf)和7天齡(7dpf) AB型斑馬魚幼魚自發運動和趨觸性行為的影響,并發現毛果蕓香堿誘導AB型斑馬魚幼魚產生的焦慮反應呈現出藥物濃度和斑馬魚幼魚年齡的依賴性。
材料和方法
實驗動物野生AB型斑馬魚,復旦大學附屬兒科醫院兒研所斑馬魚平臺。 根據Westerfield《斑馬魚胚胎發育指南》[22],將斑馬魚飼養在(28.5±0.5) ℃的條件下。斑馬魚自然交配后收集胚胎至直徑90 mm的培養皿中,每培養皿養殖50顆左右魚卵。從出生到7dpf的魚卵培養皿均放置在(28.5±0.5) ℃的培養箱中培養,期間不予喂食。每天給予14 h光照和10 h黑暗周期(08∶00時開燈,22∶00關燈)。所有動物實驗測試程序均符合國際規定,所有協議由復旦大學附屬兒科醫院動物管理委員會授權。
藥物將毛果蕓香堿(P6503-5G,Sigma-Aldrich)用無菌水溶解到10 mmol/L儲備溶液中,并以1 mL每管冷凍于-20 ℃冰箱。實驗之前,將毛果蕓香堿儲備液用斑馬魚系統水稀釋到所需濃度。
儀器和耗材斑馬魚養殖系統-北京愛生公司,Leica205C顯微鏡-德國Leica公司,ZebraLab分析系統-法國Viewpoint公司;細胞培養24孔TCT板-上海吉泰遠成公司。
曠場實驗所有行為測試都在環境溫度為28.5 ℃的房間內進行,斑馬魚幼魚魚卵隨機分配在24孔板中進行行為學測試。所有實驗均在培養箱中光照后2 h和黑暗前2 h之間進行。每個24孔板中魚卵均勻的呈現所有濃度,以避免實驗次序導致的任何差異。
行為學檢測開放曠場和光照設計如圖1所示:在實驗前將斑馬魚幼魚轉移至24孔板中,每孔中一條幼魚,孔的內徑為15.88 mm,幼魚可自由活動(圖1A)。去除孔中多余的液體后立即加入1 mL的毛果蕓香堿工作溶液,毛果蕓香堿最終的測試濃度為0、0.1、0.3、1、3、10 mmol/L。然后將24孔板放入帶有記錄儀(ViewPoint Life Sciences)的ZebraBox中記錄斑馬魚幼魚的活動視頻。實驗設置為20 min光照適應,10 min持續光照研究斑馬魚幼魚的自發運動和趨觸性,然后分別是5 min黑暗刺激和5min光照恢復(圖1B),以檢測在光照變化時毛果蕓香堿對斑馬魚幼魚自發運動和趨觸性的影響。因此,包括適應期在內,每次實驗持續40 min。圖2顯示的是不同濃度的毛果蕓香堿在整個實驗過程中對斑馬魚幼魚每分鐘自發游動距離的影響。
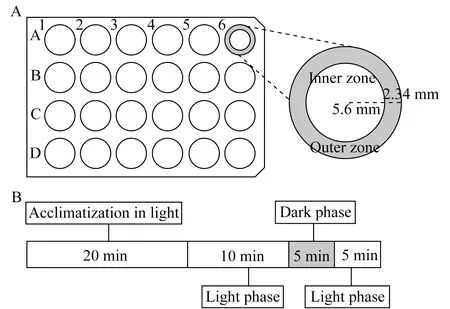
A:The testing apparatuses consist of each well in a 24-well plate format (diameter of 15.88 mm).The inner and outer zones were delineated as shown above.The width of the outer zone was set at 2.34 mm relative to the border of the well;B:The procedure including acclimatization and experiment procedure,acclimatization 20 mins,and experiment procedure light phase (0-10 min),sudden illumination transition from light to dark phase (10-15 min) and dark to light phase (15-20 min).
圖1 開放曠場和光照設計示意圖
Fig 1 The open field test and light testing apparatuses
參數計算使用ZebraLab軟件的跟蹤模式來記錄視頻,完成斑馬魚幼魚運動行為的定量。斑馬魚幼魚的視頻以25 fps的速率拍攝,可檢測幼魚任意速度級別的運動。在每個孔中,確定了一個占據單井面積一半的圓形中心區域。幼魚在整個孔中移動的距離可以用來分析斑馬魚的自發運動。對趨向性行為的分析來自于[23]對移動距離和幼魚在中心舞臺上花費的時間的修改。 公式如下所示
(公式1)
(公式2)
結 果
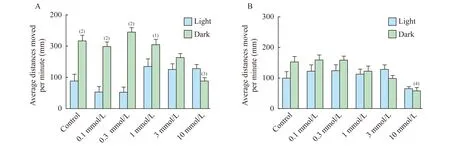
連續光照條件下毛果蕓香堿對不同年齡階段斑馬魚自發運動的影響毛果蕓香堿對5dpf斑馬魚自發運動的影響以藥物濃度0.3 mmol/L分為兩個階段。0.1 mmol/L的毛果蕓香堿對斑馬魚幼魚的自發運動有明顯的抑制作用(P=0.021 3);而當藥物濃度大于0.3 mmol/L后,隨著藥物濃度的增加反倒對斑馬魚的自發運動有一定的刺激作用但均沒有統計學意義(圖3A)。
毛果蕓香堿對7dpf斑馬魚自發運動的影響不同于對5dpf斑馬魚,只有10 mmol/L毛果蕓香堿使斑馬魚幼魚的游動距離減弱,其他各組均表現出一定的刺激作用,但所有濃度組均沒有統計學意義(圖3B)。
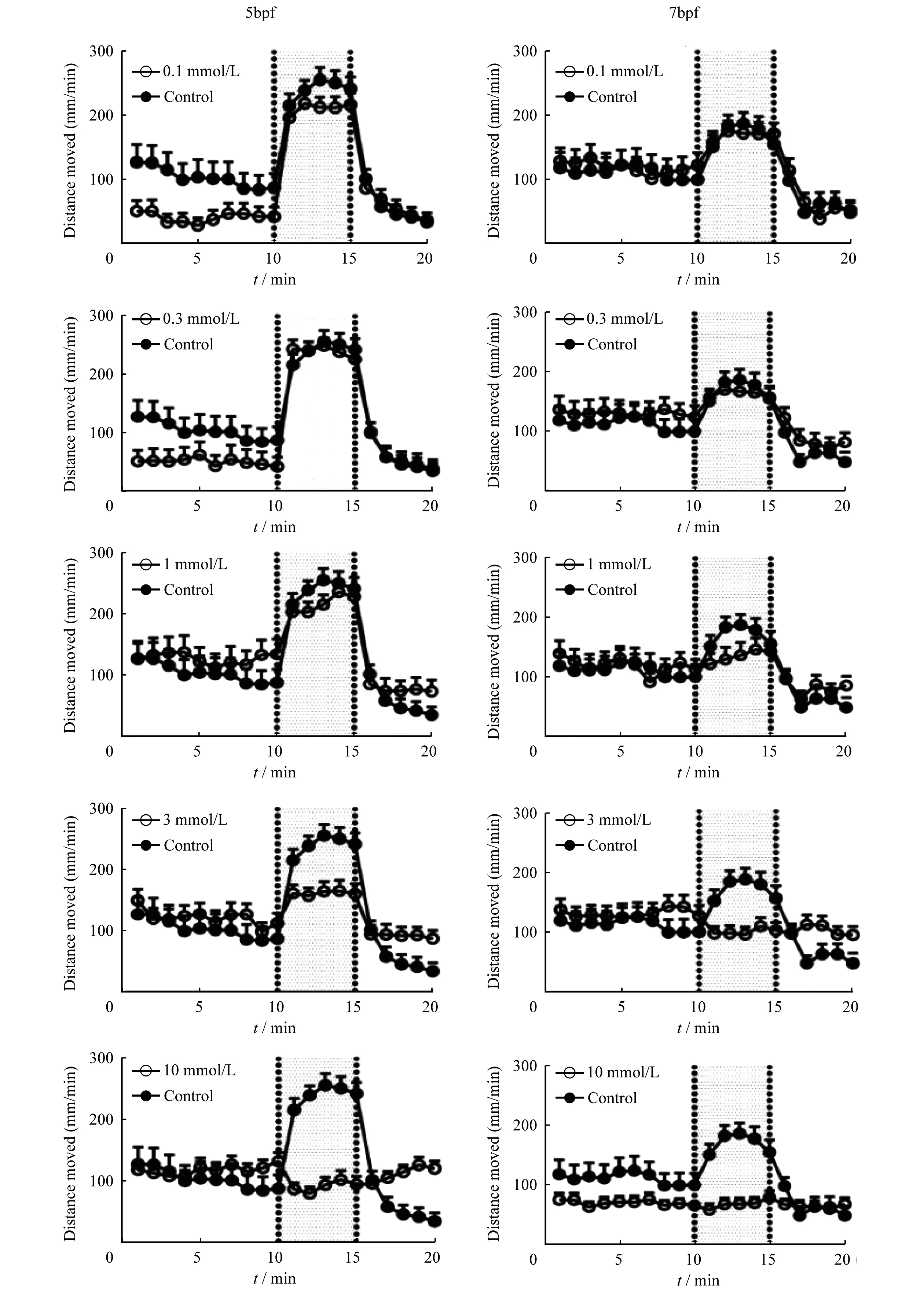
The distance moved in each 1-min time bin during 20-min measurements period (0-min to 20-min) was plotted against the progression of the experiment.The control group (open circles) was plotted with a single pilocarpine concentration group (filled circles) in each panel.The shaded part in each panel represents the 5-min dark phase,and the non-shaded part in each panel represents the light phase.
圖2 不同濃度的毛果蕓香堿在整個實驗過程中對斑馬魚幼魚每分鐘自發游動距離的影響
Fig 2 Distance moved by zebrafish larvae within the entire zone during all experiment measurements periodunder different concentrations of pilocarpine
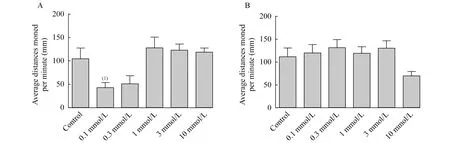
A:5dpf;B:7dpf.(1)vs.Control,P<0.05.
圖3 連續光照條件下不同濃度毛果蕓香堿對5dpf和7dpf斑馬魚自發運動的影響
Fig 3 Effects of pilocarpine exposure on the locomotor activity of zebrafish larvae at 5dpf and 7dpf under continuous light condition
連續光照條件下毛果蕓香堿對不同年齡階段斑馬魚趨觸性行為的影響在沒有使用毛果蕓香堿處理的對照組5dpf和7dpf的斑馬魚幼魚都表現出一定的趨觸性行為,而使用較高濃度的毛果蕓香堿后5dpf和7dpf的斑馬魚幼魚趨觸性行為都有所增加。 5dpf的斑馬魚幼魚的趨觸性行為并未因為毛果蕓香堿濃度的改變而產生統計學差異,但在濃度大于1 mmol/L后其中心區域游動距離(圖4A)和時間(圖4C)均隨藥物濃度增高而降低,呈現出藥物依賴性的趨觸性增加。相比之下,當藥物濃度大于0.1 mmol/L后7dpf斑馬魚趨觸性行為的改變呈現出一定的藥物濃度依賴性,尤其3 mmol/L和10 mmol/L的毛果蕓香堿使7dpf斑馬魚趨觸性行為明顯增加,即中心區域游動距離減少(P=0.005 5,P=0.000 8)(圖4B)以及中心區域游動時間減少(P=0.001 4,P=0.001 8)(圖4D)。
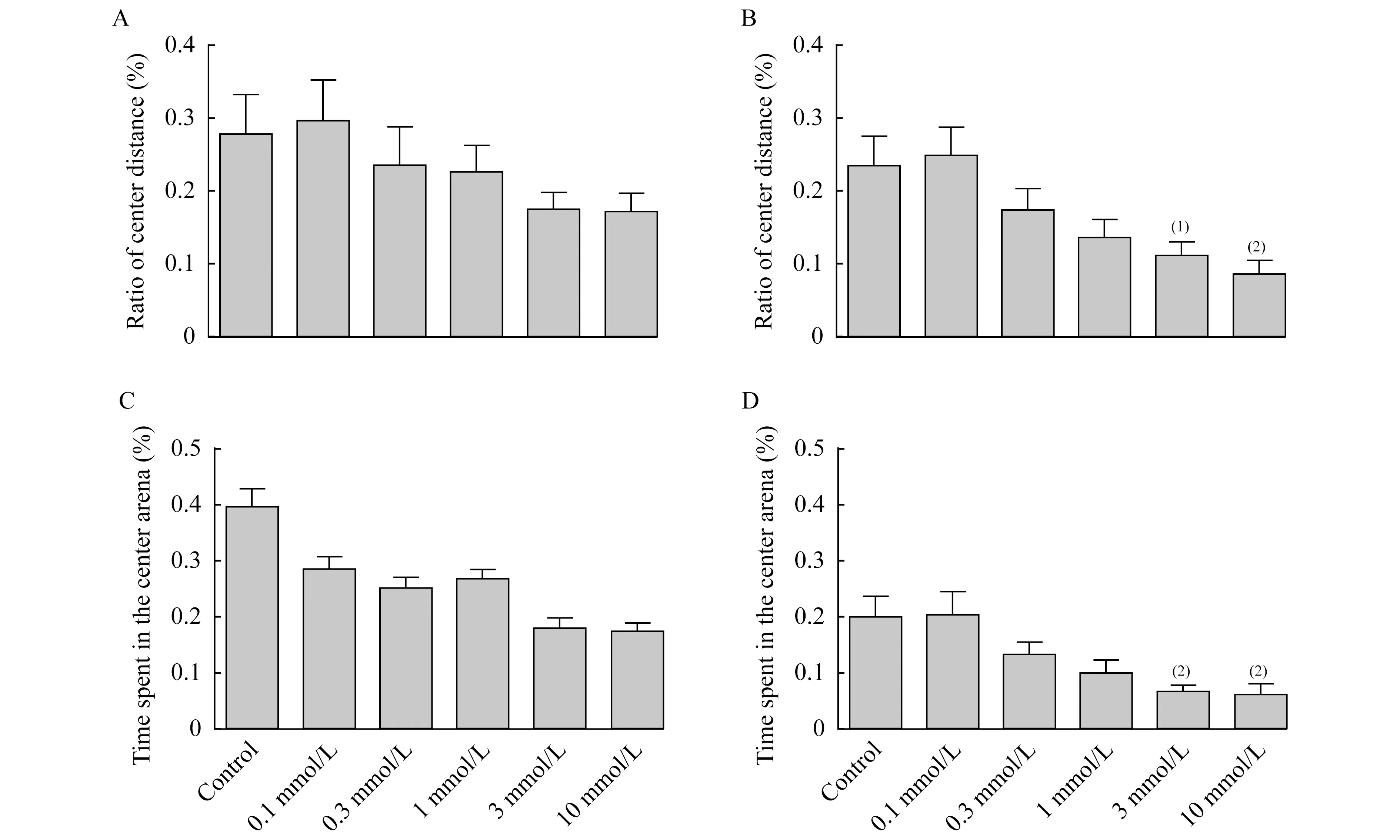
A,C:5dpf;B:7dpf.vs.Control,(1)P<0.05,(2)P<0.01.
圖4 連續光照條件下不同濃度毛果蕓香堿對5dpf和7dpf斑馬魚趨觸性的影響
Fig 4 Effects of pilocarpine exposure on the thigmotaxis of zebrafish larvae at 5dpf and 7dpf under continuous light condition
光暗交替階段毛果蕓香堿對斑馬魚自發運動和趨觸性行為的影響
毛果蕓香堿對5dpf斑馬魚光照變化刺激的影響 雙因素方差分析顯示:F光照(1,372)=80.87,P光照<0.000 1;F毛果蕓香堿 (5,372)=3.297,P毛果蕓香堿=0.006 3。光照變化前的1 min內,0.1、0.3 mmol/L毛果蕓香堿降低了斑馬魚幼魚的自發運動,而1、3、10 mmol/L均增加了斑馬魚幼魚的自發運動,但未產生統計學差異。黑暗階段,10 mmol/L毛果蕓香堿顯著抑制了斑馬魚的自發運動(P<0.000 1)。在相同濃度的毛果蕓香堿時,對照0.1、0.3、1 mmol/L毛果蕓香堿處理的斑馬魚幼魚在黑暗環境中較光照條件下自發運動明顯增強(P<0.000 1,P<0.000 1,P<0.000 1,P=0.024 2),而10 mmol/L時黑暗環境下毛果蕓香堿對斑馬魚自發運動的抑制已經完全抵消掉了黑暗本身對斑馬魚視覺刺激下的自發運動升高現象(圖5A)。
毛果蕓香堿對7dpf斑馬魚光照變化刺激的影響 雙因素方差分析顯示:F光照(1,372)=2.6,P光照=0.107 7;F毛果蕓香堿(5,372)= 6.431,P毛果蕓香堿<0.000 1,與毛果蕓香堿對5dpf斑馬魚光照變化刺激的影響明顯不同,7dpf的斑馬魚幼魚只在黑暗刺激下且用10 mmol/L的毛果蕓香堿處理時其自發運動才與對照組有明顯的差異(P=0.000 3),而其他各濃度處理后光暗變化未對斑馬魚幼魚的自發運動產生統計學的改變。3、10 mmol/L的毛果蕓香堿在黑暗環境下對斑馬魚自發運動的抑制已經完全抵消掉了黑暗本身對斑馬魚視覺刺激下的自發運動升高現象(圖5B)。
A:5dpf;B:7dpf.There were signicant differences between the light and dark conditions within the same pilocarpine concentration group,(1)P<0.05,(2)P<0.000 1.Versus the control group under the same lighting conditions (light or dark),(3)P<0.000 1,(4)P<0.001.
圖5 光暗交替條件下不同濃度毛果蕓香堿對5dpf和7dpf斑馬魚自發運動的影響
Fig 5 Effects of pilocarpine exposure on the locomotor activity of zebrafish larvae at 5dpf and 7dpf under light and dark condition
毛果蕓香堿對5dpf斑馬魚幼魚趨觸性行為的影響雙因素方差分析斑馬魚幼魚中心區域游動距離F光照(1,336)=3.398,P光照=0.005 2;F毛果蕓香堿(5,336)=91.7,P毛果蕓香堿<0.000 1。各濃度組在光照變化前的1 min內對斑馬魚幼魚趨觸性行為的影響差異無統計學意義,在黑暗刺激下3mmol/L的毛果蕓香堿顯著的增加了5dpf斑馬魚的趨觸性行為(P=0.000 1)。但光暗變化期間,所有濃度的毛果蕓香堿處理都使得5dpf的斑馬魚幼魚在黑暗環境下較光照環境下的趨觸性行為降低(P=0.000 7,P=0.013 9,P=0.038 7,P<0.000 1,P=0.000 3,P<0.000 1)(圖6A)。
雙因素方差分析斑馬魚幼魚中心區域游動時間F光照(1,341)=88.18,P光照<0.000 1;F毛果蕓香堿(5,341)=5.55,P毛果蕓香堿<0.000 1,光照條件下,3 mmol/L和10 mmol/L的毛果蕓香堿明顯的增加了斑馬魚幼魚的趨觸性行為(P=0.003 4,P=0.034 0)。但各濃度毛果蕓香堿處理后5dpf的斑馬魚幼魚在光暗交替階段黑暗環境下的趨觸性行為降低,除0.1 mmol/L之外,對照0.3、1、3、10 mmol/L的毛果蕓香堿使黑暗環境下5dpf斑馬魚趨觸性行為降低較其在光照條件下有明顯的統計學差異(P=0.002 1,P=0.014 0,P=0.000 7,P<0.000 1,P<0.000 1)(圖6C)。
毛果蕓香堿對7dpf斑馬魚幼魚趨觸性行為的影響用雙因素方差分析斑馬魚幼魚在中心區域的游動距離:F光照(1,361)=6.783,P光照<0.000 1;F毛果蕓香堿(5,361)=58.42,P毛果蕓香堿<0.000 1。光照條件下,斑馬魚幼魚的趨觸性行為隨毛果蕓香堿濃度增加而逐漸增加,當毛果蕓香堿濃度達到3 mmol/L和10 mmol/L時明顯的增加了斑馬魚幼魚的趨觸性行為(P=0.032 2,P=0.008 6);在黑暗刺激下,毛果蕓香堿濃度>1 mmol/L時隨毛果蕓香堿濃度增加而逐漸增加,10 mmol/L的毛果蕓香堿明顯的增加了7dpf斑馬魚的趨觸性行為(P=0.000 1)。所有濃度的毛果蕓香堿處理都使得7dpf的斑馬魚幼魚在黑暗環境下趨觸性行為較光照條件下降低,且0.1、0.3、1、3 mmol/L的毛果蕓香堿在黑暗環境下對斑馬魚趨觸性行為的抑制作用較光照條件下差異有統計學意義(P<0.000 1,P=0.000 7,P=0.010 3,P=0.005 4,圖6B)。
雙因素方差分析斑馬魚幼魚中心區域游動時間:F光照(1,364)=5.313,P光照<0.000 1;F毛果蕓香堿(5,364)=57.21,P毛果蕓香堿<0.000 1。 光照條件下,3 mmol/L和10 mmol/L的毛果蕓香堿明顯的增加了斑馬魚幼魚的趨觸性行為(P=0.038 2,P=0.034 9);在黑暗刺激下10 mmol/L的毛果蕓香堿增加了7dpf斑馬魚的趨觸性行為(P=0.001 1)。所有濃度的毛果蕓香堿處理都使得7dpf的斑馬魚幼魚在黑暗環境下趨觸性行為較光照降低,且對照0.1、0.3、1、3 mmol/L的毛果蕓香堿在黑暗環境下對斑馬魚趨觸性行為的抑制作用較光照條件下差異有統計學意義(P=0.033 2,P=0.000 3,P=0.003 2,P=0.023 1,P=0.006 8,圖6D)。
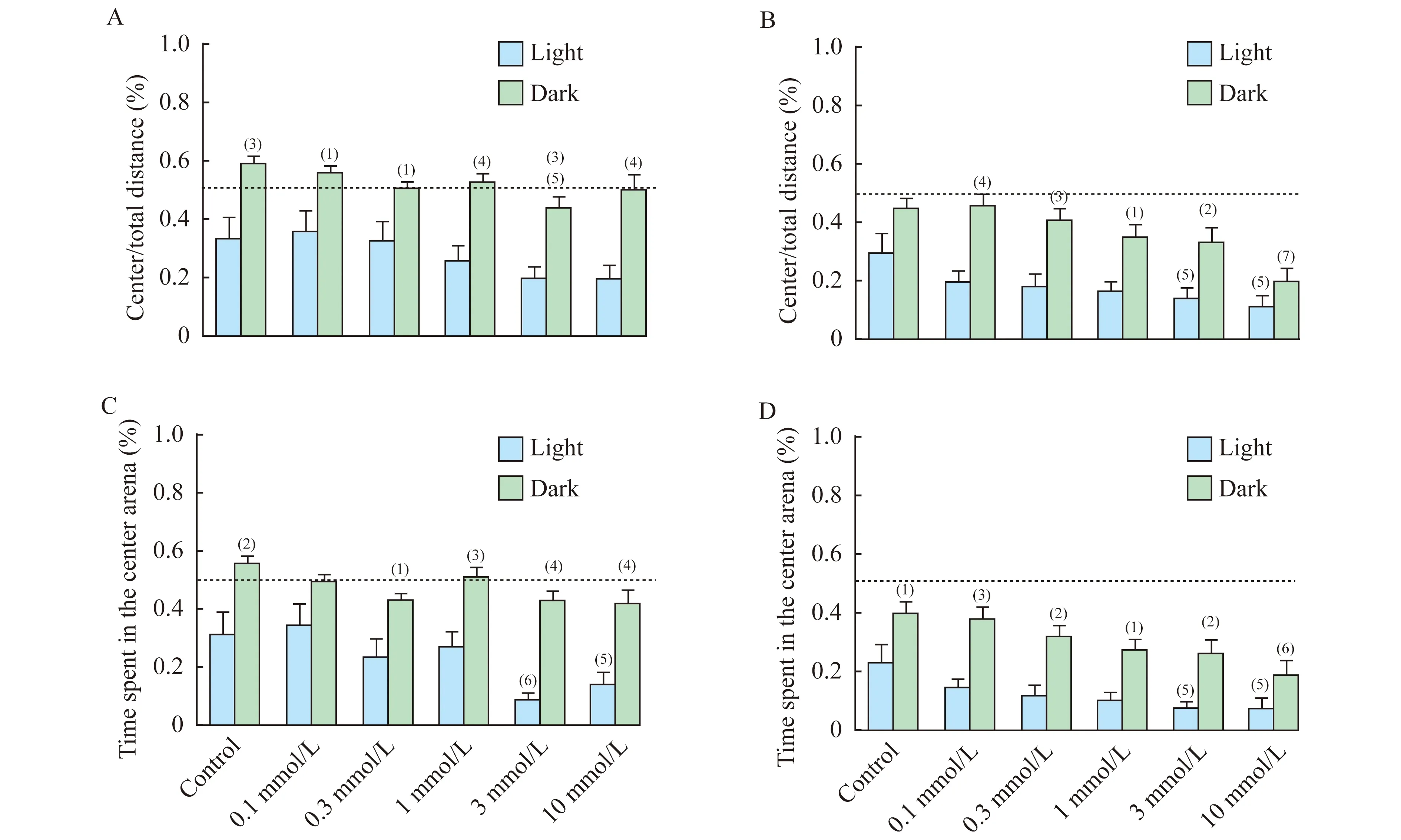
A,C:5dpf;B,D:7dpf.There were signicant differences between the light and dark conditions within the same pilocarpine concentration group,(1)P<0.05,(2)P<0.01,(3)P<0.001,(4)P<0.000 1.Versus the control group under the same lighting conditions (light or dark),(5)P<0.05,(6)P<0.01,(7)P<0.001.
圖6 光暗交替條件下不同濃度毛果蕓香堿對5dpf和7dpf斑馬魚趨觸性的影響
Fig 6 Effects of pilocarpine exposure on the thigmotaxis of zebrafish larvae at 5dpf and 7dpf under light and dark condition
討 論
毛果蕓香堿對不同年齡斑馬魚幼魚自發運動的影響不同本研究分別考察了毛果蕓香堿處理對連續光照和光暗交替條件下斑馬魚幼魚自發運動的影響,5dpf和7dpf的斑馬魚在毛果蕓香堿處理后沒有表現出相同的趨勢。在5dpf,斑馬魚幼魚的自發運動隨著毛果蕓香堿濃度的增加先降低再升高,而7dpf斑馬魚的自發運動則呈持續性的增加,直到毛果蕓香堿濃度達最高濃度10 mmol/L時斑馬魚幼魚的活動受到抑制。5dpf的斑馬魚幼魚與對照組相比,0.1 mmol/L時其自發運動受到最大的抑制,但在10 mmol/L藥物濃度時毛果蕓香堿并未對斑馬魚幼魚的自發運動造成明顯的改變,這可能與毛果蕓香堿毒蕈堿樣中毒癥狀造成的肌肉痙攣收縮相關。我們的研究中毛果蕓香堿對斑馬魚幼魚自發運動的影響與以往在環鴿研究中的結果相似[8]。在對環鴿的研究中,急性施用毒蕈堿激動劑毛果蕓香堿 (1和3 mg/kg i.p.) 后,處理組在第1 h和第2 h期間引起了環鴿自發運動劑量依賴性的降低。但是與Shannon等[25]在小鼠上的研究結果不一致,他們發現用毛果蕓香堿干預不會造成小鼠自發運動的改變,這可能是因為在不同的研究中,實驗動物的物種、年齡以及實驗干預的方式尤其時間等都不完全相同。普遍認為膽堿能神經元參與攻擊,運動及認知等高級功能的調節[26];本研究中持續光照條件下較高濃度組毛果蕓香堿處理的斑馬魚幼魚自發運動增強可能與毛果蕓香堿的膽堿能激活作用相關。
毛果蕓香堿可在一定程度上增加斑馬魚幼魚的焦慮水平Richendrfer等[21]將5-7dpf斑馬魚放入多孔板后,幼魚表現出對孔邊緣區域的強烈偏好,且這種偏好可以通過環境刺激進一步增強,反映出這種外來的環境刺激增加了斑馬魚幼魚的焦慮水平。
綜合中心區域的距離和時間兩種指標發現,除了0.1 mmol/L組外,所有濃度毛果蕓香堿處理均導致斑馬魚幼魚趨觸性隨濃度和年齡的增加而增加,反映出焦慮水平的增加。相似地Liu等[19]在試驗中用低于驚厥劑量的毛果蕓香堿對大鼠進行腹腔注射,結果表明次于驚厥劑量的毛果蕓香堿對大鼠具有致焦慮特性。我們研究中觀察到毛果蕓香堿的致焦慮作用與其他學者在嚙齒動物模型中的結果一致,且近期已有研究進一步對毛果蕓香堿誘導焦慮的機制進行了探索。普遍認為毛果蕓香堿導致的焦慮可在癲癇的診斷之前或癲癇后期合并癥中存在,主要與大腦皮質、海馬及眾多神經節和傳導通路內γ-氨基丁酸能神經元受到抑制相關,GABA的缺乏或抑制會導致機體的焦慮、不安;進一步實驗表明這種焦慮的產生可以被外源性膠質細胞源性神經營養因子挽救[27]。
我們首次研究了毛果蕓香堿處理對斑馬魚幼魚自發運動的影響,并觀察到了毛果蕓香堿對斑馬魚幼魚的致焦慮特征,且7dpf的斑馬魚幼魚趨觸性行為的兩個衡量參數較5dpf斑馬魚更一致,這可能與7dpf斑馬魚的游動能力以及自發運動行為更為完善相關。
低濃度毛果蕓香堿可引起斑馬魚幼魚的光暗應激反應,而毛果蕓香堿降低了暗刺激對斑馬魚幼魚自發運動的刺激作用4dpf時,野生型斑馬魚幼魚便可以對光暗條件的改變做出反應,在光線變暗時表現出自發運動的增加[28-29]。在本研究中,低濃度毛果蕓香堿處理組斑馬魚幼魚在整個研究過程中對光暗變化刺激始終有反應;但當毛果蕓香堿達到10 mmol/L時,黑暗刺激下斑馬魚幼魚超運動已經被完全廢除。在5dp和7dpf對照組以及0.1、0.3、1 mmol/L毛果蕓香堿處理的5dpf斑馬魚組,黑暗刺激產生的超級運動具有統計學意義,而7dpf斑馬魚在任何濃度的毛果蕓香堿處理下其超運動的產生均不具有統計學意義。也就是意味著,較高濃度的毛果云香堿及較完整的斑馬魚神經發育可以削弱光暗轉變后暗刺激所激發的斑馬魚超運動行為,具體機制有待后續實驗進一步探索。
毛果蕓香堿對5dpf和7dpf斑馬魚趨觸性行為的影響統一與差異共存與光照階段相比,我們觀察到所有濃度毛果蕓香堿處理的5dpf斑馬魚(包括對照組)在黑暗中焦慮水平的降低,但以中心區域游動時間百分比這一指標來衡量時發現黑暗刺激下5dpf斑馬魚同時存在焦慮隨毛果蕓香堿濃度變化而變化的現象,高濃度時相對增加了斑馬魚幼魚的焦慮水平。但無論以中心區域游動距離百分比還是中心區域游動時間百分比來衡量,7dpf斑馬魚的焦慮水平呈藥物劑量依賴性提高。5dpf和7dpf的斑馬魚之間明顯的差異可能反映了斑馬魚幼魚早期發育階段神經系統發育的差異。
在以往的研究中,5dpf的AB型斑馬魚幼魚在光暗交替試驗的黑暗階段表現出趨觸性行為的增加,而在黑暗階段給予微弱的照明時,可以減少趨觸性行為,表明黑暗環境比光照環境更易對斑馬魚幼魚形成壓力[8]。在課題組的多項研究中,斑馬魚對突然暗刺激的反應與Schn?rr 等的發現不一致[30-31]。我們的研究中,對照組的斑馬魚在光線突然發生轉變后更趨向于在測驗孔板的中心區域進行游動和探索,體現出在我們的暗刺激下斑馬魚壓力降低且對新環境更為新奇,趨觸性下降,焦慮水平降低。這種結果之間的差異可能是由于實驗設置中斑馬魚幼魚的遺傳背景、對黑暗適應程度以及溫度和測試時間等多種因素不同所導致。
我們研究了不同光照條件下毛果蕓香堿對5dpf和7dpf斑馬魚自發運動和趨觸性行為的影響并因此發現:在同樣的光照環境和毛果蕓香堿藥物濃度時,5dpf和7dpf的斑馬魚幼魚自發運動并不完全一致,提示毛果蕓香堿對斑馬魚幼魚自發運動的影響可能與斑馬魚的年齡相關;且毛果蕓香堿藥物濃度和斑馬魚幼魚年齡是焦慮模型中兩個關鍵因素,在我們的研究中,10 mmol/L的毛果蕓香堿對7dpf的斑馬魚幼魚有明顯的致焦慮作用。我們成功地建立了毛果蕓香堿誘導的斑馬魚焦慮模型,進一步為以斑馬魚為模式動物研究焦慮癥的發病機制和治療手段提供了更為便捷的方法。
目前,可方便地利用視頻跟蹤對斑馬魚運動軌跡進行精確的檢測統計及進一步數據分析,有助于在實驗中降低成本的同時提高效率,已廣泛應用于神經系統及行為學的研究評價中,視頻跟蹤系統的強大功能與斑馬魚模型相結合為模擬疾病的發生及探索疾病發病機制和治療方法提供了重要的方向。
