益氣解毒方藥干預致病菌入侵后腸上皮細胞自噬基因miR130a/ATG16L1的表達
2019-11-14 00:40:00樊冬梅索娜劉潔明任寶琦
廣州中醫藥大學學報 2019年12期
關鍵詞:血清
樊冬梅, 索娜, 劉潔明, 任寶琦
(1.廣州中醫藥大學第一附屬醫院,廣東廣州510405;2.廣州市番禺區何賢紀念醫院消化科,廣東廣州510006;3.廣州中醫藥大學2018級碩士研究生,廣東廣州510006;4.廣東省中醫院,廣東廣州510021)
炎癥性腸病(inflammatory bowel disease,IBD)是原因不明的慢性腸道炎癥性疾病,包括潰瘍性結腸炎(ulcerative colitis,UC)和克羅恩病(Crohn’s disease,CD)。IBD病程遷延,反復發作,嚴重影響患者的生活質量,被世界衛生組織列為現代難治病之一[1,2]。近年來,某些致病菌尤其是侵襲性大腸桿菌(adherent-invasive E.coli,AIEC)的入侵導致的腸上皮細胞(EICs)自噬功能障礙致胞內細菌清除不足,進而引起啟動炎癥反應通路激活,是腸道黏膜受損、炎癥反復發作的關鍵機制[3-7]。研究表明Atg16L1、Atg5等與IBD的發病密切相關,是控制自噬介導的炎癥反應的重要自噬基因,其介導的腸上皮細胞自噬功能異常是IBD腸道炎癥啟動及復燃的關鍵[7,8]。感染AIEC后IECs通過上調miR-30c/130a表達、下調自噬基因Atg5、Atg16L1的表達引起細胞自噬功能下降而致胞內致病菌清除不足[3-6]。因此,如何能維持適度的EICs自噬功能從而有效清除IBD相關致病菌對于IBD的預防、治療和轉歸極為重要。目前國內許多學者主張益氣解毒法用于防治IBD的復發[9,10]。廣東省名中醫勞紹賢教授就是將本法成功用于IBD防治,臨床療效確切[11-13]。我們前期研究證實以此法為主的復方干預治療IBD可有效預防復發[11],腸炎靈片作為醫院的院內制劑應用于防治IBD的臨床中取得了較好的療效,有著廣闊的應用前景。本研究進一步從干預致病菌入侵后腸上皮細胞自噬基因miR130a/ATG16L1表達的角度,探討益氣解毒方藥防治IBD的新途徑,為其進一步的臨床推廣提供理論依據,現將研究結果報道如下。
1 材料與方法
1.1 細胞與細菌HCT116細胞株,來自中國科學院細胞庫。福氏志賀菌GIM1.539標準菌株購自廣州微生物研究所。
1.2 動物SPF級SD大鼠及SPF級鼠料,均購自廣東省實驗動物中心,動物質量合格證號:0602237。
1.3 藥物及制備 益氣解毒方組成:黨參15 g,白術15 g,茯苓15 g,黃連10 g,救必應30 g,白花蛇舌草30 g,烏藥30 g,炙甘草5 g。上述藥材飲片由廣州中醫藥大學第一附屬醫院實驗中心藥理研究室鑒定并制成3 g·mL-1的中藥湯劑。
1.4 試劑與儀器DMEM高糖、雙抗、體積分數10%胎牛血清、胰酶(上海立菲生物技術有限公司);ATG16L1抗體、二抗(美國Sigma公司);總蛋白提取試劑盒(廣州市杏海生物科技有限公司);Trizol、PCRMix Taq、溴乙錠(北京艾德萊公司);miR130a模擬物及反義模擬物(美國Sigma公司);熒光素酶檢測試劑盒(美國Promega公司);瓊脂糖、PrimeScriptTMRT reagent Kit(Perfect Real Time)、SYBR?Premix Ex TaqTMII(Tli RNaseH Plus)、RNAiso Plus(日本TAKARA公司);GAPDH引物序列上游5’-GAAGGTGAAGGTCGGAGTC-3’,下游5’-GAAGATGGTGATGGGATTTC-3’;ATG16L1上游5’-GAGCTGCTCCCGTGATGACT-3’,下游5’-CGCCACGTAACTGCCATCAG-3’;miR130a上游5’-AGGCCGCAGTGCAATGTT-3’,下游5’-GTGCAG GGTCCGAGGT-3’;引物合成,于上海生工生物工程有限公司完成。3K30型臺式高速冷凍離心機(美國Sigma公司);D1008低速離心機梯度(美國Scilogex公司);D-8721PCR儀(杭州晶格公司);NanoDrop2000超微量核酸蛋白檢測儀(美國Thermo Scientific公司);CFX96熒光定量PCR儀、imark酶標儀、2211BR Trans-Blot(SD Cell轉印系統、ChemiDocTMXRS+成像儀(美國Bio-Rad公司);TS-1脫色搖床(海門市麒麟醫用儀器廠)。
1.5 實驗方法
1.5.1 HCT116細胞株的培養和傳代 以1∶9比例(體積分數)配制的含胎牛血清的高糖DMEM培養基作為完全培養基,培養基中加入雙抗(青霉素100 U/mL,鏈霉素100 U/mL)。獲得細胞后,棄上清液,加入磷酸鹽緩沖液(PBS)沖洗后棄上清液。加入2.5 g/L胰酶消化3min,輕輕吹打細胞致完全脫落,立即加入含血清的培養液形成細胞懸液。1 000 r/min離心5min后棄上清液,加入新鮮完全培養基輕輕吹打混勻細胞,后轉移至培養皿,做好標記。于37℃、體積分數5%CO2的培養箱中培養細胞2 d。待HCT116細胞貼壁生長至80%以上棄掉培養基進行傳代,根據細胞狀態可1傳2。
1.5.2 中藥含藥血清制備 按照3 g·mL-1生藥濃度制取中藥湯劑原液,置于4℃冰箱保存備用。大鼠飼養1周后開始灌胃,每只大鼠每天早9點、晚4點各灌胃1次,每次1.5mL,對照組灌服等量生理鹽水。自由進食水,及時補充鼠料。連續給藥7 d,第7天灌藥前禁食12 h。末次給藥2 h后,無菌環境下乙醚麻醉大鼠,采用腹主動脈采血法收集大鼠全血,靜置30min,血液以3 000 r/min(離心半徑13.5 cm)離心15min,用0.22μm微孔濾膜無菌分離血清,56℃水浴,30min滅活,-20℃冰箱保存備用。
1.5.3 miR130a轉染及沉默轉染 使用含體積分數10%胎牛血清的DMEM高糖培養基培養細胞,將細胞鋪在24孔板中,使用脂質體2000將熒光素酶報告載體同50 nmol/LmiR130a模擬物或相應的陰性對照物共轉染至細胞中。轉染6 h后,按照熒光強度與陰性對照組對照判斷轉染是否有效。以上述步驟中合成的目的片段為目的基因的反義基因作為miR130a反義模擬物進行沉默轉染。轉染都使用脂質體2000,含體積分數10%胎牛血清的DMEM培養基培養細胞,將細胞鋪在24孔板中,使用脂質體2000將熒光素酶報告載體同50 nmol/L miR130a反義模擬物或相應的陰性對照物共轉染至細胞中。轉染6 h后,按照熒光強度與陰性對照組對照判斷轉染是否有效。
1.5.4 福氏志賀菌GIM1.539標準菌株感染HCT116細胞模型的建立 福氏志賀菌GIM1.539標準菌株液體培養至對數生長期,離心收集細菌,用無抗生素DMEM高糖培養基制備細菌懸液,通過測定吸光度[D(600 nm)]值調整細菌濃度[1D(600 nm)=1×108CFU/mL]。以含體積分數10%小牛血清的DMEM高糖(青霉素100 U/mL,鏈霉素100 U/mL)為完全培養基,培養HCT116細胞株培養2 d。待HCT116細胞貼壁生長至80%時,棄掉培養基。按照感染復數(multiplicity of infection,MOI,MOI=細菌數∶細胞數)為100∶1加入GIM1.539懸液,37℃、體積分數5%CO2的條件下共培養。入侵分析方法為在沒有抗生素的培養基孵化3 h,PBS沖洗細胞,含有100μg/mL慶大霉素在指定時間被加入培養基中,細胞被用1%Triton X-100溶解在脫離子水中,樣品持續稀釋并固定在瓊膠上,細菌的數量通過計數菌落形成單位(CFU)來確定。
1.5.5 蛋白免疫印跡(Western Blot)法檢測HCT116細胞中ATG16L1、LC3的蛋白表達 將細胞及培養液移至離心管中,1 000 r/min離心10min,再用預冷PBS,1 000 r/min離心5min洗2次去除培養液。在每毫升冷裂解緩沖液中加入5μL磷酸酶抑制劑,1μL蛋白酶抑制劑和5μL PMSF(濃度200 mmol/L,溶于異丙醇),混勻。冰上保存數分鐘待用。在離心管的細胞中加入上述配制好的冷裂解緩沖液,5×106個細胞,加入量約為0.4mL,冰上操作,細胞與冷裂解緩沖液一同轉移至新的預冷的離心管中,置于4℃搖床平臺上;溫和振蕩15min,14 000 r/min,4℃離心15min,取上清為全蛋白提取物,蛋白定量后,分裝保存于-70℃,避免反復凍融。按照試劑盒說明繪制標準曲線測定蛋白質濃度。經十二烷基硫酸鈉—聚丙烯酰胺凝膠電泳(SDS-PAGE)分離、轉膜,20 g/L白蛋白4℃封閉過夜后,依次加入一抗,4℃過夜,棄去一抗加入相對應的二抗,ECL顯色,曝光、顯影、定影、拍照、掃描后通過BandScan軟件進行條帶灰度半定量分析,以GAPDH為內參。
1.5.6 熒光定量聚合酶聯反應(PCR)法檢測HCT116細胞中miR130a、ATG16L1的RNA表達 以6×105/孔接種至6孔板中的細胞處理6 h后,以PBS漂洗細胞3次。每孔加入1mL Trizol,按照Trizol說明書提取總RNA。從-80℃冰箱中取出樣品RNA,于4℃解凍,然后在0.2mL PCR管中配制反應溶液,37℃、42℃15min(反轉錄反應)進行cDNA的合成,SYBRGreen PCR,將PCR管置于定量PCR儀中進行反應。
1.5.7 實驗分組 體外GIM1.539感染HCT116細胞模型建立后,根據添加的血清不同、干擾方案的不同分組,每次實驗每組設6個復孔。具體分組見表1。

表1 細胞實驗分組Table 1 The grouping in the cell test
1.6 統計方法 使用SPSS 19.0統計軟件,計量資料以均數±標準差()表示,首先對每組數據進行正態分布和方差齊性分析。樣本例數較小(n=6),對于組內差異比較,若服從正態分布且方差齊性,則選用單因素方差分析;對于組間差異比較,若服從正態分布且方差齊性,采用獨立樣本t檢驗,否則選用秩和檢驗。檢驗結果以P<0.05表示差異具有統計學意義。
2 結果
2.1 Western Blot法檢測HCT116細胞中ATG16L1、LC3的蛋白含量 各組細胞轉染成功并接受干預完成后,用Western Blot法檢測各組細胞內的ATG16L1、LC3蛋白含量水平,電泳結果見圖1、2,其灰度值結果見表2、表3。結果顯示:含藥血清干預方式與無藥血清干預方式miR130a模擬物組ATG16L1、LC3蛋白含量均較空白轉染組明顯減少(P<0.05);含藥血清干預方式與無藥血清干預方式miR130a反義模擬物組ATG16L1、LC3蛋白含量較空白轉染組均明顯增多(P<0.05);含藥血清干預方式miR130a模擬物組、反義模擬物組和空白轉染組的ATG16L1、LC3蛋白含量均較同組無藥血清干預方式的含量多(P<0.05)。除此之外,無論何種干預方式,miR130a模擬物或反義模擬物的陰性對照物組的ATG16L1、LC3蛋白含量均與空白轉染組無明顯差異。
2.2 熒光定量PCR法檢測HCT116細胞中miR130a、ATG16L1的基因表達2組細胞轉染成功并接受干預完成后,用熒光定量PCR法檢測各組細胞中的miR130a、ATG16L1的基因表達水平,結果見表4、5。結果顯示:含藥血清與無藥血清干預方式miR130a模擬物組分別與本組空白轉染組比較,miR130a模擬物組miR130a的基因表達明顯上升(P<0.05),而ATG16L1的基因表達明顯下降(P<0.05);含藥血清與無藥血清干預方式miR130a反義模擬物分別與本組空白轉染組比較,miR130a反義模擬物組miR130a的基因表達明顯下降(P<0.05),ATG16L1的基因表達明顯上升(P<0.05);而含藥血清干預方式miR130a模擬物組、反義模擬物組和空白轉染組的miR130a基因表達均較同組無藥血清干預方式的下降(P<0.05),而ATG16L1基因表達均較同組無藥血清干預方式的上升(P<0.05)。
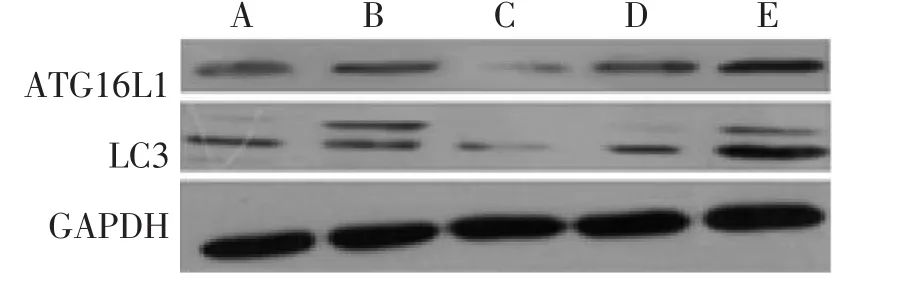
圖1 含藥血清干預的HCT116細胞中ATG16L1、LC3蛋白表達Figure 1 The protein expression of ATG16L1 and LC3 in HCT116 cells intervened by drug-containing serum
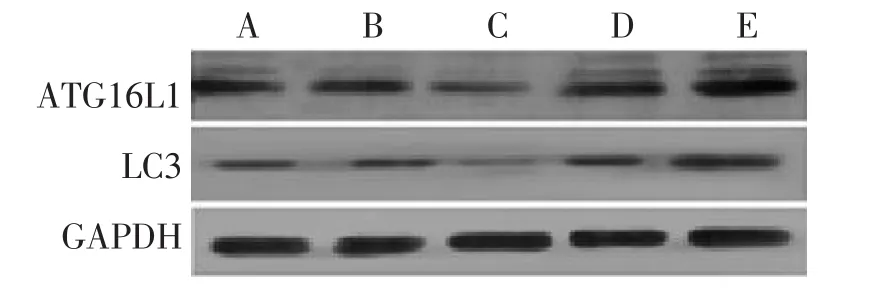
圖2 無藥血清干預的HCT116細胞中ATG16L1、LC3蛋白表達Figure 2 The protein expression of ATG16L1 and LC3 in HCT116 cells intervened by drug-free serum
表2 2種不同干預方式下各組HCT116細胞ATG16L1蛋白含量比較Table 2 Comparison of the protein content of ATG16L1 in HCT116 cells of various groups after2 types of intervention(,n=6)
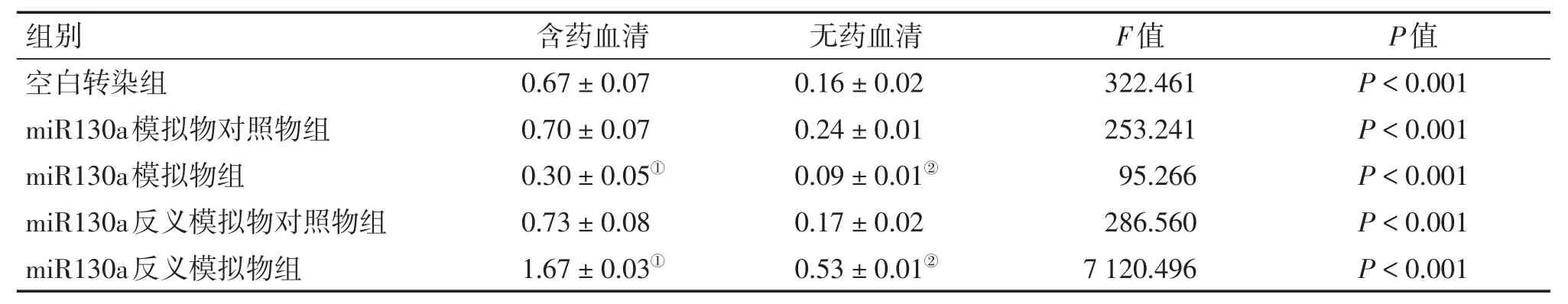
表2 2種不同干預方式下各組HCT116細胞ATG16L1蛋白含量比較Table 2 Comparison of the protein content of ATG16L1 in HCT116 cells of various groups after2 types of intervention(,n=6)
①P<0.05,含藥血清干預方式中與空白轉染組比較;②P<0.05,無藥血清干預方式中與空白轉染組比較
組別 含藥血清 無藥血清F值P值空白轉染組miR130a模擬物對照物組miR130a模擬物組miR130a反義模擬物對照物組miR130a反義模擬物組0.67±0.07 0.70±0.07 0.30±0.05①0.73±0.08 1.67±0.03①0.16±0.02 0.24±0.01 0.09±0.01②0.17±0.02 0.53±0.01②322.461 253.241 95.266 286.560 7 120.496 P<0.001 P<0.001 P<0.001 P<0.001 P<0.001
表3 2種干預方式下各組HCT116細胞LC3蛋白含量比較Table 3 Com parison of the protein contentof LC3 in HCT116 cells ofvarious groups after2 types of intervention(,n=6)
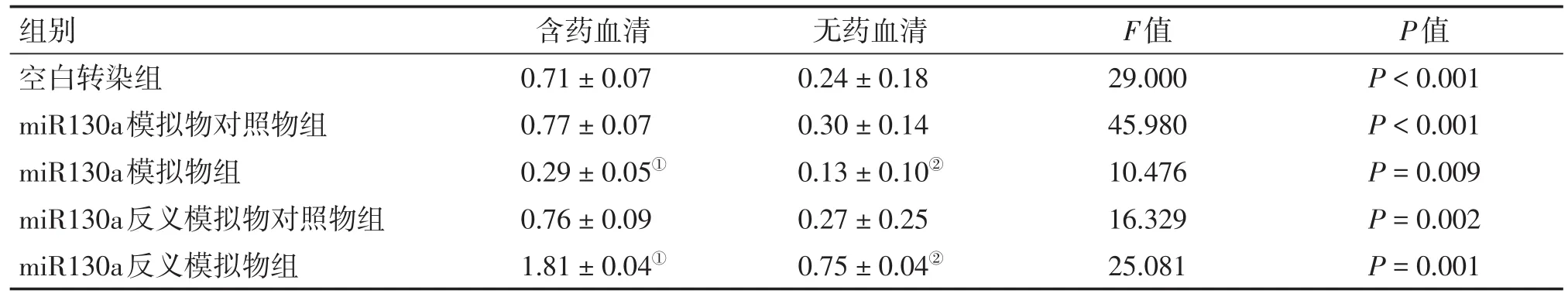
表3 2種干預方式下各組HCT116細胞LC3蛋白含量比較Table 3 Com parison of the protein contentof LC3 in HCT116 cells ofvarious groups after2 types of intervention(,n=6)
①P<0.05,含藥血清干預方式中與空白轉染組比較;②P<0.05,無藥血清干預方式中與空白轉染組比較
組別空白轉染組miR130a模擬物對照物組miR130a模擬物組miR130a反義模擬物對照物組miR130a反義模擬物組含藥血清0.71±0.07 0.77±0.07 0.29±0.05①0.76±0.09 1.81±0.04①無藥血清0.24±0.18 0.30±0.14 0.13±0.10②0.27±0.25 0.75±0.04②F值29.000 45.980 10.476 16.329 25.081 P值P<0.001 P<0.001 P=0.009 P=0.002 P=0.001
表4 2種干預方式下各組HCT116細胞m iR130a的基因表達比較Table 4 Com parison of the gene expression ofm iR130a in HCT116 cells ofvarious groups after2 types of intervention(,n=6)
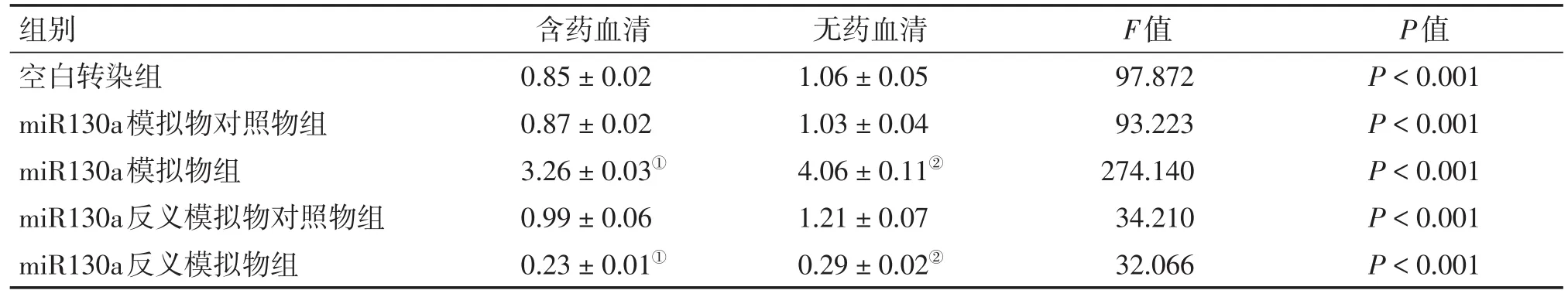
表4 2種干預方式下各組HCT116細胞m iR130a的基因表達比較Table 4 Com parison of the gene expression ofm iR130a in HCT116 cells ofvarious groups after2 types of intervention(,n=6)
①P<0.05,含藥血清干預方式中與空白轉染組比較;②P<0.05,無藥血清干預方式中與空白轉染組比較
組別空白轉染組miR130a模擬物對照物組miR130a模擬物組miR130a反義模擬物對照物組miR130a反義模擬物組含藥血清0.85±0.02 0.87±0.02 3.26±0.03①0.99±0.06 0.23±0.01①無藥血清1.06±0.05 1.03±0.04 4.06±0.11②1.21±0.07 0.29±0.02②F值97.872 93.223 274.140 34.210 32.066 P值P<0.001 P<0.001 P<0.001 P<0.001 P<0.001
表5 2種干預方式下各組HCT116細胞ATG16L1的基因表達比較Table 5 Com parison of the gene expression of ATG16L1 in HCT116 cells ofvarious groups after 2 types of intervention(,n=6)
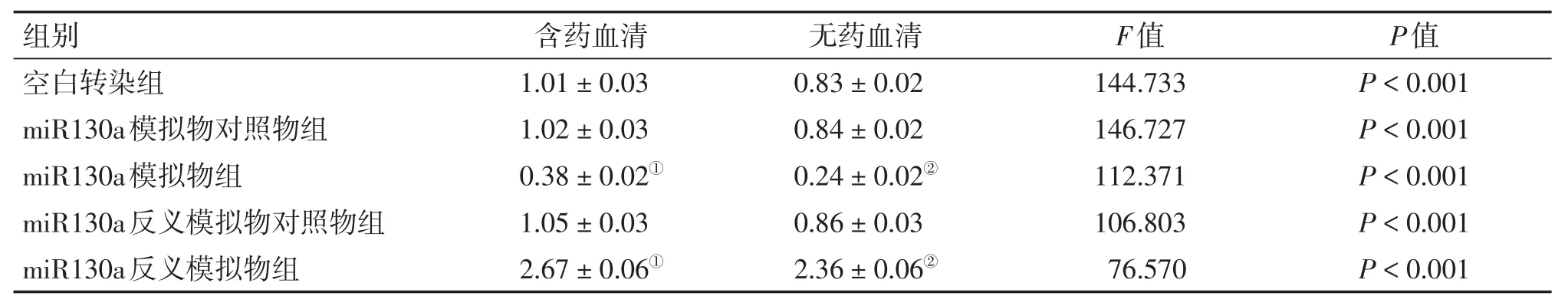
表5 2種干預方式下各組HCT116細胞ATG16L1的基因表達比較Table 5 Com parison of the gene expression of ATG16L1 in HCT116 cells ofvarious groups after 2 types of intervention(,n=6)
①P<0.05,含藥血清干預方式中與空白轉染組比較;②P<0.05,無藥血清干預方式中與空白轉染組比較
組別 含藥血清 無藥血清F值P值空白轉染組miR130a模擬物對照物組miR130a模擬物組miR130a反義模擬物對照物組miR130a反義模擬物組1.01±0.03 1.02±0.03 0.38±0.02①1.05±0.03 2.67±0.06①0.83±0.02 0.84±0.02 0.24±0.02②0.86±0.03 2.36±0.06②144.733 146.727 112.371 106.803 76.570 P<0.001 P<0.001 P<0.001 P<0.001 P<0.001
3 討論
某些致病菌尤其是AIEC入侵導致的腸道穩態及免疫系統平衡被打破是腸道黏膜受損、炎癥反復發作的關鍵機制[2]。充分的證據顯示IBD患者腸道微生態紊亂,表現為潛在的有益菌如雙歧桿菌、乳酸桿菌、厚壁菌數量的下降以及假定的致病菌如擬桿菌屬、大腸埃希桿菌數量的上升,并證實了在IBD患者回腸末端黏膜附著大量AIEC,能接近及入侵人IECs,導致IECs自噬能力下降,細菌復制,促使炎癥激活,不僅在IBD急性期,也是緩解期導致炎癥復發的主要因素。因此,IBD致病菌感染相關性細胞自噬功能障礙與IBD發病密切相關[3-6]。志賀菌屬又稱痢疾桿菌,是重要的侵襲性大腸桿菌之一[14]。目前國際上有采用志賀菌作為入侵菌進行入侵IECs探討IBD發生機制的研究[5,6,8],因此,本研究采用志賀菌作為入侵菌進行研究。
自噬是真核生物普遍存在的自穩機制,在胚胎發育、細胞自我保護和生存等過程中發揮關鍵作用。自噬參與胞內微生物感染具有雙重作用:一方面,自噬能夠降解入侵的微生物,即以異源吞噬的方式清除胞內的病原體;另一方面,有些微生物如IBD某些致病微生物能夠抑制細胞自噬而利于自身存活導致炎癥反應。人類全基因組關聯研究揭示,ATG和其他已知影響自噬過程的基因中的微小單核苷酸多態性位點,與克隆氏病的易感性有相關性[7]。ATG16L1在自噬小體形成中發揮關鍵作用,遺傳刪除ATG16L1能損害自噬小體的形成以及蛋白通過自噬途徑的清除,而且還導致TLR激動劑誘導的巨噬細胞促炎因子分泌的增強[15],葡聚糖硫酸鈉誘導的結腸炎加重[16],ATG16L1基因表達在EICs和潘氏細胞,其基因突變誘導亦導致不能有效清除腸道細菌感染,引起腸黏膜炎癥啟動[17,18]。LC3是酵母自噬相關蛋白ATG8的哺乳動物同源基因,LC3翻譯后經修飾形成主要位于胞漿中的LC3-Ⅰ,自噬發生時,LC3-Ⅰ與磷脂酰乙醇胺(PE)結合,定位于自噬體膜上,即LC3-Ⅱ。它在自噬小體的延伸和閉合過程中均發揮重要功能,其作為自噬體膜的重要組分,對于自噬的產生和檢測起著不可替代的作用。對于LC3調控的研究主要集中在轉錄翻譯后的修飾水平,有研究發現IBD致病菌所致IECs自噬障礙通路可檢測到LC3的表達調控異常[4]。因此,也作為本研究評價細胞自噬功能指標之一。目前細胞自噬對IBD炎癥啟動的潛在作用已經越來越明朗,細胞自噬與抗感染免疫密切相關,病原體導致的細胞自噬障礙是誘發炎癥反應和炎癥性疾病的一個重要因素。新的研究已經證明IBD相關病原體介導的自噬缺陷、病原體清除障礙和IBD發病機制之間存在相關性[4,5]。ATG介導的細胞自噬可清除降解細胞內受損傷的細胞結構以及細胞內細菌感染等,是重要的自噬蛋白。許多研究表明致病菌感染相關IBD自噬異常可能與自噬蛋白調控缺陷有關,但其深入的機制可能與調控自噬的相關miRNAs有關[4,5,19]。最新研究[4,5,19]顯示,IBD相關致病菌AIEC感染后可通過上調miR130a表達下調自噬基因ATG16L1的表達,引起細胞自噬功能下降。靶基因預測發現ATG16L1基因存在miR130a的靶點。即miR130a可與靶ATG16L1基因的3’端非翻譯區結合,引起其降解或抑制其蛋白翻譯,推測miR130a可能通過該機制下調ATG16L1基因表達從而影響細胞自噬。
本研究通過對福氏志賀菌GIM1.539入侵HCT116人結腸癌上皮細胞模型進行miR130a模擬物、miR130a反義模擬物轉染,即miR130a過表達和沉默轉染,檢測細胞ATG16L1的蛋白含量以及基因表達,并與空白轉染組對照,同時分別對應進行miR130a模擬物陰性對照物和miR130a反義模擬物陰性對照物轉染作為參照,證實自噬基因ATG16L1表達受miR130a調控。當細胞內miR130a模擬物含量增多時,抑制ATG16L1基因表達,生產該蛋白的量減少,稱為過表達;而當細胞內miR130a反義模擬物增多時,競爭性地與胞內miR130a結合,從而使ATG16L1得到更多表達,稱為沉默。同樣原理,通過同時檢測細胞內LC3自噬蛋白含量,結果顯示miR130a過表達時,LC3蛋白含量亦隨之減少;miR130a沉默時,LC3蛋白含量隨之增加。雖然LC3蛋白調控機制在本課題中未作深入研究,但是當miR130a過表達時,ATG16L1、LC3含量均減少,足以證實自噬小體的形成必然受阻,從而表明miRNAs/ATG網絡調控是影響細胞自噬的重要機制。
IBD病因復雜,被世界衛生組織列為難治性疾病。2006年中醫消化病診療指南將本病劃歸中醫“痢疾”“腸澼”“腹痛”“便血”范疇。多數醫家[9,10]認為,IBD是在脾胃虛弱的基礎上感受寒、熱、濕毒之邪,傷于飲食勞倦或情志,致熱、濕、痰等邪客于腸道,阻滯氣血,腸絡失和,脂膜受損而發病。脾胃虛弱、運化失常是IBD發病過程中的內在共性。鑒于此,國內許多學者提出氣虛毒邪內伏是IBD病情反復及復發的病機關鍵,并指出氣虛尤其脾氣虛是毒邪內伏難以去除的主要原因,而毒邪多由風寒暑濕燥火侵犯人體后,深伏其中,纏綿不去,導致毒邪稽留,暗耗正氣,每于正氣虛弱或其他誘因觸發,屢發屢重,纏綿難愈,因此在治療上主張“益氣解毒為主”是IBD的防治大法[9,10]。中醫毒邪類似于致病菌,但IBD致病菌入侵后單純用清熱解毒方法治療效果卻不明顯,因此類致病菌可導致細胞自噬功能障礙,細菌清除不足,毒邪反復侵襲留存于體內促使炎癥啟動及復發。那么,益氣解毒防治IBD復發的有效機制可能是干預細胞自噬障礙維持適度的細胞自噬功能促使毒邪(致病菌)外出,與中醫益氣促使毒邪外出理論不謀而合。自噬的自我調控功能與中醫“氣”相通。自噬的自我調控是有一定條件的,當細胞受到不良環境的刺激,通過激發一系列信號因子(目前科學界認為與miRNAs/Atg調控網絡有關)對自噬進行調控,這與中醫氣的推動和調控功能相似。中醫認為氣虛則動力不足、功能低下,氣盛則易生邪毒,而氣具有自我調節功能,若氣的調控作用正常,自噬既無太過,也無不及,細胞內部各種功能活動則取得協調平衡,內環境得到穩定[20]。當正氣不足,氣的防御功能異常,細胞抗邪能力下降,自噬不足,外邪(致病菌)首先攻破人體免疫的第一道防線(入侵腸上皮細胞),繼而因正氣虛弱無法有效激活自噬網絡調控,自噬相關基因低表達,無法清除胞內菌和有害物質,為炎癥的進一步發展提供了機會。故認為自噬障礙是中醫氣虛的表現。因此,推測益氣解毒法防治IBD復發的有效機制可能是通過干預細胞自噬障礙以扶正,助腸EICs促使毒邪(致病菌)清除而發揮作用。基于此,構建AIEC感染EICs體外模型,檢測益氣解毒方藥干預后胞內自噬基因表達,從而探討miRNA/ATG調控通路,一方面可為闡明IBD通過自噬障礙引起炎癥復發的病理機制以及益氣解毒方藥干預途徑,另一方面可為揭示IBD中醫氣虛毒邪內伏理論的科學內涵提供新思路。
本院勞紹賢教授最早提出氣虛毒邪內伏是IBD反復發作的病理機制,并以益氣解毒法為主制成了院內制劑腸炎靈片在臨床推廣應用40余年,取得了滿意療效。益氣解毒方(腸炎靈片)主要由黨參、白術、救必應等藥物組成,其中:救必應為冬青科植物鐵冬青的干燥樹皮或根皮,味苦、性寒,異名為白木香,始載于《嶺南采藥錄》,稱“白木香,味苦,清熱毒”,為我國南方地區民間慣用草藥,具有清熱解毒、消腫止痛、利濕、祛風解毒等功效;黨參溫補中焦、益氣補虛,具有健脾功效。在此基礎上加入黃連、白花蛇舌草、烏藥、木香,黃連配以白花蛇舌草擅清胃腸濕熱,加強清熱燥濕、澀腸止痢、瀉火解毒的功效;木香、烏藥辛散溫通,使方中有升有降,亦可健脾益氣,行氣止痛。本研究通過對體外福氏志賀菌GIM1.539感染HCT116細胞模型進行miR130a過表達和沉默轉染后,分別予以大鼠益氣解毒方含藥血清和大鼠無藥血清進行干預,檢測細胞LC3的蛋白含量、ATG16L1的蛋白含量和基因表達以及miR130a的表達進行對照。結果顯示,含藥血清干預方式miR130a模擬物組、反義模擬物組和空白轉染組的LC3蛋白、ATG16L1蛋白和基因表達均較同組無藥血清干預方式的表達增多。含藥血清干預方式miR130a模擬物組、反義模擬物組和空白轉染組的miR130a基因表達均較同組無藥血清干預方式下降。表明益氣解毒方藥可能是通過干預miR130a的基因表達使ATG16L1蛋白得到調控。研究結果同時證實了LC3自噬蛋白的含量有所提升。
綜上所述,益氣解毒方藥可能是通過抑制miRNAs的基因表達、上調ATG自噬基因的表達來維持腸上皮細胞自噬功能,從而有效清除胞內菌控制IBD炎癥復發。但仍需進一步從自噬小體觀察、胞內菌清除、炎癥因子、體內實驗等方面證實。
猜你喜歡
中老年保健(2021年3期)2021-08-22 06:50:04
天津醫科大學學報(2021年2期)2021-03-29 05:31:08
昆明醫科大學學報(2021年1期)2021-02-07 01:06:36
現代臨床醫學(2021年1期)2021-01-26 00:56:02
昆明醫科大學學報(2020年12期)2021-01-26 00:44:04
中華養生保健(2020年4期)2020-11-16 01:31:40
中西醫結合肝病雜志(2020年2期)2020-10-27 02:18:50
豬業科學(2018年8期)2018-09-28 01:27:38
中成藥(2017年8期)2017-11-22 03:18:47
川北醫學院學報(2015年5期)2015-12-05 08:22:29
