土壤中鎘離子對紅蕓豆幼苗葉綠素熒光的影響
2019-11-21 01:32:58趙麗娟王創云
山西農業科學 2019年11期
關鍵詞:質量
張 洪,趙麗娟,王創云
(1.山西省分析科學研究院,山西太原030006;2.忻州師范學院生物系,山西忻州034000;3.山西省農業科學院作物科學研究所,山西太原030031)
伴隨著城鎮化的持續進行,城市產生的污水逐日增多,將污水處理后用于農業灌溉是解決水資源短缺的一條可行途徑。我國的污水灌溉從20 世紀50 年代開始推廣,主要經歷了起步、發展、快速發展3 個階段[1]。在實踐過程中發現,來自冶煉、電鍍、采礦產生的污水灌溉極易導致土壤中鎘含量超標,土壤中的鎘離子對植物具有潛在的危害作用,當鎘離子濃度超出一定范圍時,除影響植物的正常生長外,還會導致植物死亡[2]。
紅蕓豆屬豆科菜豆屬,適宜在晉西北干旱溫熱丘陵山區或溫潤冷涼土石山區種植,目前在山西忻州地區神池、五寨、岢嵐3 個縣的種植面積約為0.67 萬hm2,已經成為當地農民重要的經濟收入[3]。山西水資源貧乏,污水灌溉面積較大,重金屬鎘極易在土壤中富集,而關于其對紅蕓豆種植的影響尚未見相關報道。
本研究采用手持式葉綠素熒光儀,分析在土壤中不同濃度鎘的脅迫下紅蕓豆幼苗葉綠素熒光參數的變化,進而辨析鎘對紅蕓豆幼苗光合作用的影響,從而為農田土壤中的鎘污染治理提供合理的依據。
1 材料和方法
1.1 試驗材料
供試紅蕓豆品種為金蕓3 號,購買于忻州市匯豐糧業有限公司。
1.2 主要儀器與試劑
電子分析天平(ME104T,梅特勒),手持式葉綠素熒光儀(配置PEA PLUS 軟件,Pocket PEA,英國Hansatech 公司);氯化鎘(分析純,南試化學試劑有限公司)、蒸餾水。
1.3 試驗方法
1.3.1 紅蕓豆幼苗種植及鎘脅迫處理 根據土壤污染的實際程度共設置5 個處理組,其Cd2+質量分數分別為5,10,15,20,25 mg/kg,以一個空白為對照組,每組分別設3 個平行。選取大小相同的紅蕓豆種子放到容器里浸泡12 h,播種到經過鎘處理過的育苗盆土壤中,每隔48 h 澆水一次,培養至所有的紅蕓豆幼苗長出2 片子葉,在幼苗長出2 片子葉后的7,14 d 測定其葉片的葉綠素熒光參數。
1.3.2 葉綠素熒光測定 將葉片夾夾在紅蕓豆幼苗葉片上,并滑動鐵片將葉片夾上的圓孔完全覆蓋,暗適應20 min,暗反應結束后,將手持式葉綠素熒光儀與葉片夾相連接,滑動鐵片露出葉片夾上的圓孔,測該植株的葉綠素熒光值。
1.4 數據分析
使用PEA PLUS 軟件對試驗所測數據進行整理分析。
2 結果與分析
2.1 不同質量分數鎘脅迫7 d 葉綠素熒光參數的變化分析
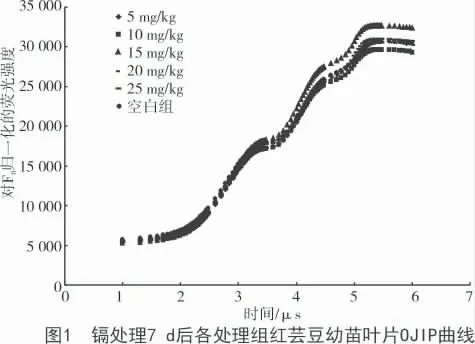
從圖1 可以看出,在O- J- I- P 曲線的P 段,鎘溶液質量分數為15 mg/kg 時,熒光強度顯著增強;鎘溶液質量分數為10 mg/kg 時,熒光強度減弱(P<0.05);而其他質量分數的鎘溶液對紅蕓豆幼苗的葉綠素熒光效應無顯著性差異。在試驗中,所使用的5 個鎘溶液質量分數脅迫的紅蕓豆幼苗在O- J段與空白對照組相比,均無顯著性差異。當鎘溶液質量分數為15 mg/kg 時,J- I 段、I- P 段、P 段曲線中的紅蕓豆幼苗葉綠素熒光顯著增強,且呈現為擴大趨勢。鎘溶液質量分數為10 mg/kg 時,I- P 段和P段的葉綠素熒光強度開始減弱,且在P 段呈現為顯著減弱效應。
Fv/Fm(即φP0)表示的是反應初始時PSⅡ的最大光能轉換效率,是當所有PSⅡ反應均處于開放狀態時的量子產量,是應用最廣、使用頻率最高的一個熒光參數[4]。該參數在非脅迫條件下變化極小,并不受物種和生長條件的影響,脅迫條件下該參數會顯著下降[5]。由表1 可知,Fv/Fm 在5~15 mg/kg區間,隨著土壤中鎘離子質量分數的增加,光能轉化率增強,而隨著鎘離子質量分數的進一步增加達到20 mg/kg 時,Fv/Fm 開始降低,光和轉化率減弱,與空白對照組相比,光能轉化效率總體呈現下降的趨勢。

表1 鎘處理7 d 后各處理組葉綠素熒光儀的參數變化
TR0/RC 表示PSⅡ系統中單位反應中心捕獲的光能,鎘離子質量分數為20 mg/kg 處理組的值最大;ET0/RC 表示單位反應中心傳遞的光能,鎘離子質量分數為10,15 mg/kg 處理組的值較大;ABS/RC表示單位反應中心吸收的光能,鎘離子質量分數為10 mg/kg 處理組的值最大;三者在趨勢中均表現為隨著質量分數的升高逐漸上升,當到達一定質量分數時,又呈現下降的趨勢。與空白對照組相比,受到不同質量分數鎘污染后,TR0/RC、ET0/RC、ABS/RC總體均呈現下降趨勢。
2.2 不同質量分數鎘脅迫14 d 葉綠素熒光參數的變化分析
由圖2 可知,鎘溶液質量分數為5 mg/kg 時,熒光強度最強;質量分數為20 mg/kg 時,熒光強度最弱。鎘離子溶液質量分數為10 mg/kg 時,其熒光強度在O- P 段高于空白對照組,而5,15,25 mg/kg 處理組均是在P 段的熒光強度超過空白對照組。
從表2 可以看出,紅蕓豆幼苗隨著土壤中鎘離子質量分數的升高,Fv/Fm 表現為先升高后降低的趨勢,其最高值出現在10 mg/kg,但是與空白對照組相比,總體呈下降趨勢。當土壤中的鎘離子質量分數為15 mg/kg 時,TR0/RC 值最大且高于空白對照組,當鎘離子質量分數為5,10 ,25 mg/kg 時,紅蕓豆幼苗的TR0/RC 值低于空白對照組。在各質量分數鎘離子污染的土壤中,ET0/RC 均小于空白對照組,但是其總體趨勢表現為隨著鎘離子質量分數的增加呈先升高后下降,其出現峰值的質量分數為15 mg/kg。ABS/RC 值在15 mg/kg 處理組顯著上升(P<0.05),10,20 mg/kg 處理組無顯著性差異,5,25 mg/kg 處理組低于空白對照組,但其總體趨勢也表現為先升高后下降。
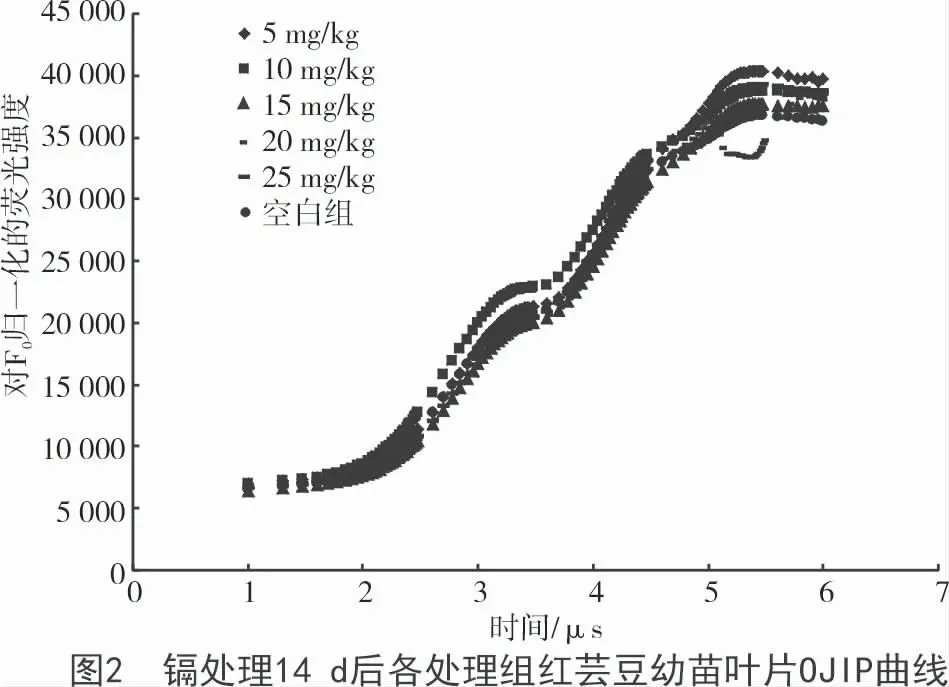
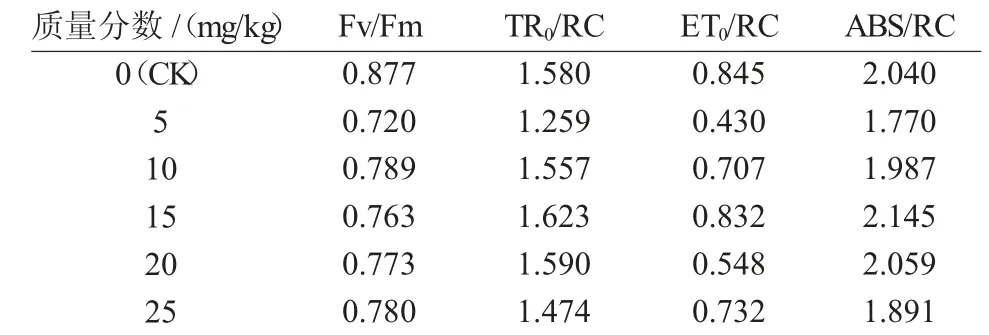
表2 鎘處理14 d 后各處理組葉綠素熒光儀的參數變化
3 結論與討論
本研究結果表明,在鎘處理7 d 時,Cd2+質量分數為15 mg/kg 時,紅蕓豆幼苗葉片的光能轉化率最高,10 mg/kg 處理組最低;而鎘處理14 d 時,質量分數為10 mg/kg 處理組的紅蕓豆幼苗葉片的光能轉化率最高,5 mg/kg 處理組最低,但是隨著鎘處理紅蕓豆幼苗時間的延長,高濃度鎘對紅蕓豆幼苗葉片的光合反應速率抑制作用明顯。
從不同時間段采集的紅蕓豆幼苗葉片的OJIP曲線和所測得相關熒光參數可以得出,紅蕓豆幼苗隨著土壤中鎘離子濃度的增加,光合反應速率和光量子產量會出現一個暫時的上升,但隨著鎘離子濃度的增加,總體呈現下降趨勢。同樣在為期14 d 的鎘離子對幼苗處理期間,隨著時間的延長,紅蕓豆幼苗的光合反應速率和光量子產量呈現下降趨勢。究其原因,可能是由于鎘會破壞植物體的光合反應系統[6],從而導致光量子產量、光合反應速率降低。但是,紅蕓豆幼苗在低濃度Cd 的脅迫下,光量子產量和光合暗反應速率暫時恢復正常,其可能的原因是,植物通過結構和代謝上的變化對逆境進行響應,第一,利用沉積或螯合作用將吸收的Cd 固定,避免其對植物造成傷害;第二,通過改變體內代謝過程,使其能夠適應脅迫環境,主要表現在抗氧化能力、滲透調節能力以及植物內源物質上[7]。
趙子豪等[8]研究發現,南方四季楊幼苗雌株光合結構對鎘的敏感性明顯高于雄株,在鎘處理的第45 天,其凈光合速率、PSⅡ最大光化學量子產量顯著下降,最小熒光和非光化學淬滅系數顯著上升,各種表現均表明PSⅡ中心已經受到鎘的危害,由此也造成了植物對水分的利用效率顯著降低。鎘脅迫對水稻農藝性狀的影響表現出明顯的品種間差異。彭鷗等[9]研究發現,培養液中的Cd2+質量分數為1 mg/mL 時,水稻中嘉早17 在不同生育期的根長和株高均顯著降低,水稻不能正常結實,籽粒空癟;水稻泰優390 的根長和株高未受到顯著影響,但產量下降了54.62%。趙麗娟等[10]研究發現,土壤中的Cd2+質量分數在5 mg/kg 時即可引起菠菜幼苗葉綠素含量的顯著下降。
玉米是典型C4 植物,CO2的固定還原分別在葉肉細胞和維管束鞘細胞中進行,研究發現,Cd2+不僅影響玉米光合器官發育,還可以破壞光合結構,并通過抑制光化學反應降低玉米光合效率[11]。VACULíK 等[12]分別用5,50 μmol/L 的Cd2+處理玉米幼苗10 d,發現葉肉細胞結構和葉綠體超微結構未受到影響,但玉米葉鞘細胞和葉綠體類囊體發生解體;當Cd2+濃度為80 μmol/L 時,葉片光合色素含量降低15%~30%[13],推測其可能的原因是Cd2+的吸收抑制了Fe 攝入,從而使鐵氧化還原蛋白減少,影響了光誘導的氧化還原反應,導致葉綠素生物合成受損[14]。ANJUM等[15]用225,300,375 μmol/L的Cd2+處理玉米,結果表明,成熟期的玉米葉綠素a、葉綠素b 含量和凈光合速率都呈下降趨勢。植物光合速率由光反應和暗反應2 個階段共同決定,經過Cd2+處理后,玉米品種3223 和3299 的Fv/Fm 值分別比空白組下降24%和16%,Cd2+對玉米光化學反應脅迫存在品種之間的差異[16]。
植物對環境的脅迫最直觀的反應表現在形態,但往往滯后于生理反應,一旦傷害造成,則難以恢復[17]。葉綠素熒光與植物的光合作用反應過程緊密相關,因此,Cd2+對紅蕓豆幼苗的影響可以通過葉綠素熒光誘導曲線反映出來。在本試驗中,紅蕓豆幼苗的光合速率雖然在低濃度Cd2+的脅迫下出現了暫時性的恢復,但是隨著土壤中Cd2+濃度的增加和脅迫時間的增加,總體仍呈現出下降的趨勢,由此可見,土壤中富集的Cd2+會對紅蕓豆的光合系統產生一定的破壞作用。
猜你喜歡
中學生數理化·中考版(2022年10期)2022-11-10 09:37:42
中學生數理化·八年級物理人教版(2022年12期)2022-02-14 07:08:42
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(2018年6期)2018-04-22 03:16:54
產品可靠性報告(2017年7期)2017-09-05 09:49:12
中學生數理化·八年級物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
