轉Bar基因紫花苜蓿雜交組合的篩選與評價
2019-11-08 07:48:22王英哲徐安凱
草地學報 2019年5期
周 仂,王英哲,徐 博,譚 晶,徐安凱*
(1. 吉林農業大學動物科學技術學院,吉林 長春 130118; 2. 吉林省農業科學院,吉林 長春 130118; )
紫花苜蓿(Medicagosativa)是起源于伊朗的一種產量高、營養品質優良的牧草,在我國各地均有分布[1],在美國不足200年的栽培歷史中,已成為第四大栽培作物,年經濟收入超過100億美元[2-5]。紫花苜蓿在我國已有2000多年的栽培歷史,2019年中央1號文件提出“合理調整糧經飼結構,發展青貯玉米、苜蓿等優質飼草料生產”等目標任務,這為紫花苜蓿產業的發展提供了新的動力。
隨著植物基因工程的迅速發展,植物轉基因技術在紫花苜蓿遺傳改良方面發揮著愈加關鍵的作用[6]。除草劑抗性一直是轉基因作物的主要特征。如今轉基因作物的全球種植面積已經接近2億公頃,且其中抗除草劑作物的種植面積占80%[7]。目前,抗草銨膦基因被較早利用的抗性基因有2種,分別是Bar基因(Bialaphos resistance gene,Bar)和Pat基因(Phosphinothricin N-acetyltransferase gene,Pat),這2種基因已經通過在水稻中的轉化成功應用于商業化生產中[8]。陳倩楠等[9]通過農桿菌轉化法將Bar基因穩定整合到谷子‘冀谷11’的基因組中。劉艷芝等[10]將Bar基因轉入‘草原1號’苜蓿中,經篩選驗證,證實得到的轉基因植株對除草劑具有抗性。
傳統的轉基因作物通過回交進行育種,并且需要依靠分子標記的輔助。丁曉艷等[11]通過回交育種,將FBP7::iaaM基因導入短季棉品種‘晉棉11’,獲得了相比親本纖維產量提高的轉基因回交后代JBC4。王文靜等[12]應用分子跟蹤檢測技術,對4個T4代轉TaEBP,GhDREB,GmDREB1,GmDREB3基因的小麥通過有限回交、滾動回交育種,獲得抗旱相關轉基因“矮敗周麥18”小麥新種質。
利用雄性不育系育種,可以省去人工去雄的工作量,降低生產成本,提高經濟效益。雄性不育系在普通雜交育種方面具有廣泛的應用,可明顯提高F1代雜交種子生產質量與產量,縮短育種周期[13],但其在轉基因作物育種上鮮有應用。
本試驗以吉林省農科院的選育出的優良紫花苜蓿雄性不育系(MS-GN-1A)作為母本,篩選優異的抗除草劑轉基因紫花苜蓿‘公農1號’為父本,配置雜交組合。對紫花苜蓿的雜交組合F1代分子水平以及抗除草劑特性等方面進行檢測和相關分析,結合農藝學和品質性狀篩選出抗除草劑的優異雜交組合,為轉基因紫花苜蓿雜交種選育奠定基礎。
1 材料與方法
1.1 試驗材料與試驗器材
1.1.1試驗材料 植物材料:‘公農1號’紫花苜蓿(由吉林省農業科學院牧草生物技術育種研究室提供),雄性不育系MS-GN-1A(由吉林省農業科學院草地所提供)。以普通‘公農1號’紫花苜蓿為父母本配置1個對照組合(CK),以紫花苜蓿雄性不育系MS-GN-1A為母本,以11個農藝學性狀有差異的抗除草劑紫花苜蓿株系為父本,配制11個雜交組合。組合配置編號見表1。
1.1.2試劑 采購由上海佑隆生物公司生產,編號為AS-013-LS的PAT/Bar轉基因速測試紙;采購由北京康為世紀生物公司生產,編號為EG101-2的DNA提取試劑盒。
1.1.3試驗地基本情況 試驗地位于吉林省西部公主嶺市(東經124°02′~125°18′,北緯43°11′~44°09)吉林省農科院畜牧分院院內。該地土壤為退化黑鈣土,肥力中等,該地主要有龍葵(Solanumnigrum)、蒼耳(Xanthiumsibiricum)、小薊(Cirsiumsetosum)、鐵莧菜(Acalyphaaustralis)等多種雜草。F1代雜交種種于吉林省農科院農業生物研究所的溫室內。
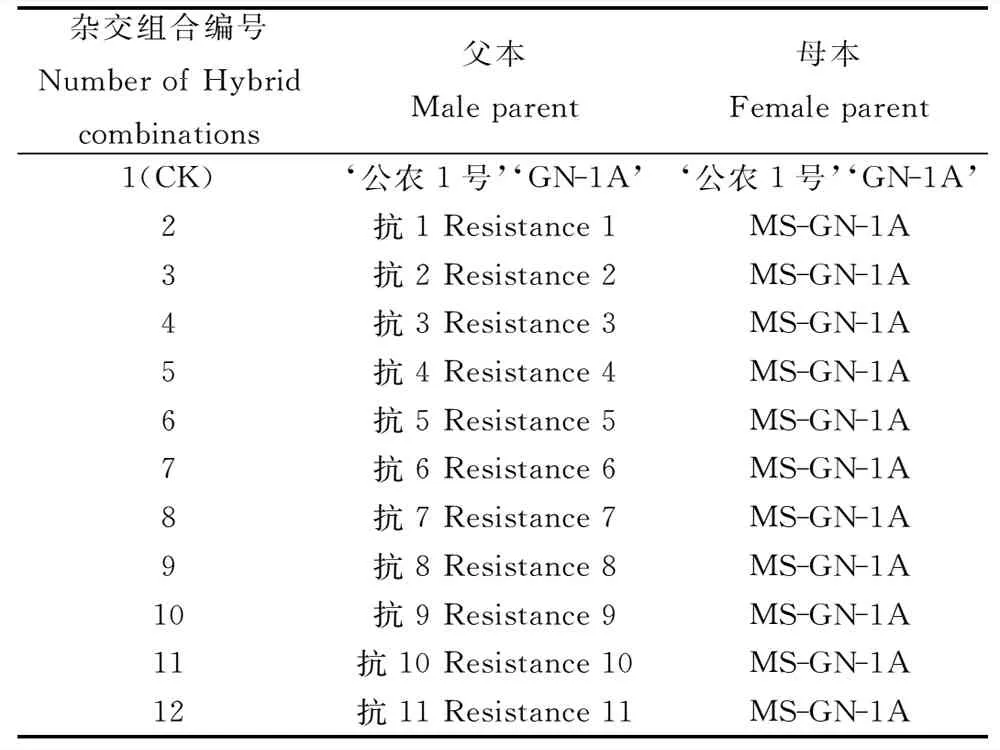
表1 試驗材料
1.2 試驗方法
1.2.1雜交F1代抗除草劑特性的檢測與篩選 在雜交F1代紫花苜蓿進入2~3葉期時用Bar試紙條對雜交后代進行初步篩選,得到Bar試紙條檢測到的陽性紫花苜蓿葉片,然后根據試劑盒提示提取基因組DNA,檢測所提取的DNA質量。以Bar基因序列作為特異引物用PCR儀(型號A37834)進行PCR擴增,擴增程序為94℃ 預變性7 min;94℃ 變性30 s,56℃ 退火30 s,72℃ 延伸30 s,30個循環;72℃ 延伸4 min;4℃ 保存。反應產物在1.5%瓊脂糖凝膠100 V電壓下電泳約2 h,在凝膠成像儀上拍照并保存。Bar基因引物設計為:F:5′ATGAGCCCAGAACGACGCCC3′,R:5′CAGAT CTCGGTGACGGGCAGGAC3′。
1.2.2雜交組合產量和品質性狀的測定 在初花期,對紫花苜蓿各個雜交組合F1代的株高和分枝數進行測量。將各雜交組合的后代全部留茬5 cm進行刈割。每組合雜交F1中隨機挑取10株,進行鮮重的測定,隨后放入溫度為105℃的烘箱殺青15 min,再放入65℃的烘箱烘干至恒重,測定并記錄莖、葉干重,計算莖葉比。
莖葉比(stem/leaf,S/L)=莖干重/葉干重
以索氏提取法(參考GB/T6433-2006)測定粗脂肪;以凱氏定氮法(參考GB/T6432-1994)測定粗蛋白;以酸堿洗滌重量法測定粗纖維(參考GB/T6434-1994)含量。
1.2.3灰色關聯度分析方法 用X表示12個雜交組合,用K表示農藝學性狀,以分枝、株高、干重、鮮重、葉莖比、粗纖維、粗脂肪和粗蛋白等8項性狀中的最大值作為參考數列,記為X0= {X0(1),X0(2),X0(3),……X0(n)};各雜交組合X在各農藝學性狀k處的值構成比較數列Xi,Xi={Xi(1),Xi(2),Xi(3),……Xi(n)},其中i=1,2,3,……N,N指的是組合數目,n為性狀數目,分別為 12 和 8。具體公式如下:
關聯系數:
(1.1)
等權關聯度:
(1.2)
權重系數:
(1.3)
加權關聯度:

(1.4)
其中|X0(k)-Xi(k)|表示絕對差值△i(k),minimink|X0(k)-Xi(k)|最小絕對差,maximaxk|X0(k)-Xi(k)|表示最大絕對差。ρ代表絕對分辨系數,一般在[0,1],本文取ρ=0.5。
1.2.4數據分析 采用Exce1 2003進行基礎數據的記錄與統計,用SPSS 22.0統計分析軟件進行方差分析和相關性分析,其中均值的差異顯著性和多重比較分別采用ANOVA和Duncan法分析,灰色關聯度分析采用灰色關聯度公式與Excel 2003相結合來完成,各參數全部采用均值±標準誤表示。
2 結果與分析
2.1 雜交后代的Bar試紙條檢測
使用Bar試紙條對雜交后代紫花苜蓿中是否有Bar基因進行檢測。經過Bar試紙條快速鑒定得出轉Bar基因抗除草劑的紫花苜蓿,部分檢測結果如圖1所示:編號1,2,5,8有2條帶出現,表明是轉Bar基因抗除草劑紫花苜蓿陽性植株;編號3,4,6,7,9,10有1條帶出現,表明是轉Bar基因抗除草劑紫花苜蓿陰性植株。

圖1 試紙條檢測結果
2.2 雜交后代的PCR檢測
以質粒為陽性對照,非轉基因苜蓿為陰性對照,對Bar試紙條檢測到的陽性苜蓿葉片提取基因組DNA,并以Bar基因序列作為特異引物進行PCR擴增。PCR檢測后,部分檢測結果如圖2所示:在11份F1代材料中都發現了特異性片段,說明配置轉基因紫花苜蓿雜交組合的可行性。

圖2 雜交后代的PCR鑒定圖
2.3 F1代農藝學性狀的分析
將篩選得到的具有草銨膦抗性的雜交后代進行農藝學性狀(產量和品質性狀)分析,如圖3所示,組合1為對照組(CK),株高最高的組合是4,之后依次是組合5、組合12、組合3,株高最低的組合是9,其次是組合6,最高組合比最低組合株高高出23.67 cm。組合3、組合4、組合5、組合7、組合8、組合11和組合12表現較好,株高均高于對照組合,其中組合4顯著高于對照組合(P<0.05)。

圖3 12個苜蓿雜交組合株高性狀的比較
如圖4所示,單株分枝數最多的組合為4,其次是組合3,最低的組合是9,最高組合比最低組合分枝數高出達32個分枝。組合3、組合4、組合5、組合7和組合8表現較好,單株分枝數均高于對照組合,其中組合4顯著高于對照組合(P<0.05)。
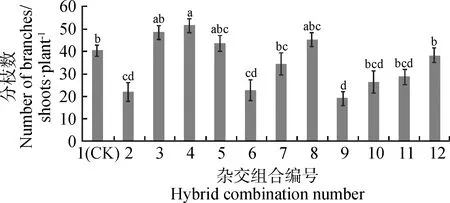
圖4 12個苜蓿雜交組合分枝數的比較
如圖5所示,干草產量最高的是組合4,之后是組合3和組合1,干草產量最低的是組合9,最高組合比最低組合的干重高出0.44 kg。組合3、組合4、組合5、組合7和組合8表現較好,干草產量均高于對照組合,其中組合4顯著高于對照組合(P<0.05)。
如圖6所示,鮮草產量最高的組合是4,其次是組合3、組合5和組合8,鮮草產量最低的組合是9,其次是組合12,最高組合比最低組合的鮮重高出0.68 kg。組合3、組合4、組合5、組合7和組合8表現較好,鮮草產量均高于對照組合,其中組合4顯著高于對照組合(P<0.05)。

圖5 12個苜蓿雜交組合干重的比較

圖6 12個苜蓿雜交組合鮮重的比較
如圖7所示,葉莖比最高的組合是4,最低的是組合2,各組合葉莖比從大到小依次是:組合4>組合5>組合3>組合8>組合12>組合7>組合1>組合11>組合10>組合6>組合9>組合2。組合3、組合4、組合5、組合7、組合8和組合12表現較好,葉莖比均高于對照組合。

圖7 12個苜蓿雜交組合葉莖比的比較
如圖8所示,粗蛋白最高的是組合5,最低的是組合6,各組合粗蛋白百分比從大到小依次是:組合5>組合3>組合4>組合12>組合8>組合7>組合1>組合10>組合11>組合2>組合9>組合6。組合3、組合4、組合5、組合7、組合8和組合12表現較好,粗蛋白百分比均高于對照組合,其中組合3、組合4、組合5、組合8和組合12顯著高于對照組合(P<0.05)。

圖8 12個苜蓿雜交組合粗蛋白的比較
如圖9所示,粗脂肪最高的是組合8,最低的是組合9,各組合粗脂肪百分比從大到小依次是:組合8>組合5>組合4>組合3>組合7>組合12>組合1>組合11>組合2>組合6>組合10>組合9。組合3、組合4、組合5、組合7、組合8和組合12表現較好,粗脂肪百分比均高于對照組合,其中組合8顯著高于對照組合(P<0.05)。
如圖10所示,粗纖維百分比最高的組合是9,最低的是組合8,各組合粗纖維百分比從大到小依次是:組合9>組合10>組合6>組合2>組合11>組合1>組合12>組合7>組合3>組合4>組合5>組合8,粗纖維含量的變化規律與粗蛋白含量相反。組合3、組合4、組合5、組合7、組合8和組合12表現較好,粗纖維百分比均低于對照組合,其中組合3,組合4,組合5,組合8顯著低于對照組合(P<0.05)。

圖9 12個苜蓿雜交組合粗脂肪的比較

圖10 12個苜蓿雜交組合粗纖維百分比的比較
2.4 雜交后代農藝學性狀的灰色關聯度分析
2.4.1原始數據的標準化處理 本試驗包括的8項指標分別為:株高、分枝數、鮮重、干重、粗脂肪、粗蛋白、粗纖維和葉莖比。各雜交組合的等權和加權關聯系數值,如表2,3所示。

表2 各雜交組合的等權關聯系數值
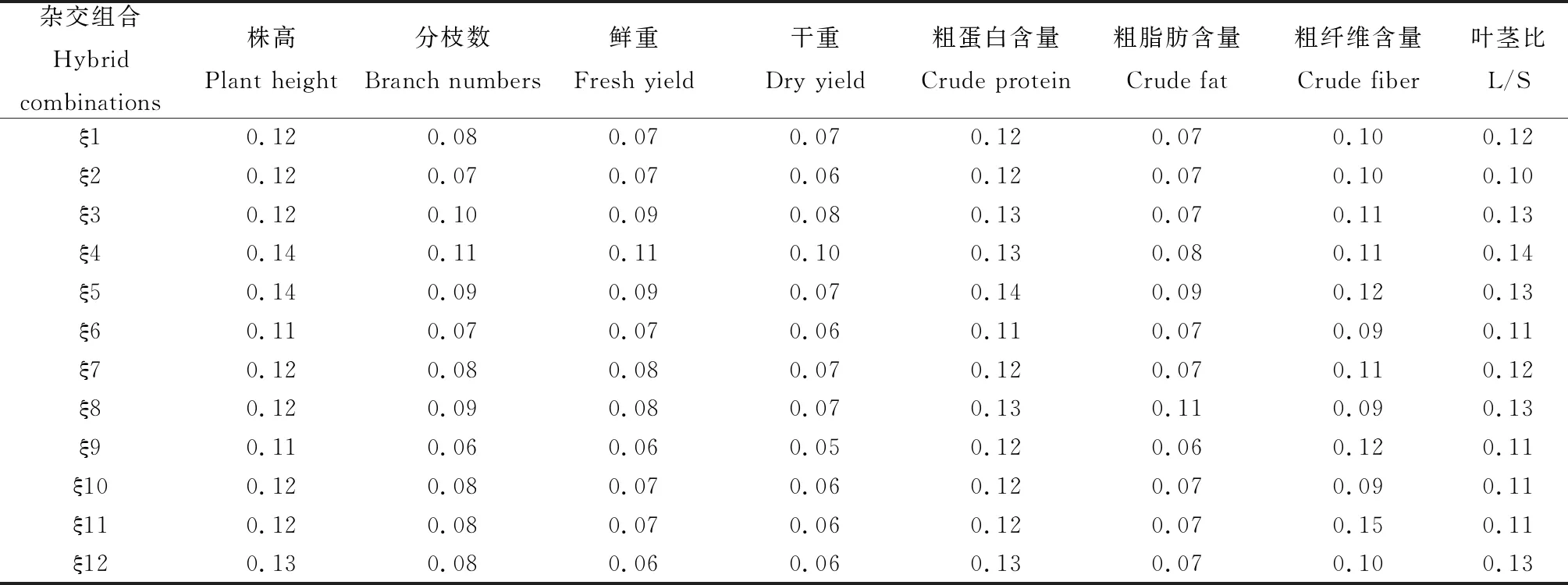
表3 各雜交組合的加權關聯系數值
2.4.2各雜交組合的關聯度分析 由表4可知,加權關聯度和等權關聯度的各項指標的排列順序基本一致,但個別幾項有細小出入,最后對各個指標的等權關聯度結果進行由大到小的順序排列:γ4>γ8>γ5>γ3>γ11>γ12>γ7>γ1>γ10>γ6>γ2>γ9,加權關聯度結果進行由大到小的順序排列:γ4>γ5>γ3>γ8>γ11>γ12>γ7>γ1>γ10>γ2>γ9>γ6,優勢組合除組合3、組合5和組合8,劣勢組合除組合2、組合6和組合9在加權和等權關聯度分析位次中有些許差異,基本上等權關聯度和加權關聯度呈正相關的關系,優勢組合和劣勢組合兩種關聯度法表現基本一致。優勢組合包括組合3,4,5,7,8,11,12,各種性狀綜合表現突出,綜合性狀高于對照組合;劣勢組合包括組合2,6,9,10,各種性狀綜合表現較差,綜合性狀低于對照組合。

表4 各雜交組合的關聯度及排序
3 討論
3.1 紫花苜蓿雜交后代抗性分析
抗除草劑育種是最早開展和應用的轉基因育種,由于紫花苜蓿本身組織培養再生性差,重復性低等原因,從而造成轉化率低、遺傳或表達不穩定等問題。因此,利用轉基因工程技術手段來改良紫花苜蓿的遺傳特性受到很大限制。如何利用好現有的紫花苜蓿轉基因材料,培育出高產、優質且抗除草劑的苜蓿品種是本試驗主要研究的問題。
本試驗利用抗除草劑基因紫花苜蓿和雄性不育系紫花苜蓿進行雜交,使轉基因技術與傳統育種方法有效結合,發揮各自優勢,加快和完善轉基因目標性狀導入紫花苜蓿的進程。國內外研究表明外源基因在受體作物內的遺傳表達很不穩定,易出現基因沉默現象[14-15]。抗除草劑轉基因作物的目的基因遺傳表現復雜,部分轉基因作物自交后代表現3∶1分離規律,與非轉基因親本雜交后代表現1∶1分離規律,但也有一部分除草劑轉基因作物的表現不符合孟德爾分離規律[16]。本試驗以轉Bar基因紫花苜蓿為父本,雄性不育系紫花苜蓿為母本,進行人工雜交。對雜交組合F1代進行抗性檢測,發現均有特異性條帶出現,證明這些后代中已經存在抗除草劑基因。孟慶忠[17]采用農達41%水劑作為殺雄劑與轉EPSPS基因的棉花進行雜交棉制種,棉花雜交種純度不低于96%。本試驗研究結果與其相似,獲得的F1代純度在98%以上。
3.2 紫花苜蓿雜交后代產量及品質性狀的分析
苜蓿雄性不育系的選育及其利用已成為當前苜蓿生產中的熱點,一旦雜種優勢獲得顯著增產優質的效果,就可以大幅度提升該品種的市場競爭力[18]。吳永敷等[19]研究提出利用苜蓿雄性不育是獲得雜種優勢以得到高產優質抗逆的苜蓿品種的有效途徑。Brummer[20]指出雄性不育系的選育是苜蓿雜種優勢利用的基礎,利用雄性不育系和常規品種雜交獲得的F1代能夠有效避免自交衰退。特木爾布和等[21]以MS-4配置40個雜交組合,得到雜交F1代的產量與對照相比平均增產25.2%。本研究結果與其相似,本試驗中雜交組合3、組合4、組合5、組合7、組合8的F1代鮮草產量和干草產量均高于組合1(CK),初步說明這幾個雜交組合得到的后代在產量方面優于常規品種的雜交后代。
粗蛋白含量是劃分紫花苜蓿等級的重要標準依據。一般來說優質的紫花苜蓿干草中粗蛋白含量大于18%[22]。粗脂肪是提供能量的主要物質,其重要性僅次于粗蛋白,其含量影響苜蓿的適口性。本試驗研究發現,雜交組合3、組合4、組合5、組合7、組合8、組合12的F1代中粗蛋白和粗脂肪含量均高于常規品種的雜交后代(CK),且粗蛋白含量均在18%以上,說明本試驗選取的常規材料‘公農1號’紫花苜蓿為優質苜蓿,以上6個雜交組合在品質方面體現出雜種優勢。根據2006年我國頒布的牧草草產品標準[23]指標規定,試驗中雜交組合5為一級(粗蛋白質含量:20%~22%)牧草、雜交組合3、組合4、組合7、組合8、組合12為二級(粗蛋白質含量:18%~20%)牧草。
研究發現,雜交組合2、組合6、組合9、組合10、組合11的F1代的產量與品質均低于對照,未獲得雜種優勢,可能與親本的性狀有關,尤其是轉基因材料,親本的性能比常規品種稍弱一些。
3.3 紫花苜蓿雜交后代產量、品質相關性分析研究
產量是衡量植物生產性能和經濟性能的重要指標[24],因此,高產育種對紫花苜蓿尤為重要。紫花苜蓿產量是受多基因控制的數量性狀,遺傳和環境對產量影響較大,主要由單株株高、分枝數等多個性狀與環境互作的結果[25]。海棠[26]認為產量大小是產量各構成因子平衡的結果,產量的高低取決于各因子間能夠達到最佳平衡狀態。以往對苜蓿產量的研究普遍認為,株高與干草產量、鮮草產量呈顯著或極顯著正相關。本試驗通過11個雜交組合的產量因子分析研究表明,鮮草產量、干草產量均與株高和分枝數呈極顯著正相關,說明株高與分枝數對苜蓿產量優勢的形成非常重要,這與已有的研究[27]結果基本一致。趙松義等[28]認為優良的煙草雜種F1代,不僅產量具有超親表現,農藝性狀與煙葉產量優勢的形成也密切相關,本研究結果與其相似。
3.4 F1代產量和品質的灰色關聯度分析與評價
評價牧草的好壞的因子很多,僅以單一性狀的方差分析作為評價標準在一定程度上有失全面或無所側重[29]。鄧聚龍[30]的灰色關聯分析法中,等權關聯度可反映草種間差異大小,即關聯度越大的因素,相似度越高[31]。加權關聯系數可真實反應出供試材料與最優指標的差異,即關聯度越大,該材料與最優指標越接近,且相關研究證明,加權關聯度更能準確反應供試材料的優劣[32]。
本試驗對12個雜交組合的產量及品質性狀進行灰色關聯度分析,結果表明組合4的等權關聯度與加權關聯度均排名第一,可能與該組合草產量大、粗蛋白和粗脂肪含量高有重要關系。本試驗結果表示,組合3,5,8,11,12的產量與品質性狀指標相對較高,與參考材料有較大的相似度,使綜合排名相對靠前,均在組合1(CK)之上,說明以上的雜交組合后代表現出不同程度的雜種優勢,各指標性狀均優于常規品種。由此可以看出,株高、分枝數、干草產量、鮮草產量、莖葉比對紫花苜蓿綜合生產性能最大。賈瑞等[33]對不同紫花苜蓿雜交組合F1代的生物學特性的研究表示紫花苜蓿的選育過程中應將單株分枝數、干草產量、粗蛋白粗脂肪作為主要考慮因子,本試驗結果與其不完全一致,這可能由于試驗選擇的雜交組合親本品種不同,前者選用的雜交親本多為國外品種,本試驗則是選用國內東北地區的當家品種,兩者親本的遺傳差異較大,另外也與栽培環境等條件不同有關。
3.5 三系配套技術在轉基因紫花苜蓿育種中的應用
農業多樣化對牧草產業的發展有著積極而又深遠的影響[34]。我國在應用生物技術進行牧草遺傳改良方面已經取得了許多成就[35]。王木月等[36]通過回交轉育的手段,把抗除草劑基因Bar導入到多個秈稻(OryzasativaL.)品種中,并選育出不同基因組合的秈型轉基因水稻株系。本試驗以優良紫花苜蓿雄性不育系作為母本、優異的抗除草劑轉基因紫花苜蓿公農1號為父本,配置雜交組合,得到具有抗除草劑特性的雜交組合F1代。
根據已有研究[37-38],轉基因作物的育種方法主要是集中在以分子標記為輔助的回交育種法,但是轉基因作物與現有且相應的雄性不育系進行雜交育種的研究鮮有報道,在抗除草劑轉基因紫花苜蓿中尚未見相關研究。使用雄性不育系來組配雜交種,可免去人工去雄的繁瑣問題,簡化了雜交育種的進程,提高了雜交種種子產量和制種效率,并使雜交種的產量和質量得到保證[39]。本試驗將轉基因技術的研究和應用與現有的紫花苜蓿苜蓿育種技術結合,通過雜交的方式,抗除草劑紫花苜蓿的轉基因特性可以穩定遺傳到F1代。這樣不僅可以縮短育種周期,節省資源,而且目標性狀也能得到穩定統一的遺傳,抗除草劑轉基因苜蓿也可以更加高效穩定的參與到現代的農業生產方式中。在此基礎上,本試驗針對抗除草劑轉基因紫花苜蓿F1代的產量和品質性狀,采用灰色關聯度的方法進行評價和篩選,試驗結果得到7組優秀的雜交組合。
4 結論
本試驗通過抗除草劑紫花苜蓿與非轉基因雄性不育紫花苜蓿雜交后代分析研究,發現抗除草劑Bar基因可穩定遺傳給后代,說明Bar可以通過雜交轉育的方式轉入其他非轉基因紫花苜蓿中。
相關性分析結果表明,紫花苜蓿鮮草產量、干草產量均與株高和分枝數呈極顯著正相關。灰色關聯度分析表明,株高、分枝數、干草產量、鮮草產量、莖葉比對紫花苜蓿綜合生產性能最大。
通過綜合性能指標評定,組合4為最佳雜交組合,其次,組合3,5,7,8,11,12也表現出不同程度的雜種優勢,均可作為配置雜交組合儲備的材料。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
今日農業(2021年14期)2021-11-25 23:57:29
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
中國果業信息(2019年10期)2019-11-13 01:21:34
今日農業(2019年15期)2019-01-03 12:11:33
聚氯乙烯(2018年9期)2018-02-18 01:11:34
現代園藝(2017年19期)2018-01-19 02:50:21
現代農業(2016年5期)2016-02-28 18:42:36
營銷界(2015年23期)2015-02-28 22:06:18
