2014—2018 年豬流行性腹瀉病毒N 基因遺傳變化分析
2019-10-25 02:28:46邢廣旭焦文強劉運超王治方王克領徐引弟李海利張改平
山西農業科學 2019年10期
關鍵詞:研究
邢廣旭,焦文強,劉運超,王治方,王克領,徐引弟,李海利,張改平
(1.河南省農業科學院動物免疫學重點實驗室,河南鄭州450002;2.河南省農業科學院畜牧獸醫研究所,河南鄭州450002;3.河南省畜禽繁育與營養調控重點實驗室,河南鄭州450002)
豬流行性腹瀉(Porcine epidemic diarrhea,PED)是由豬流行性腹瀉病毒(Porcine epidemic diarrhea virus,PEDV)感染豬引起的一種高度接觸性腸道傳染性疾病,臨床主要表現為嘔吐、水樣腹瀉,最后脫水死亡。該病可感染各年齡階段的豬,7 日齡尤其是3 日齡以內的哺乳仔豬發病后死亡率最高可達100%[1]。該病在1971 年由英國首次報道[2],并且迅速傳播到歐洲其他國家,并在比利時流行期間被證實其病原為PEDV,給歐洲的養豬業帶來了重大損失[3]。亞洲的日本、韓國、泰國等國家都曾先后報道過PED 的流行[4-8]。我國學者宣華等[9]于1984 年首次報道PEDV 在我國豬場的存在,并且在2010 年以前主要是以散發為主。但是從2010 年10 月開始,我國南部多省份暴發了仔豬嚴重腹瀉,并迅速蔓延到全國多個省份和地區,測序結果證明,PED暴發是由變異的PEDV 引起[10-12];2013 年5 月,美國暴發了PED 疫情,并很快蔓延到美國全國,其分離株與我國安徽分離株(AH20122)親緣關系最為接近,表明PEDV 突變重組現象非常嚴重,同時這也是現有疫苗難以提供免疫保護的主要原因[13]。
PEDV 屬于冠狀病毒科(Coronaviridae)冠狀病毒屬(Coronavirus),核酸類型為單股正鏈RNA[14]。基因 組 由5′非 編碼 區(untranslated region,UTR)、3'UTR 以及7 個開放閱讀框(open reading frames,ORFs),分別編碼3 個非結構蛋白(non-structural proteins,NSPs)和4 個結構蛋白(structural proteins,SPs)組成,也就是S 蛋白、E 蛋白、M 蛋白和N 蛋白[15]。其中,N 蛋白是所有結構蛋白質中含量最多的蛋白,在PEDV 感染早期機體會產生大量針對N 蛋白的抗體,因此,N 蛋白被用于早期診斷[16]。N 蛋白在宿主細胞內誘導產生細胞免疫過程中扮演著非常重要的角色[17]。此外,N 基因參與病毒基因組的轉錄、病毒核衣殼的形成以及病毒RNA 的包裝[18-19]。
為深入了解PEDV 在我國遺傳變異的情況,本研究對2014 年10 月至2018 年3 月采集到的205 份PEDV 疑似病料進行RT-PCR 檢測,擴增陽性樣品的N 基因序列并進行測序,旨在為指導臨床PEDV的防控工作提供參考。
1 材料和方法
1.1 病料采集
本研究于2014 年10 月至2018 年3 月共采集來自河南、河北、山東、山西等地區疑似PEDV 病料205 份,包括仔豬小腸組織、糞便以及母豬血液等。
1.2 主要試劑
Taq DNA聚合酶、鼠源反轉錄酶(M-MLV)、RNA酶抑制劑、pMD18-T Simple 載體、DL2000 Marker RNA 提取試劑盒,均購自大連寶生物工程有限公司;瓊脂糖購自Oxoid 公司;其他試劑均為國產分析純。
1.3 引物設計
參考GenBank 中收錄的PEDV 毒株N 基因序列,使用Primer Premier 5.0 軟件設計引物,上游引物為:5′-TACGGATCCATGGCTTCTGTCAGC-3′,下游引物為:5′-TCGCTCGAGTTAATTTCCTGTATC-3′,引物由生工生物工程(上海)股份有限公司合成。
1.4 PEDV 的N 基因的克隆與測序
按照寶生物RNA 提取試劑盒操作說明書完成疑似樣品的RNA 提取,隨即合成cDNA,反應體系為:5×NLV 反轉錄緩沖液2 μL,dNTP 2 μL,MLV 反轉錄酶0.5 μL,rNAse 抑制劑0.3 μL,總RNA4.9 μL,充分混勻后置離心機瞬時離心混勻,42 ℃1.5 h,70 ℃10 min。以cDNA 為模板進行PCR 擴增,PCR反應體系為:Taq DNA 聚合酶1 μL,cDNA 1 μL,上下 游 引 物 各1 μL,dNTP 4 μL,10×PCR Buffer(Mg2+plus)5 μL,無菌雙蒸水補足50 μL。反應程序為:94 ℃預變性5 min;94 ℃30 s,57 ℃30 s,72 ℃1 min,35 個循環;最后72 ℃延伸10 min。擴增結束后取5 μL PCR 產物于1.0%瓊脂糖凝膠電泳檢測擴增結果,回收與預期大小一致的PCR 產物,并與pMD18-T simple 載體連接,4 ℃過夜后轉化至JM109 感受態細胞,藍白斑篩選后挑取單克隆進行培養,并提取質粒送至生工生物工程(上海)股份有限公司進行測序。
1.5 數據處理
使用MEGA 5.0 軟件構建系統進化樹。
2 結果與分析
2.1 PEDV 的N 基因的擴增
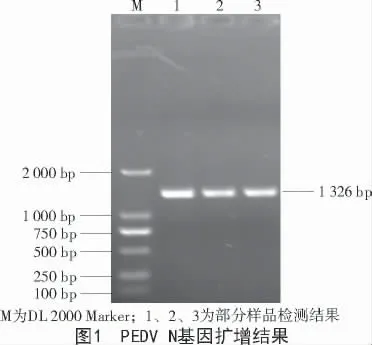
205 份待檢樣品中,有191 份樣品檢測為PEDV 陽性,陽性率高達93.17%。從圖1 可以看出,PCR 擴增PEDV N 基因的引物可以擴增出1 326 bp大小的片段,片段符合預期大小。
2.2 PEDV 的N 基因比對結果
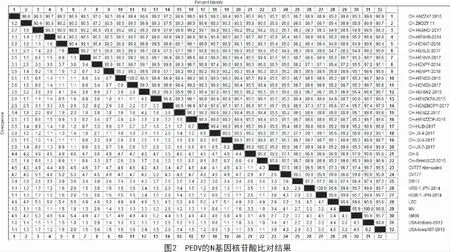
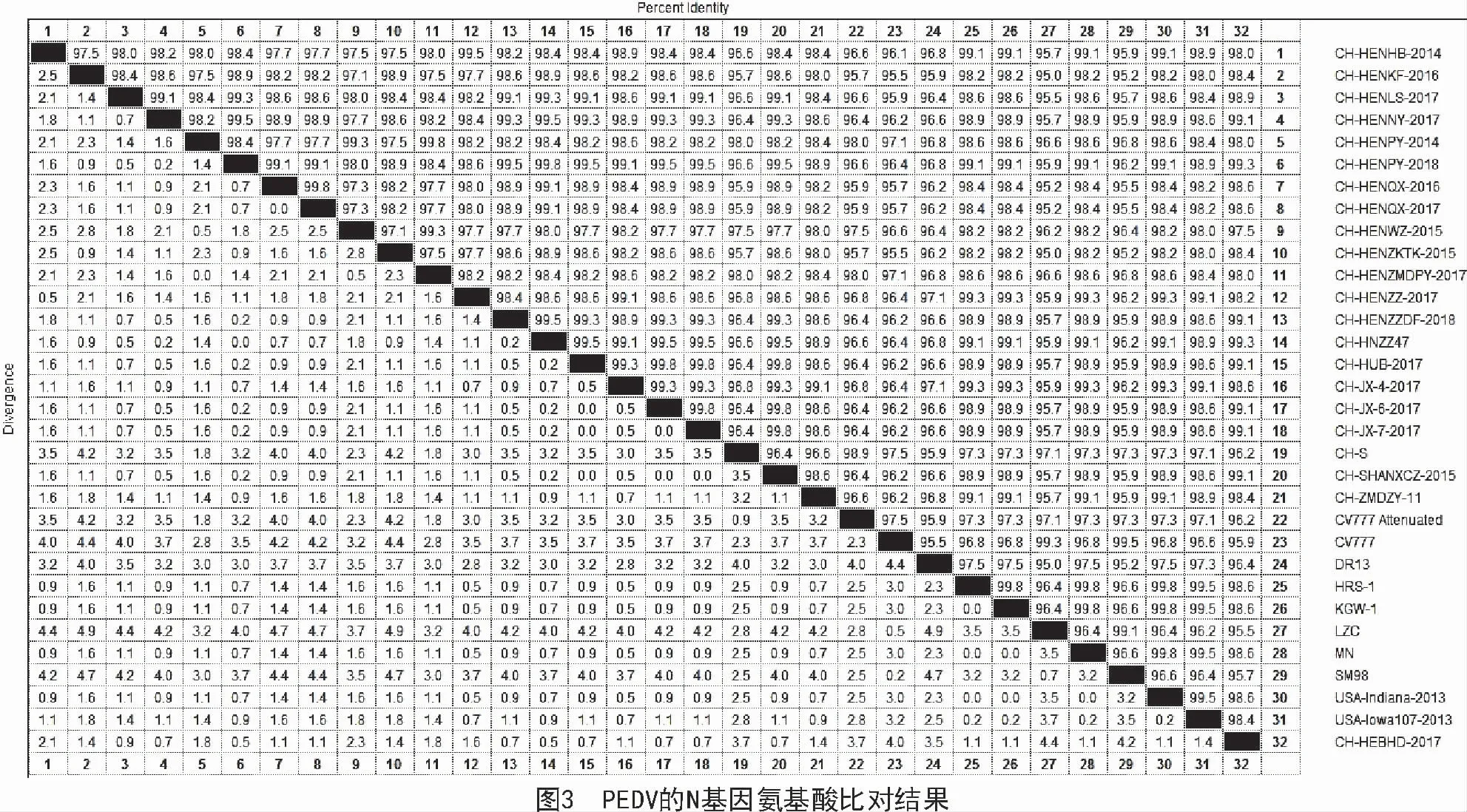
本研究得到的19 株N 基因序列與國內外參考毒株進行比對,發現這19 株N 基因序列與國內外代表毒株相比,其核苷酸的同源性為94.8%~99.5%(圖2),氨基酸的同源性為95.0%~99.3%(圖3)。
2.3 PEDV 的N 基因的進化分析
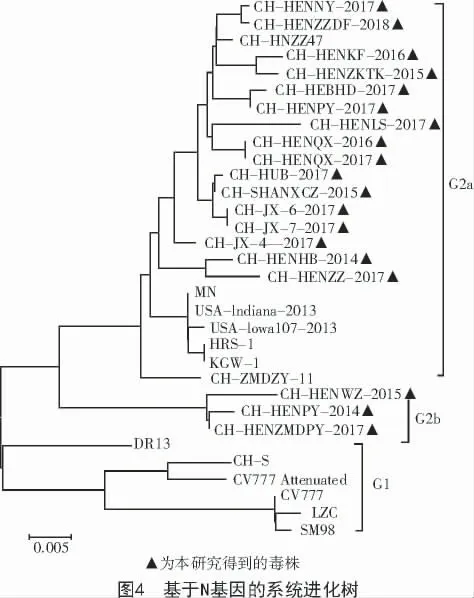
使用MEGA 5.0 軟件,將本研究得到的19 株PEDV 的N 基因序列與GenBank 中下載的序列構建系統進化樹(圖4)。從圖4 可以看出,本研究得到的19 株PEDV 的N 基因序列與CV777、LZC、SM98、CH-S 和DR13 等毒株不在同一個分支上,表明這19 株N 基因序列與傳統經典毒株進化關系較遠;而CH-HENWZ-2015、CH-HENPY-2014、CH-HENZMDPY-2017 3 株序列形成一個分支,為G2b 亞群;其余16 株N 基因序列形成另外一個分支,為G2a亞群。表明N 基因進化趨勢日益復雜,需要加強此方面的監控。
3 結論與討論
通過RT-PCR 方法,本研究得到了19 株PEDV 的N 基因序列,與先前的報道一致。本研究得到的PEDV 的N 基因由1 326 個核苷酸組成,共編碼441 個氨基酸,分子量為56 ku 左右,沒有任何堿基的缺失和插入。有研究顯示,PEDV 基因組有4 個超變區[20],其中,V4 位于N 蛋白內,這與以前的結論不一致[21]。
本研究得到的19 株PEDV 的N 基因序列與GenBank 中下載的參考序列核苷酸的同源性為94.8%~99.5%,氨基酸的同源性為95.0%~99.3%。考慮到本研究所采集的樣品來自河南、河北、山西、山東、湖北、甘肅、江西、上海等多個省市,并且本研究所采集的樣品一共205 份,樣本數量足夠大,而且檢測出191 份PEDV 陽性樣品,所以,不管是樣品的采集數量還是樣品的覆蓋面積,均具有較強的說服力,證明N 基因是PEDV 基因組中很保守的一個結構蛋白。氨基酸比對結果發現,本研究得到的19 株N 蛋白氨基酸序列主要發現以下位點的突變:K123-N123,A142-T142,R241-K241,K252-R252,N255-S255,M394-T394,E400-D400,A408-L408,V402-S402,從而從氨基酸水平證實N 基因缺失很保守。
采用MEGA 5.0 軟件構建的基于N 基因的系統進化樹發現本研究得到的19 株N 基因序列與CV777、LZC、SM98、CH-S 和DR13 等國內外代表性毒株不在同一個分支上,表明這19 株N 基因序列與傳統經典毒株進化關系較遠;其中,16 株N 基因序列與MN、USA-Indiana-2013、USA-Iowa107-2013、HRS-1、KGW-1、CH-ZMDZY-11 形成一個分支,為G2a,CH-HENWZ-2015、CH-HENPY-2014、CHHENZ-MDPY-2017 這3 株序列形成另一個分支,為G2b。綜上所述,表明我國目前PEDV 的N 基因與以往流行的經典毒株親緣關系較遠,大部分毒株與目前國內外分離的毒株在一個進化分支中,部分毒株獨立進化成一個分支。表明PEDV 遺傳進化趨勢日益復雜,需要長期監控。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
