天祝縣燕麥葉斑病的發生情況及其病原鑒定
2019-11-07 11:55:08聶秀美趙桂琴孫浩洋柴繼寬吳文斌孫雷雷王苗苗康曉強
草地學報 2019年5期
聶秀美,趙桂琴,孫浩洋,柴繼寬,吳文斌,林 龍, 孫雷雷,王 軍,王苗苗,康曉強
(甘肅農業大學草業學院,草業生態系統教育部重點實驗室,中-美草地畜牧業可持續發展研究中心,甘肅 蘭州 730070)
燕麥(Avenasativa)是禾本科燕麥屬一年生世界性栽培作物[1],具有廣泛的生態適應性、豐富的營養價值和獨特的保健功能[2-3],在我國主要分布于西南、華北、西北等高寒冷涼地區[4],對我國畜牧業生產、農業結構調整、生態環境改善等方面具有重要意義[5-6]。
燕麥葉斑病(Drechsleraavenacea)又稱條紋葉枯病,主要為害燕麥葉片和葉鞘,發病初期病斑呈灰綠色水漬狀,隨著病情加重,漸變為淺褐色至紅褐色橢圓形病斑。在低溫高濕環境中,病斑還可擴展成不規則形條斑甚至融合成片,導致葉片枯死脫落,嚴重影響燕麥的生產性能和飼用價值[7]。其病原菌多以分生孢子或菌絲在田間病殘體上越冬,第2年回春時產生的病原物可從幼嫩組織侵入燕麥植株體,發病后的燕麥植株所攜帶的致病菌菌絲或分生孢子又可反復進行多次侵染[8]。該病害在燕麥重病田的發病率高達90%以上,在美國、加拿大、波蘭等國家都是燕麥生產中的主要病害,在巴西被認為是流行最快的病害,可危害燕麥的幼苗、葉片及種子,一般減產5%~10%,重者可達30%以上[9-11]。
燕麥葉斑病盡管在很多國家都有發生,但都鮮見報道,我國對這種病害的研究報道更少,僅見袁軍海等[12]于2013對河北省張家口市的51個燕麥品種進行葉斑病抗性鑒定,其結果表明不同品種間的抗病性差異較大,病葉率為70.83%~100%,病情指數為20.31~84.87,僅4個燕麥品種表現為中度抗病。東保柱[13]在內蒙古武川縣和河北壩上對兩地共有的39個主栽燕麥品種進行研究表明,不同品種對葉斑病的抗性不同,同一品種在不同地區的抗性表現也存在差異,如‘白燕2號’在河北表現為免疫,在內蒙古表現為髙抗;‘壩燕4號’、‘壩莜8號’、‘埂莜5號’在河北表現為中感,在內蒙古表現為高感。劉萬友[14]對不同栽培條件下燕麥葉斑病的發病情況進行研究表明,播期是影響該病害的最主要因素,通過利用不同殺菌劑進行試驗表明40%的多菌靈對該病害具有較好的防治效果,且拌種處理較葉面噴施效果更好。張笑宇等[15]采用組織分離法從內蒙古自治區武川縣和河北省張家口市張北縣采集的燕麥葉斑病病葉中共分離培養出3株菌株,鑒定表明這3株病原菌均為燕麥內臍蠕孢菌(Drechsleraavenae),其有性態為燕麥核腔菌(Pyrenophoraavenae)。天祝藏族自治縣地區是甘肅省的主要畜牧業基地之一,具有典型的農牧交錯區特點。燕麥是該區域主要的栽培牧草,也是當地家畜冷季補飼的主要飼草來源。但近年來,該地區燕麥葉斑病逐年加重,嚴重影響當地燕麥產業的發展。因此,本研究擬通過對天祝縣燕麥葉斑病的發生情況進行調查,對采集到的病葉進行病原菌的分離和鑒定,以明確葉斑病在天祝縣的發生情況,確定病原菌種類,為當地燕麥葉斑病病害的進一步研究和防治提供理論依據。
1 材料與方法
1.1 病害調查
于2018年7月對甘肅省天祝縣松山鎮黑馬圈河村(103°40′ E,37°17′ N,海拔2 885 m)、東大灘鄉上泉村(103°42′ E,37°20′ N,海拔2 699 m)和酸茨溝村(103°30′ E,37°25′ N,海拔2 653 m)、安遠鎮柳樹村(102°85′ E,37°25′ N,海拔2 708 m)、抓喜秀龍鄉南泥溝村(102°79′ E,37°21′ N,海拔2 866 m)種植的甜燕麥品種進行了田間葉斑病發生情況的調查。每個點隨機選取3塊樣地,避開地邊5 m,采用“Z”字型5點取樣法,每點調查燕麥植株的上部、中部、下部共計50片葉,逐片記錄發病情況,并進行病葉率和病情指數的計算。葉斑病的病葉率、病情指數的計算和評價標準參照袁軍海等[12]的計算方法和評價標準,病級劃分標準見表1。
病葉率(%)=病葉數/調查葉片總數×100
式中:S為病情指數;i為病級數(1~n);Xi為病情為i級的單元數;Si為病情為i級的嚴重度值。
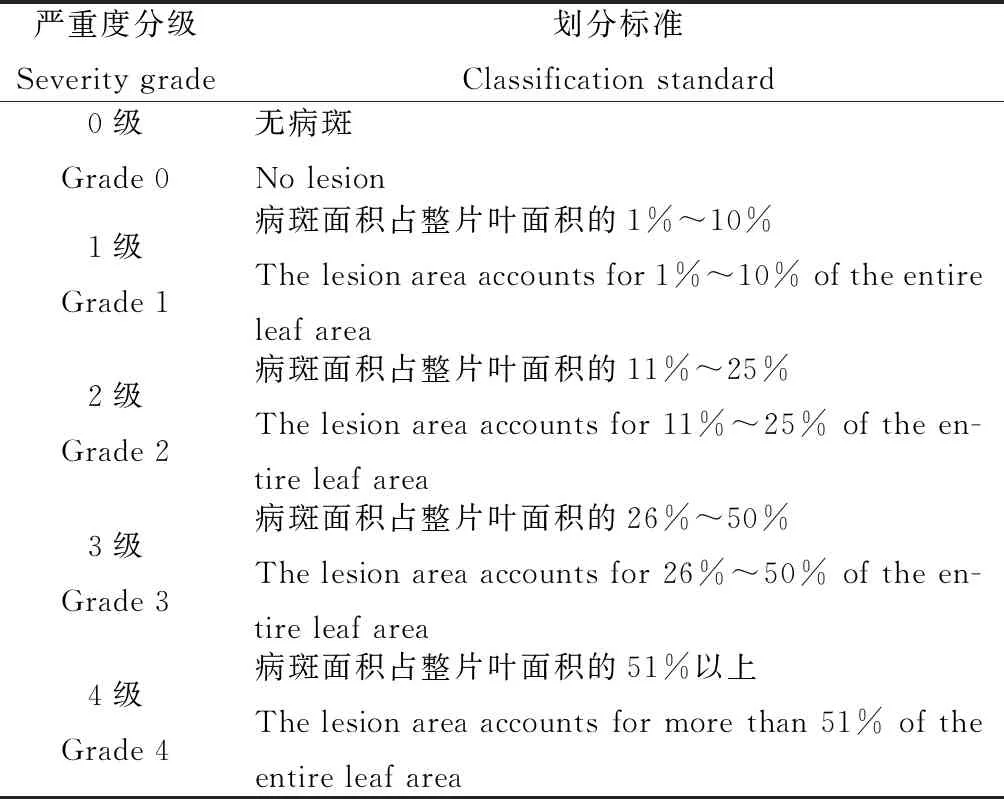
表1 燕麥葉斑病病級劃分標準
1.2 病原菌鑒定
1.2.1病原菌的分離培養 將采集的燕麥病葉標本帶回實驗室后,參照Xue等[16]和李戌清等[17]的方法在病健交界處選取具有典型葉斑病癥狀的組織,經自來水沖洗,用70%酒精表面消毒1 min,5%次氯酸鈉消毒5 min,無菌水沖洗3~5次后用滅菌濾紙吸干表面水分,再剪成5 mm的小塊,隨機選擇5塊均勻放在直徑90 mm的馬鈴薯葡萄糖瓊脂(Potato dextrose agar,PDA)平板上,5皿為1重復,4次重復,置25℃恒溫培養箱中黑暗培養。待長出菌絲后及時進行單菌分離和純化,純化后的菌株用20%的甘油于—20℃冰箱保存備用。
1.2.2病原菌的致病性鑒定
離體葉片接種 張莉等[18]和張笑宇等[15]的方法進行病原菌的致病性鑒定。選取健康、無病斑的燕麥葉片,自來水沖洗干凈,剪取葉片中間5 cm長的葉段,用70%酒精消毒1 min,5%次氯酸鈉消毒5 min,無菌水沖洗3~5次后用滅菌濾紙吸干葉片表面的水分。用無菌打孔器在培養7 d的菌株上取直徑5 mm的待測菌株塊接種于燕麥葉片表面,每皿放4片葉段,每處理重復4次,以接種相同大小的新鮮PDA培養基塊為對照。于25℃恒溫培養,及時觀察發病情況,第4 d對發病燕麥葉片組織進行病原菌的再分離,檢驗接種前后病原菌是否一致。
活體葉片接種 每盆留苗10株。當燕麥長至3~4葉期時,選取大小一致的3株幼苗,用無菌打孔器在培養7 d的菌株上取直徑5 mm的待測菌株塊,接種在燕麥葉片第3葉中間,用紗布和保鮮膜固定菌餅,每處理重復4次,以接種相同大小的新鮮PDA培養基塊為對照。接種后的燕麥葉片用塑料袋罩住置于25℃,黑暗14 h、濕度95%,光照10 h、濕度75%的恒溫保濕培養箱黑暗光照交替培養48 h后取下塑料袋繼續培養,觀察發病情況。第7 d時對病斑進行病原菌的再分離,檢驗接種前后病原菌的一致性。
1.2.3病原菌的形態學鑒定 將純化后的病原菌接種在新鮮PDA平板上于25℃黑暗培養7 d后,觀察菌落和分生孢子的形狀、顏色和大小等形態學特征進行初步鑒定。
1.2.4病原菌的分子生物學鑒定 采用真菌基因組Omega試劑盒提取菌株DNA,以真菌特異性引物對ITS1和ITS4進行PCR擴增,反應條件為:94℃預變性5 min;94℃變性30 s,52℃退火30 s,72℃延伸1 min,34個循環后在72℃下再延伸7 min。取PCR擴增產物5 μL,用1%的瓊脂糖凝膠進行電泳檢測后送樣進行測序,所得序列經Clustal W軟件進行堿基校對后,與GenBank中已知序列進行相似性比對,用Mega 7.0軟件經Bootstrap 1000次循環檢驗后以鄰接法構建系統發育樹。
2 結果與分析
2.1 葉斑病的發生情況
通過對天祝縣葉斑病發生情況調查發現,各燕麥種植區域的葉斑病發病情況存在明顯差異(表2),發病率為64.20%~99.58%,病情指數介于23.00~61.81之間。松山鎮黑馬圈河村和東大灘鄉上泉村的發病情況最嚴重,前者的葉斑病病葉率高達92.78%,病情指數為61.81;后者的病葉率最高,為99.58%,病情指數也較高,為60.02。柳樹村、南泥溝村、酸茨溝村的發病也較重,其中柳樹村和酸茨溝村的發病程度接近,病葉率均為64.51%,病情指數為25.47。南泥溝村的病葉率和病情指數相對較低,分別為64.20%和23.00。
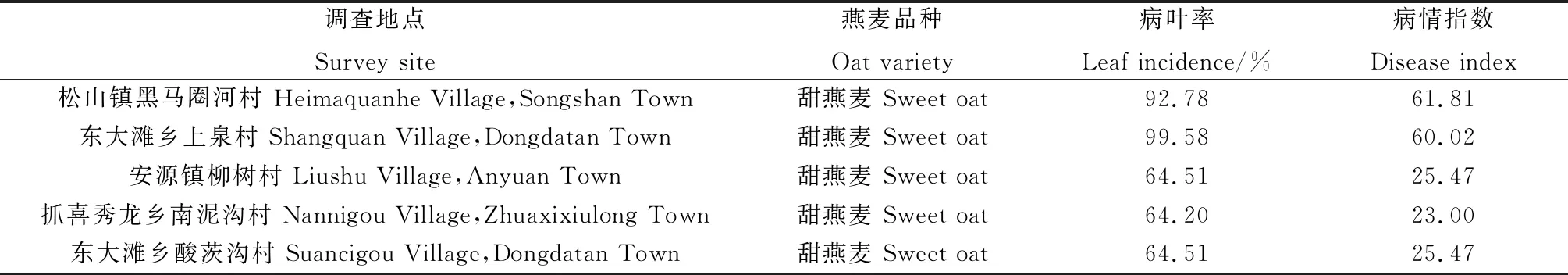
表2 天祝縣地區燕麥葉斑病的發生情況
2.2 病原菌的致病性測定
對從燕麥葉斑病病葉上分離獲得的菌株通過離體葉片和活體葉片致病性接種試驗發現,有6株菌株對燕麥葉片具有致病性,將其依次編號為AS-L-1~AS-L-6。圖1為病原菌的致病性表現。離體葉片接種2 d時(圖1 A~D),菌餅周圍布滿菌絲,葉片接種部位開始出現輕微的褪綠癥狀,部分接種菌餅的葉片表面逐漸浮現出輕微的病斑,對照無任何變化;接種4 d時(圖1 E~H),菌餅面積明顯擴大,葉片出現大面積褪綠現象,接種部位有明顯的病斑,葉片表面也開始出現病斑,而對照不發病。活體葉片接種3 d時,接種部位開始顯癥,有輕微的褪綠現象,葉片表面開始出現菌絲,而接種PDA培養基的對照葉片未發病;接種5 d時,接種部位褪綠現象加重,并出現淺褐色或褐色小斑點;接種7 d時(圖1 I),病斑面積明顯擴大,病斑呈褐色或黃褐色,對照未發病。將發病燕麥離體葉片和活體葉片進行病斑再分離,發現所得病原菌與接種病原菌的培養性狀和形態特征相同,符合致病菌的柯赫氏法則驗證,說明這6株菌株均為燕麥葉斑病的致病菌。
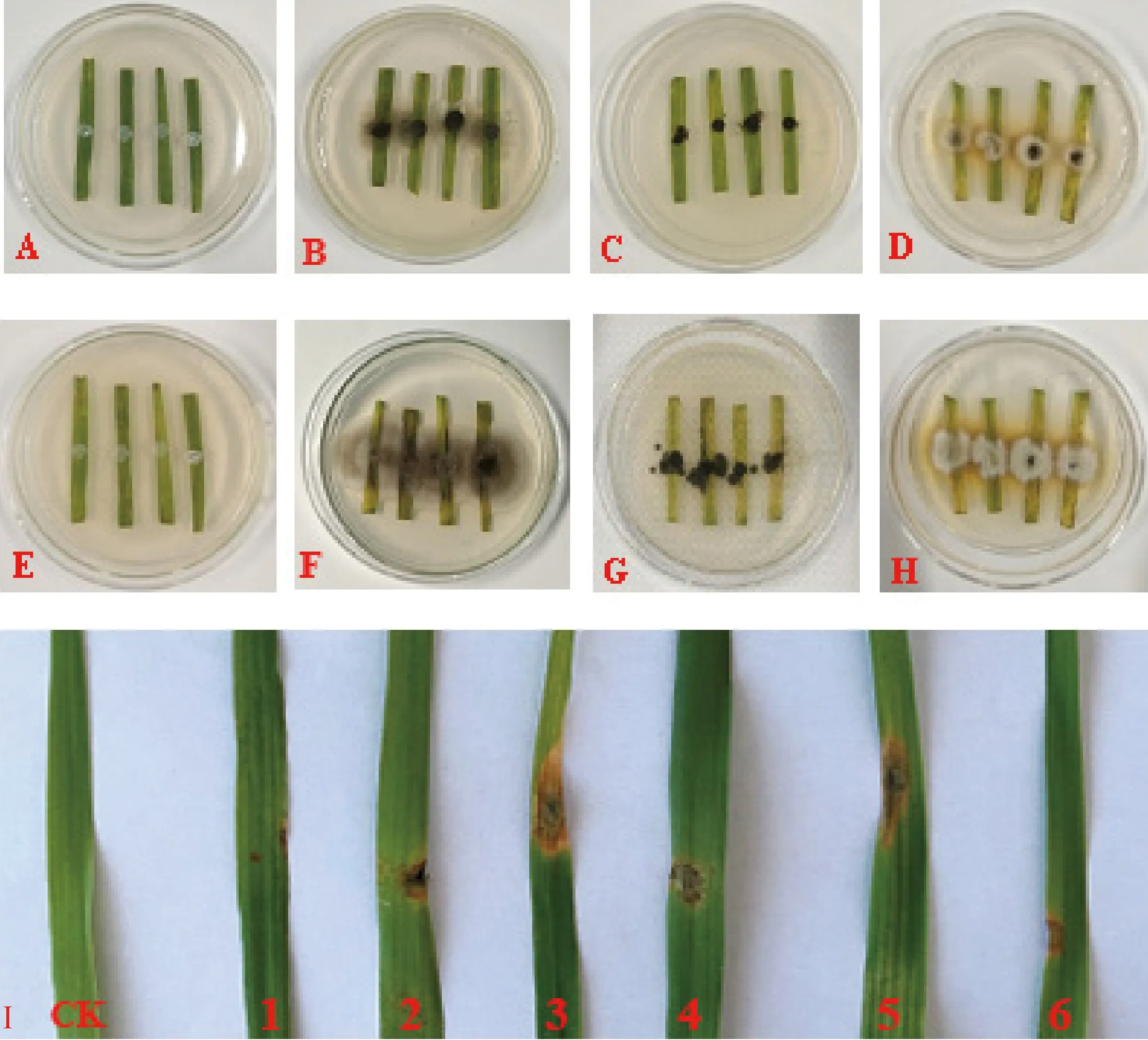
圖1 燕麥離體葉片和活體葉片的接種癥狀
2.3 病原菌的形態特征
對分離到的燕麥葉斑病病原菌在PDA培養基上純化培養7 d后,進行形態觀察,發現其菌落培養特征和孢子形態特征明顯不同,部分病原菌的菌落和孢子形態如圖2所示。
細交鏈孢(圖2 A~C)菌落生長速度較快,稍隆起,近似圓形,直徑約62 mm,表面粗絨狀,呈灰褐色,有同心圓,邊緣整齊,為白色,氣生菌絲濃密。背面黑褐色,邊緣褐色。分生孢子單生或串生,倒棍棒狀,倒梨形或橢圓形,淡黃褐色至褐色,大小為(9.10~24.20)μm×(3.40~8.50)μm,具縱橫隔膜,橫隔1~6個,縱隔0~3個。
芽枝狀枝孢菌(圖2 D~F)的菌落生長速度較慢,平鋪,圓形,直徑約30 mm,表面粉狀,邊緣整齊,氣生菌絲稀疏,有放射狀紋理,整體呈深綠色,背面黑綠色。分生孢子橢圓形,橄欖色,大小為(4.30~8.50)μm×(2.20~4.80)μm,無隔膜。
黑附球菌(圖2 G~I)的菌落生長速度緩慢,均勻隆起,近似圓形,直徑約25 mm,表面絨狀,呈米黃色,氣生菌絲濃密,背面焦糖色。分生孢子單生,球形,黃褐色至褐色,大小為1.10 μm×2.50 μm,無隔膜。
燕麥內臍蠕孢(圖2 J~L)的菌落生長速度較快,平鋪,近似圓形,直徑約68 mm,表面粗絨狀,中央淺棕色至棕色,邊緣乳白色,氣生菌絲稀疏,背面中央黑褐色。分生孢子單生,圓柱狀,褐色,大小為(10.52~32.47)μm×(3.24~9.63)μm,具橫隔3~7個。
2.4 病原菌rDNA-ITS序列系統進化分析
采用真菌ITS區通用擴增引物對ITS1和ITS4對6株致病菌的DNA進行擴增測序,并通過GenBank數據庫對這6個菌株的堿基序列同已知物種序列進行同源性比對,結果表明,各比對樣品與數據庫中序列相似性均在98%以上。進一步采用Mega 7.0軟件用鄰接法構建系統發育樹進行物種間序列比對分析(圖3),結果表明菌株AS-L-1~AS-L-6的rDNA-ITS序列依次與細交鏈孢(Alternariaalternata(MF1011))、芽枝狀枝孢菌(Cladosporiumcladosporioides(KY400091))、燕麥內臍蠕孢(MK632061)、黑附球菌(Epicoccumnigrum(MK632017))、燕麥核腔菌(MK044614)和德氏霉菌(Pyrenophorachaetomioides(MH3992780)) 的親緣關系最近,單獨形成分支。結合形態特征,最終確定AS-L-1~AS-L-6這6個致病菌株分別為細交鏈孢、芽枝狀枝孢菌、燕麥內臍蠕孢、黑附球菌、燕麥核腔菌和德氏霉菌。
通過對6個菌株的rDNA-ITS序列進行聚類分析(圖4)表明,6株菌株分屬于4個真菌屬,其中德氏霉屬(Drechslera)3株,分別為燕麥內臍蠕孢、燕麥核腔菌和德氏霉菌;鏈格孢屬(Alternaria)、附球菌屬(Epicoccum)和枝孢屬(Cladosporium)各1株,分別為細交鏈孢、芽枝狀枝孢菌和黑附球菌。
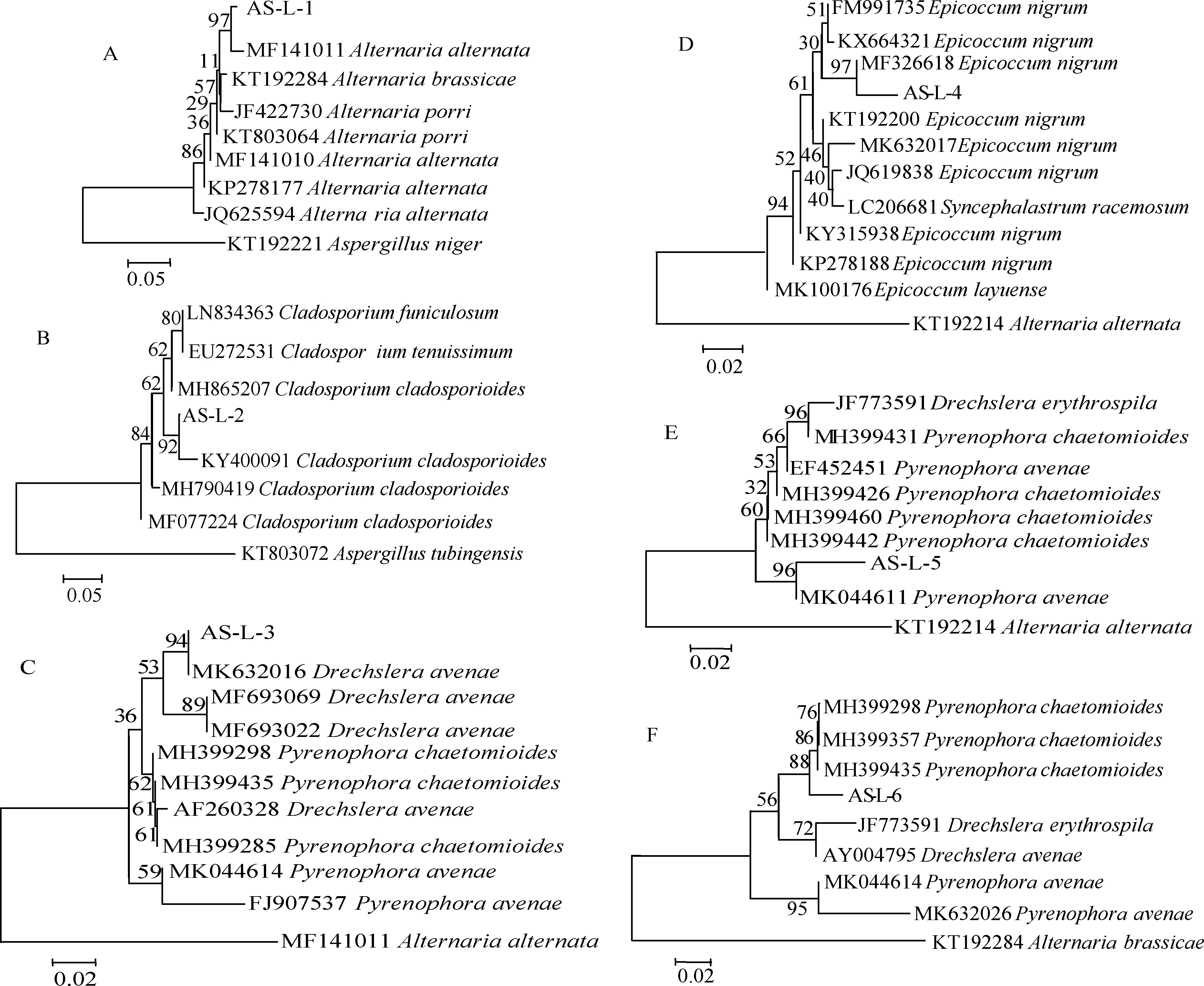
圖3 6個病原菌株的rDNA-ITS序列同已知物種同源序列的系統進化分析
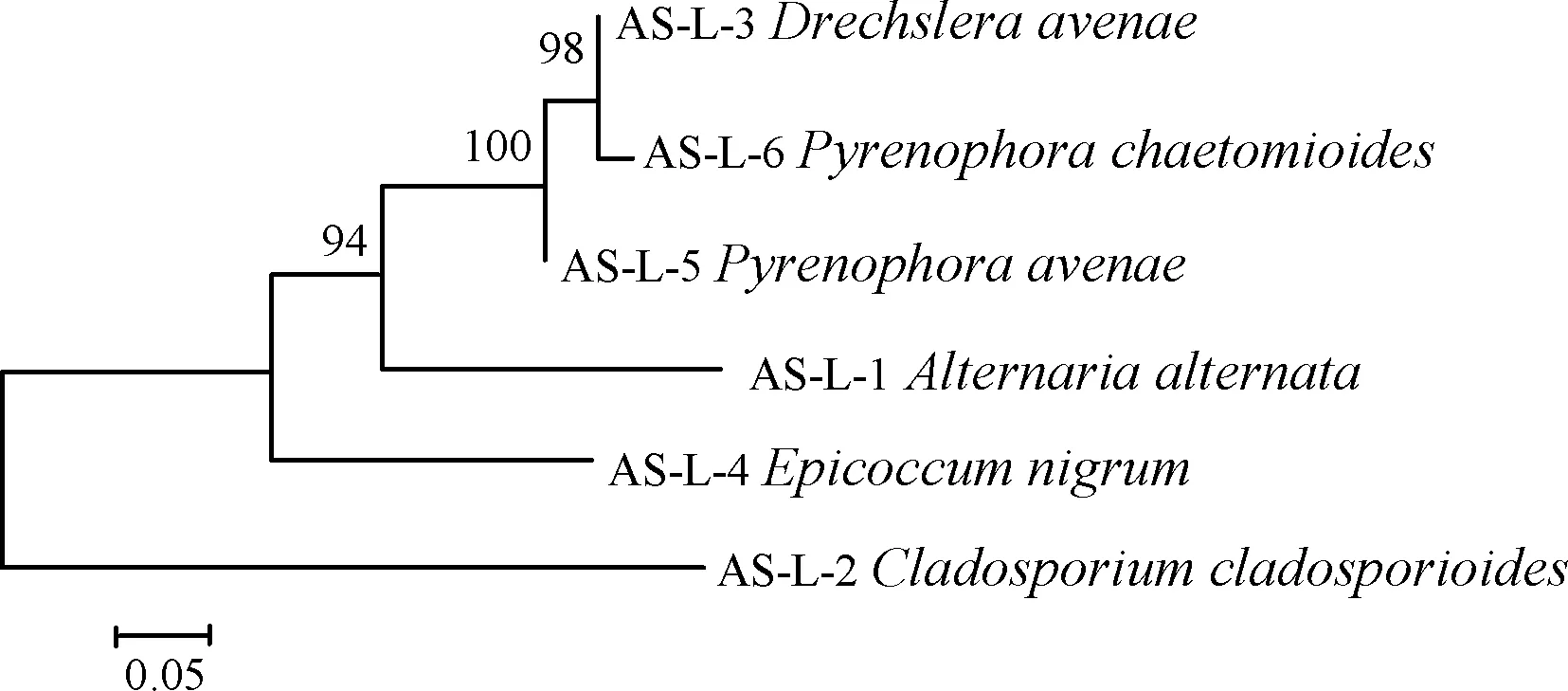
圖4 菌株AS-L-1~AS-L-6的rDNA-ITS序列系統發育樹
2.5 病原菌的分布情況
由表3可知,不同來源的供試材料攜帶的病原菌種類存在明顯差異,來自安源鎮柳樹村的材料攜帶的病原菌株最多,為6株;其次是松山鎮黑馬圈河村和抓喜秀龍鄉南泥溝村的材料,除未攜帶芽枝狀枝孢菌和黑附球菌外,其它4株病原菌均能在這2份材料中檢出;東大灘鄉上泉村和酸茨溝村材料中分離出的病原菌株相對較少,都僅有3株。另外,從各供試材料中均可分離出細交鏈孢和燕麥內臍蠕孢,而芽枝狀枝孢菌僅在安源鎮柳樹村材料中分離到。燕麥核腔菌除從東大灘鄉上泉村材料中未檢出外,在其它材料中均可檢測到。
3 討論
作物在整個生長發育過程中,都會受到各種病害的危害,這些病害主要由病原菌侵染引起[19]。燕麥葉斑病是一種真菌性病害。整個生物界由病原真菌引發的真菌病害約占植物病害的70%~80%,這些真菌具有分布廣、種類多、發病快、隱蔽性強、危害重等特性,通常會侵入植物的質外體或細胞獲取營養,形成寄生關系,從而使植物致病[20-21]。
病葉率和病情指數是反映植物病害發生情況或危害狀況的重要指標。本試驗通過對天祝縣5個燕麥種植區的葉斑病發生情況調查發現,其病葉率在64.20%~99.58%之間,病情指數變化范圍為23.00~61.81,其中黑馬圈河村和上泉村葉斑病發病最嚴重,其它3個調查區域的發病也較重,這與金柳艷等0對在不同環境下的5個相同玉米(Zeamay)品種的病害調查結果類似。植物與病原菌的相互作用是一個復雜的生物學過程,植物對病原菌的防御能力除受植物本身遺傳特性的影響外,還可能與其自身對不同地理環境的適應性和管理措施有差異有關[23-24]。本研究通過對不同來源的供試病樣進行致病菌檢測后發現,在柳樹村材料中檢出的病原菌種類最多;黑馬圈河村和南泥溝村次之;上泉村材料中分離出的病原菌最少,這可能是因為葉斑病致病菌在低溫高濕環境中易發生侵染和傳播,而且其發生程度與播期、種植密度、施肥量、品種和灌水量等田間管理措施密切相關,但由于在不同地理環境條件下病害發生和流行的致病環境因子存在差異,因此不同調查區域的病害致病菌有所不同[25-26]。本研究是對天祝縣不同燕麥種植區域同一燕麥品種的病害情況進行調查,可以說明造成其發病情況存在差異的主要因素是區域間的環境因子和管理措施。

表3 燕麥葉斑病病原菌的分布情況
注:“+”表示從供試材料中可分離到該菌,“-”表示從供試材料中未分離到該菌
Note:“+” indicates the fungus was isolated from the tested materials,“-” indicates the fungus was not isolated from the tested materials
采用柯赫氏法則、傳統形態學分類法和rDNA-ITS序列分析法可以準確鑒定致病真菌種類,克服傳統分類鑒定方法中致病菌株培養數量多、形態特征復雜和易受外界環境條件影響等因素,從而可以對病原菌進行快速準確的診斷和鑒定[27-28]。本研究通過對燕麥葉斑病病原菌分離物經致病性鑒定、形態學鑒定和分子生物學鑒定,共發現5種不同種類的致病菌,這些菌株與其各自所對應的GenBank庫中的序列同源性均在98%以上,具有很近的親緣關系。進一步對其進行聚類分析后發現,這些致病菌株分為4個真菌屬,其中德氏霉屬3株,鏈格孢屬、附球菌屬和枝孢屬各1株。目前,國內僅對由燕麥內臍蠕孢引起的燕麥葉斑病有所報道[15],對本研究發現的細交鏈孢、芽枝狀枝孢菌和德氏霉菌引起的燕麥葉斑病還未見報道。但已有研究表明,這些病原菌可在高粱(Sorghumbicolor)[29]、芒果(Mangiferaindica)[30]和小麥(Triticumaestivum)[31]等植物上引起葉斑病。因此,本研究新發現的這些病原菌為國內首次報道,關于這些病原菌在燕麥上的發生規律、致病機理及防治措施等還需進一步研究。
基于ITS基因構建的系統發育樹對于不同屬的真菌鑒別能力較強,在種水平上也有較好的鑒別力[32]。本研究也證實了這一點,能夠將待檢測菌株在屬和種水平上分開。細交鏈孢和黑附球菌的寄主范圍較廣泛,易造成嚴重危害,而且這些病原菌可以通過侵染田間雜草完成其生活史或者越冬,在適宜條件下,進行病原菌的再次傳播,成為葉斑病的侵染源[33-34]。因此,及早重視其潛在威脅,加強該病害致病機理方面的研究,篩選和使用高效低毒殺菌劑或生防菌劑,進行田間雜草防除是控制燕麥葉斑病的重要措施。另外,本研究在病原菌的分離過程中發現,病原菌以單獨侵染為主,但存在2~3種共同侵染燕麥葉片的現象。這與已有的研究結果類似[35-36],這可能是因為致病菌對外界環境的適應性具有相似性,能夠進行生長和繁殖,而且某些致病菌在產生病斑后對其他病菌的侵入有誘導作用,使復合侵染成為可能[37-38]。因此,還需對燕麥葉斑病致病菌的復合侵染規律進行明確,從而能夠制定更好的防控措施。
4 結論
天祝縣燕麥種植區的葉斑病發生較嚴重。5個村鎮葉斑病的發病率為64.20%~99.58%,病情指數為23.00~61.81。其中松山鎮黑馬圈河村和東大灘鄉上泉村發病最嚴重,其病葉率在92%以上,病情指數60以上。引起天祝縣燕麥葉斑病的病原菌共有4屬5種,分別為燕麥內臍蠕孢、細交鏈孢、芽枝狀枝孢菌、黑附球菌和德氏霉菌。
