脈沖電場促進黑曲霉生長提高產酶活性
2019-11-28 08:53:34劉振宇宋艷波高瑞梅李林葳賈麗艷
農業工程學報 2019年19期
關鍵詞:質量
劉振宇,宋艷波,高瑞梅,李林葳,楊 政,賈麗艷
(1.山西農業大學信息科學與工程學院,晉中030801;2.山西農業大學生命科學學院,晉中030801;3.山西農業大學工學院,晉中030801;4.山西農業大學食品科學與工程學院,晉中030801)
0 引 言
黑曲霉(Aspergillus niger)是國際公認的生物技術中最重要的微生物之一。從1919 年黑曲霉生產檸檬酸被工業化開發,首次獲得實際應用開始,至今黑曲霉一直是工業應用中重要的研究對象[1]。目前,黑曲霉發酵產生的多種代謝產物在發酵飼料、食品添加配料、工業酶制劑以及醫藥方面都有重大作用[2-4]。黑曲霉是多種酶的來源,成為眾所周知的外源蛋白和酶的細胞生產工廠,其生產的糖化酶已經實現了規模化工業應用[5]。而且,檸檬酸和許多黑曲霉的酶被美國食品和藥物管理局(Food and Drug Administration,FDA)認為是GRAS(Generally regarded as safe)產品,此外,黑曲霉還可用于生物轉化和廢物處理[1]。因此研究黑曲霉菌種特性、培養環境、發酵體系,采用物理、化學和生物技術等手段改造黑曲霉菌種,提高菌株對目標代謝物的生產能力成為目前國內外研究的熱點。研究黑曲霉菌絲生長形態和特異性蛋白酶活性與高水平葡萄糖淀粉酶活性關系[6],優化生產工藝提高黑曲霉中殼聚糖產量和功能活性[7],采用60Co射線和紫外輻照結合處理黑曲霉孢子獲得環氧化物水解酶(epoxide hydrolase,EH)高產菌株[8]。這些技術本身具有一定限制,如產生不需要的化學殘留物和需要很長時間培養等。而脈沖電場作為非熱處理技術,以其綠色環保,節約能源和可提高某些微生物細胞活性和發酵能力,有望發展為解決這些問題的可替代技術。
脈沖電場(pulsed electric field,PEF)處理是一種新型的細胞電穿孔技術[9]。在外加PEF 作用下,細胞膜上形成小孔的過程即為電穿孔。在一定條件下形成的電穿孔是不可逆的,使細胞死亡,可以有效殺滅食品中的多種食源性,腐敗性和致病性微生物,廣泛用于果汁等流動和果醬等半流動及鮮果和肉類保鮮貯藏及品質改善[9-12];在醫學中作為聯合治療技術和直接治療方法進行腫瘤細胞殺滅[13-14]。在某些條件下,形成了可逆的膜電穿孔,細胞是活的,但細胞透性增大,這利于組織中活性化合物和基因提取[15];利于果蔬干燥時水分散失,提高干燥速率[16]。近年來PEF 對活細胞的刺激越來越受到廣泛關注,用于促進植物種子萌發[17-19]和改良菌種等,如用PEF 處理酵母,既可改變釀酒酵母的培養周期,促進細胞分裂同步化[20],也可增加酵母細胞生長和酒精生產率[21],改善酵母發酵特性,提高酵母菌的代謝,成為刺激微生物反應(即發酵)的積極工具[22]。目前,關于PEF對黑曲霉作用報道較少,本研究以黑曲霉糖化酶高產菌株為作用對象,探討脈沖工作參數對黑曲霉生長和產糖化酶性能影響,力求獲得兩者間的互作規律,為黑曲霉生產效能的提高提供一種新的技術和方法。
1 材料與方法
1.1 試驗材料
試驗菌株:黑曲霉AS3.4309,購買于中國科學院微生物研究所。
試驗試劑:馬鈴薯葡萄糖(PDA,potato dextrose agar)液體培養基:馬鈴薯200 g切成小塊,加適量水煮沸30 min過濾,濾液中加20 g 蔗糖,用蒸餾水補至1 000 mL;培養基加入20 g 瓊脂即為固體培養基。察氏培養基:蒸餾水1 000 mL,硝酸鈉3 g,蔗糖30 g,磷酸氫二鉀1 g,硫酸鎂(MgSO4·7H2O)0.5 g,氯化鉀0.5 g,硫酸亞鐵0.01 g,瓊脂20 g,加熱溶解。蛋白胨:1 g蛋白胨,蒸餾水溶解,定容至1 000 mL,滅菌備用。
試驗儀器:SW-CJ-2G 型超凈臺,蘇州江東精密儀器有限公司;ECM830 型脈沖發生器,美國BTX 公司;JS-2000W-A 型交流電源電壓轉換器,上海景賽電子科技有限公司;SPX-300-F 生化培養箱,上海龍躍儀器設備有限公司;血球計數板(QIUJING,25 格×16 格),上海求精生化試劑儀器有限公司;E100 型顯微鏡,上海維翰光電科技有限公司;Megafuge 8R 臺式高速冷凍離心機,Thermo Fisher 美國公司;DH5000-AB 干燥箱,德州市昊誠實驗儀器有限公司;THZ-98A 恒溫振蕩培養箱,上海一恒科學儀器有限公司;UV752N 紫外可見分光光度計,上海佑科儀器儀表有限公司;HH-42 水浴鍋,福州精科儀器儀表有限公司;DYCZ-24A 電泳儀,北京市六一儀器廠;Chemixrsf化學發光凝膠成像儀,美國Bio-Rad伯樂公司等。
1.2 試驗方法
1.2.1 黑曲霉的活化和孢子懸液制備
黑曲霉接種于斜面培養基上,28 ℃培養7 d,4 ℃保存。向上述培養物加適量無菌蛋白胨,用接種針刮下培養物,無菌紗布濾去菌絲體,使孢子分布均勻,用血球計數板計量3 次孢子數,求其平均值,得到孢子濃度為4.2×106個/mL的菌懸液。
1.2.2 菌懸液的PEF處理及培養
吸取150 μL 孢子懸液于電擊杯(聚碳酸酯材質,兩電極間距2 mm)中進行脈沖處理:取100 μL 放入裝有25 mL PDA 液體培養基離心管中,28 ℃、150 r/min培養60 h,測定菌絲干質量;另取10 μL 接于裝有30 mL PDA 液體培養基的錐形瓶中培養108 h,測定糖化酶活性;取2 μL 進行微室培養(察氏培養基)10~12 h,顯微鏡下觀察孢子萌發;取1 μL 接于PDA 固定平板中央,觀察菌絲生長,以未進行PEF 處理的黑曲霉為對照。所有試驗重復3次。
1.2.3 指標測定方法
1)菌絲干質量測定[23]:用定量濾紙過濾培養物,70 ℃烘干至恒質量。
2)糖化酶活性的測定:培養后的菌液,4 ℃5 700 r/min離心20 min,上清液即為粗酶液[24],酶活性測定采用3,5-二硝基水楊酸法[25]。
酶活性單位為pH值5、40 ℃和15 min反應時間分解淀粉產生1 μmol 葡萄糖(0.18 mg)所需酶量[24]。
3)黑曲霉孢子萌發測定[26-27]:菌絲長度比孢子直徑一半長時,認為孢子已萌發。
孢子萌發率=已萌發孢子數目(/已萌發孢子數+未萌發孢子數)×100%。
4)同工酶測定
孢子懸液接于裝有25 mL PDA 液體培養基的錐形瓶中培養72 h,過濾并收集菌團,蒸餾水沖洗,吸干。稱菌絲團放入預冷的研缽,加適量石英砂和0.1 mol/L pH值8.0 Tris-HCl提取液(1 g菌絲/2 mL提取液)冰浴研磨,低溫浸提0.5 h或過夜。12 000 r/min 4 ℃離心15 min,上清液用于酯酶(EST)同工酶分析[28]。
聚丙烯酰胺凝膠電泳:分離膠質量分數12%,pH值8.9;濃縮膠5%,pH 值6.8,電泳緩沖液為Tris-Gly pH 值8.3 系統;初始電壓160 V,分離膠電壓為240 V。染液配制參考文獻[29]稍作修改:50 mg固藍B鹽(氯化四氮鄰二甲氧基苯胺),溶于150 mL 0.1 mol/L pH 值6.2磷酸緩沖液,過濾;50 mg α-乙酸萘酯和50 mg β-乙酸萘酯溶于1 mL 丙酮,倒入固藍溶液中,邊倒邊攪拌。電泳結束后將膠轉移至染色液中,37 ℃黑暗染色至褐色條帶清晰,水漂洗,7%的醋酸固定,用凝膠成像儀拍照。
1.2.4 試驗設計
依據脈沖儀器性能、菌液承受能力和前期試驗研究結果選擇脈沖參數和參數范圍。采用三因素五水平中心組合設計(central composite experimental design,CCD)確立脈沖強度(5~15 kV/cm)、脈沖持續時間(10~100 μs)和脈沖數(50~99)3 個脈沖參數組合,以黑曲霉菌絲干質量和產糖化酶活性為主要響應指標,研究這些指標在不同脈沖參數下的變化規律,優化脈沖工藝。各變量的編碼值和實際值如表1。
1.2.5 統計分析
所有結果用平均值表示。數據統計分析和建模均由Design Expert 10.0.7 統計分析軟件(Stat-Ease 公司,明尼阿波里斯市,美國)完成。試驗數據進行多元回歸和顯著性分析(P<0.05)。從回歸模型中生成響應面圖,直觀反映響應值與自變量之間的關系,確定最優條件。
2 結果與分析
PEF 處理技術可以產生多種生物學效應,如殺滅食源性和腐源性致病菌,從而被廣泛用于液態食品加工中;增加植物和肉類等固態食品傳質,改善加工中的傳質品質;促進植物種子的代謝活動,提升種子活力;提高物料干燥速率;增加細胞內活性物質提取率;生物大分子改性和刺激酵母細胞生長等。而要達到以上某種特定效應的前提是執行相應的PEF 加工參數[30]。研究獲得了PEF 工作參數對黑曲霉生長和產糖化酶活性關系。
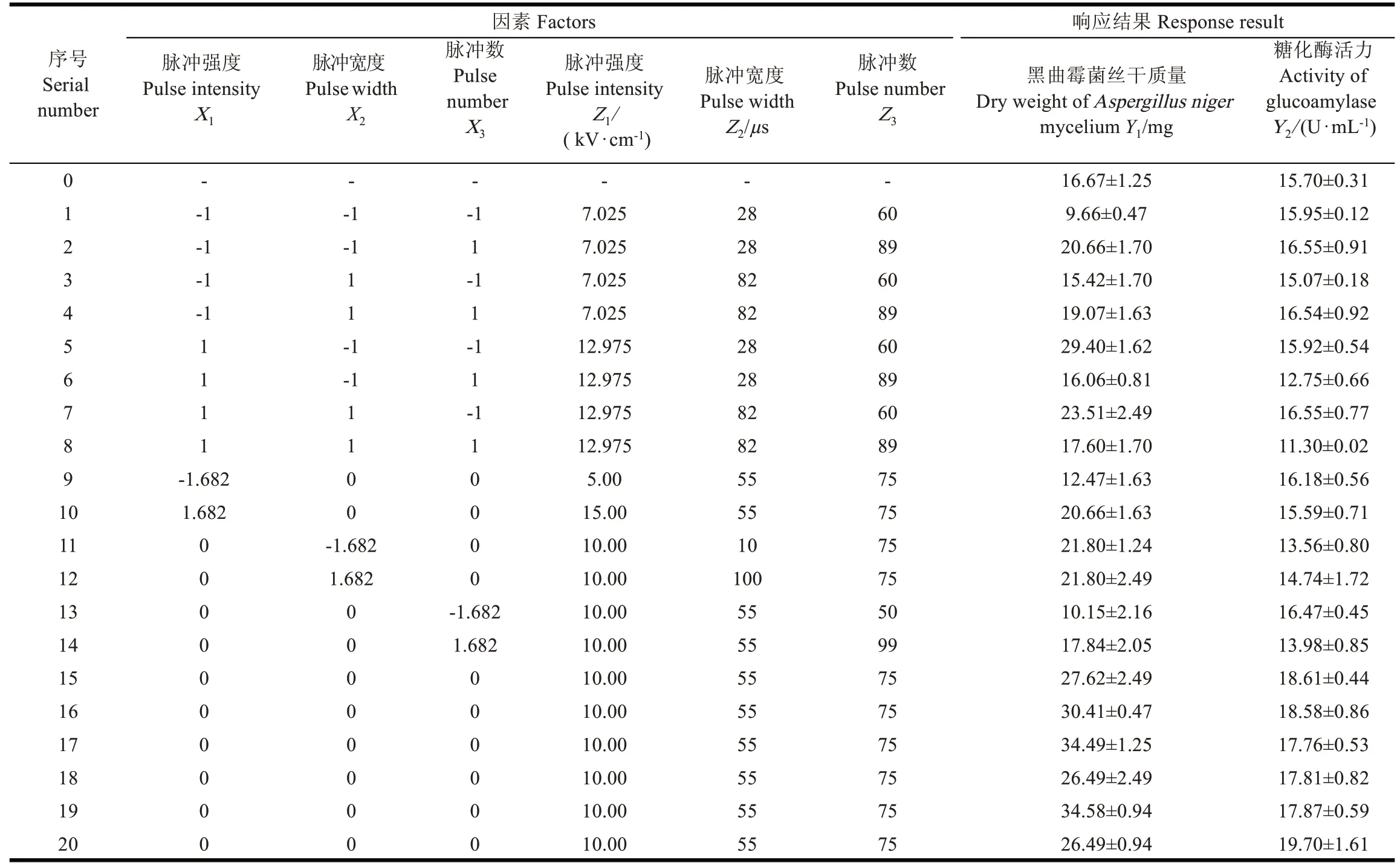
表1 試驗設計與實測結果Table1 Experimental design and measured results
2.1 PEF 參數對黑曲霉菌絲干質量和產糖化酶活性影響規律
2.1.1 PEF作用對黑曲霉菌絲干質量和產糖化酶活性影響
PEF 處理對黑曲霉菌絲干質量和產糖化酶性能有明顯影響,不同參數效果不同。未經PEF 處理的黑曲霉菌絲干質量和糖化酶活性分別為(16.67±1.25)mg 和(15.70±0.31)U/mL;PEF 處理后,菌絲干質量和糖化酶活性最大達(34.58±0.94)mg 和(19.70±1.61)U/mL,比對照分別增加了107%和25.47%(見表1)。在個別參數下菌絲干質量和糖化酶活性比未處理低,從表1 不難看出,在低電場強度處理下菌絲干質量未增反而下降;而脈沖數較高的處理下,產酶活性下降。Zhao 等采用20 kV/cm、100 和200 ms 的短時間PEF 處理釀酒酵母細胞,發現α-葡萄糖苷酶、N -乙酰-β-糖苷酶和α-甘露糖苷酶活性增加;當PEF 處理時間超過200 ms 時,觀察到了蛋白團聚體的形成,同時發現這些酶活性降低甚至完全喪失[31]。在本研究中,脈沖處理個數高,脈沖作用時間積累效應導致黑曲霉產糖化酶活性的下降,與上述研究結果一致。
因此,尋求脈沖參數及參數間相互作用對響應指標影響規律,確立脈沖參數與響應指標間的量化關系尤為重要。
2.1.2 PEF作用參數與兩響應指標回歸方程建立與分析
用統計分析軟件對表1 結果進行回歸分析和顯著性檢驗,建立了脈沖強度、脈沖時間和脈沖數3 個自變量和黑曲霉菌絲干質量、產糖化酶活力2 個響應指標關系的二階多項式方程:
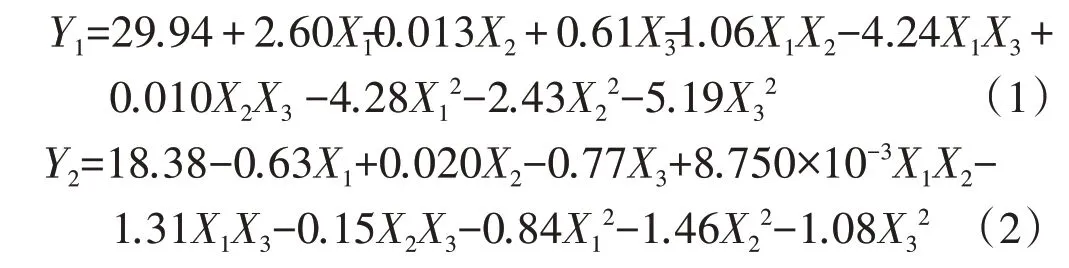
式中X1,X2,X3分別為脈沖強度、脈沖持續時間和脈沖數的編碼水平;Y1為黑曲霉菌絲干質量,mg;Y2為糖化酶活力,U/mL。
采用方差分析評價兩模型的充分性和適合度,以相應的P 值評價模型的顯著性(表2)。黑曲霉菌絲干質量(Y1)F 值為6.89,糖化酶活性(Y2)F 值為12.02,F 值較大,說明了模型的正確性;P<0.01,模型均顯著;兩模型失擬項均不顯著,且決定系數R2分別為0.861 2 和0.915 4,值較高,表明該模型可用。方程能反映和預測試驗選定參數范圍內3 個脈沖參數與2 個響應指標間的關系。
回歸系數檢驗顯示:對黑曲霉菌絲干質量影響最大的因素是脈沖強度(P<0.01),其次是脈沖數和脈沖持續時間。根據回歸模型方差結果繪制3D 效果圖:從圖1a
可以看出,曲面比較陡,表示脈沖強度與脈沖數互作時,對黑曲霉菌絲干質量影響顯著(P<0.01)。而脈沖強度與脈沖寬度互作、脈沖寬度與脈沖數互作對黑曲霉菌絲干質量的影響不顯著(表2)。
與菌絲干質量略有不同,影響糖化酶最大的因素是脈沖數(P<0.01),其次是脈沖強度(P<0.05)和脈沖寬度。綜合響應曲面圖和測定結果表,只有脈沖強度和脈沖數的互作效應3D響應曲面圖的凸起幅度較強,互作效應顯著P<0.05,與對黑曲霉干質量作用結果一致(圖1b)。綜合脈沖參數對2 個響應指標影響可知,脈沖強度對兩指標影響顯著,這與當前研究認為PEF 的作用主要取決于電場強度[32]結果一致。另外,脈沖數與脈沖強度的互作也顯著影響兩指標變化,應用時也需重點考慮。
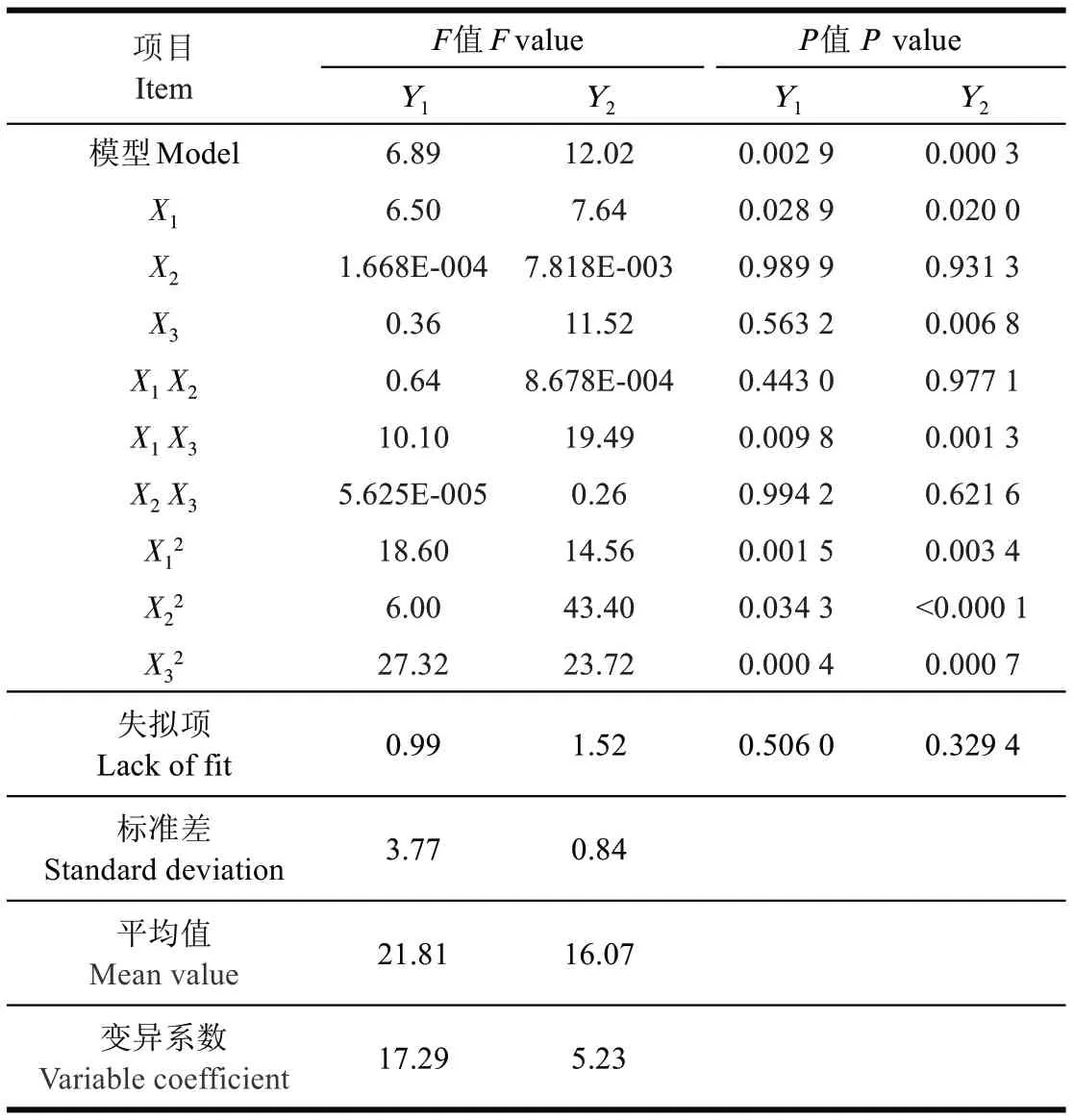
表2 PEF參數對黑曲霉菌絲干質量和產糖化酶活性影響的方差分析Table 2 Variance analysis from influence of PEF parameters on Aspergillus niger mycelium dry weight and activity of glucoamylase
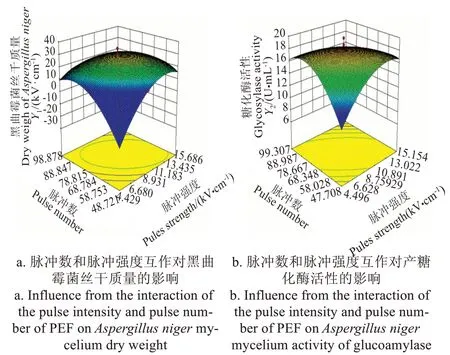
圖1 PEF脈沖數和脈沖強度兩參數互作對黑曲霉菌絲干質量和產糖化酶活性的影響Fig.1 Influence from the interaction of the pulse intensity and pulse number of PEF on Aspergillus niger mycelium dry eight and activity of glucoamylase
2.2 PEF對黑曲霉菌落形態特征的影響
PEF 處理對黑曲霉菌落生長形態產生了明顯影響(圖2)。在表1脈沖參數處理下,所有菌落雖然都呈圓形生長,但未處理的菌落(圖2 中0)可見清晰的菌落圈;而PEF 處理后,有些菌落圈隆起高,出現星狀褶皺,菌落圈較難分辨,孢子數量顯著增加,孢子顏色沒變但孢子周圍出現明顯不同的深黃色(圖2 中PEF-7 和PEF-11 分別為表1 中序號7 和11 處理)。說明,PEF 不但刺激了黑曲霉生長,也影響其代謝。
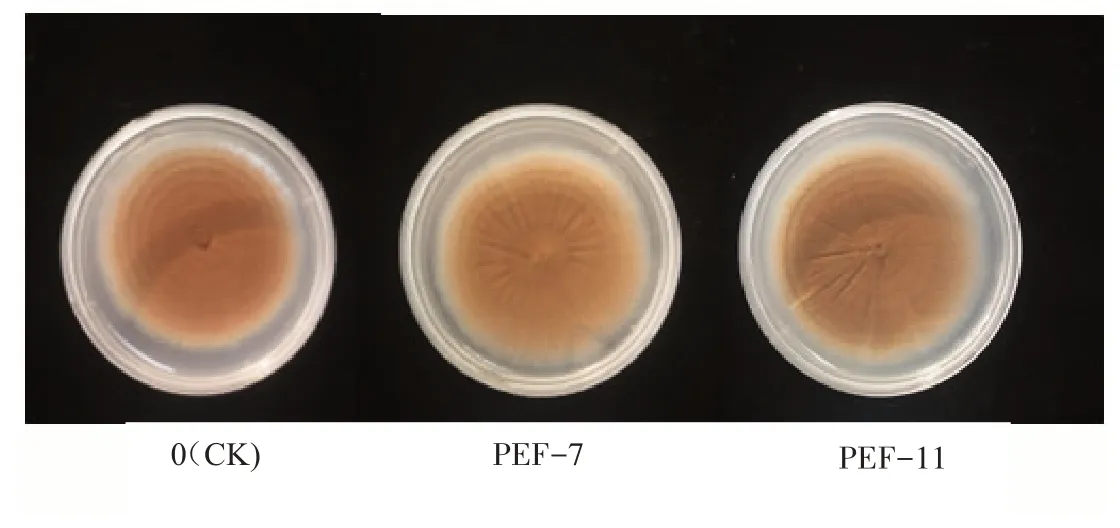
圖2 PEF對黑曲霉菌落形態特征的影響Fig.2 Influence of PEF on Aspergillus niger colony shape feature
2.3 PEF對黑曲霉孢子萌發的影響
顯微鏡下觀察和統計黑曲霉孢子萌發情況(圖3)。未經PEF 處理的黑曲霉孢子萌發率為14.06%±0.59%,采用表1 中PEF 工作參數處理后,孢子萌發率為25.00%±0.33%~46.54%±0.55%,比對照增加了77.8%~231%。據此認為PEF促進菌絲生長原因之一是刺激了孢子萌發。
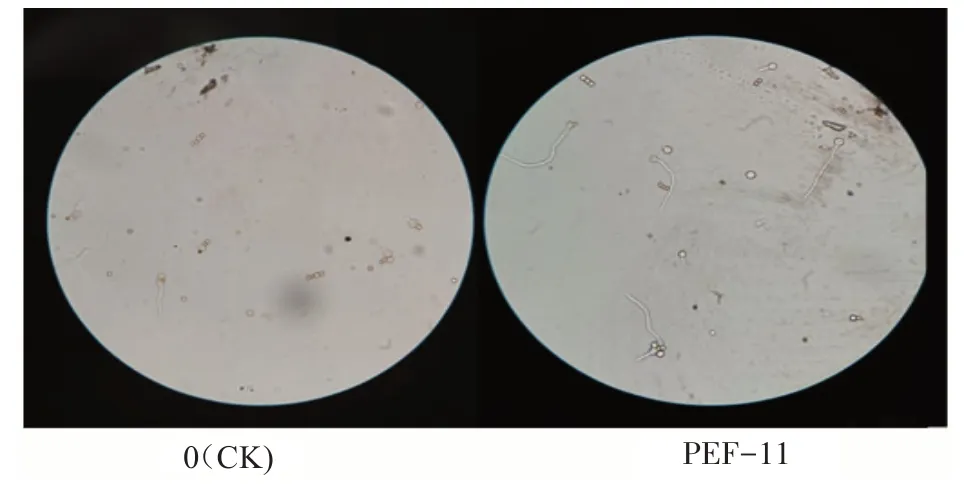
圖3 PEF對黑曲霉孢子萌發的影響(×400)Fig.3 Influence of PEF on Aspergillus niger spore germination(×400)
2.4 PEF對EST同工酶的影響
PEF處理在某些條件下可能對菌株產生誘變作用[33],為了檢測PEF處理后的黑曲霉是否發生質變,基于菌落形態特征觀察,對生長褶皺、顏色和疏密度等表現出差異的處理和對照同時收獲孢子,進行酯酶同工酶酶譜分析。
由圖4a 和4b 可知,黑曲霉的EST 同工酶出現了10條譜帶,標記為EST-01~EST-10,其遷移率為0.267~0.750。EST-01~EST-05 條帶強度明顯強于其他條帶,說明這5條條帶對EST 活性貢獻較大。而EST-08~EST-10條帶最弱,對EST活性貢獻最小。同工酶是基因表達產物,是遺傳與變異的重要指標;而酯酶同工酶由于易于檢測,且具有高度多態性,被作為遺傳研究中常見的標記物,廣泛用于微生物、昆蟲和植物物種鑒定及分類與進化研究中[34-36]。本文,PEF 處理組與對照組的EST-01~EST-08 的遷移率都相同。說明PEF處理并未對黑曲霉的EST同工酶形成明顯遺傳變異。另外,酯酶活性可作為微生物細胞代謝性能的指標,圖中編號7~9 樣本EST-01 和EST-02條帶顏色比對照深,11~16 號樣本與對照無明顯差異,而其他樣本比對照淺。這一結果表明PEF 影響了黑曲霉的代謝性能。

圖4 PEF對黑曲霉酯酶同工酶影響Fig.4 Influence of PEF on esterase isozyme of Aspergillus niger
2.5 工藝參數的優化驗證
黑曲霉菌絲干質量和黑曲霉糖化酶活性在相應約束條件下都應達到最大值,即Y1max和Y2max確立為目標函數,經Design Expert 10.0.7 統計分析軟件對兩個回歸模型分別優化求解。在此基礎上,利用多目標決策理論及非線性優化方法,綜合目標函數采用線性加權法進行規劃求解,由于糖化酶活性是主要目標,其加權系數K2取0.6,則黑曲霉菌絲干質量的加權系數K1為0.4[16]。獲得綜合優化條件為脈沖強度12.974 95 kV/cm、脈寬為53.517 μs、脈沖數66.052,相應的黑曲霉菌絲干質量為28.662 mg,糖化酶活性為17.746 U/mL。經修正,選擇脈沖強度為12.975 kV/cm、脈寬為54 μs、脈沖數66,獲得黑曲霉的菌絲干質量和糖化酶活性分別為28.05 mg和18.01 U/mL,與預測值基本一致,表明優化結果合理可靠,比對照提高了68.27%和14.71%,可以作為進一步試驗研究的指導。
3 討論
3.1 PEF對黑曲霉作用機理分析
PEF 處理影響黑曲霉分生孢子活力,增加檸檬酸產量[37]。在本研究中,PEF 處理后產糖化酶能力增強,菌絲干質量增加,這與PEF刺激孢子萌發有關,與對萌發中和萌發后代謝刺激也有關。PEF 處理酵母細胞研究中發現,PEF 處理促進細胞中鋅離子、鎂離子和鈣離子積累[38-39],促進果糖等物質吸收,刺激酵母細胞代謝[22]。以及造成細胞代謝過程中酶的變化,如PEF 處理新鮮藍莓提高了花青素濃度,改變了其在藍莓中的分布,可能通過影響反應動力學中涉及的分子,如酶,改變花青素降解的機制[11]。作為一種絲狀真菌,黑曲霉孢子萌發常需要較長的延滯期,因此縮短孢子萌發時間,可以提高發酵效率,而PEF處理可以通過改變細胞膜組分排列,產生電穿孔,增加細胞膜滲透性或激活轉運蛋白質[40],增加對水分、離子和有益營養物質的吸收運轉,活化代謝過程中的酶,促進細胞代謝,從而促進生長,提高其產酶性能。
3.2 PEF參數優化
PEF 可以刺激細胞代謝,也可以殺死細胞。這主要取決于脈沖工作參數。研究認為PEF的作用主要取決于電場的強度,Sotelo 等研究了中、輕度PEF(電場強度0.3和2.5 kV/cm,脈沖頻率100 Hz,脈沖寬度20 μs)處理櫻桃提高了其可食品質且不影響益生菌的生長[41]。電場強度5.0 kV/cm,脈沖寬度20 μs,電場頻率1 Hz下處理酵母細胞15 min 后培養20 h,鎂和鋅的積累比對照分別增加了1.5 倍和2 倍[38],過高電場強度會使細胞損傷嚴重,甚至死亡。因此,高的電場強度(10~80 kV/cm)常被用于巴氏殺菌,使微生物膜破壞到無法修復的程度,最終死亡[42]。影響果汁質量最重要的電學參數不只電場強度,還有處理時間[10],用于有機物提取研究中,使細胞膜發生可逆穿孔,增加細胞滲透性的脈沖強度范圍在1~10 kV之間,脈沖數在5~50 之間[42-43]。可見,PEF 作用中脈沖工藝參數確立的重要性。本文研究同樣得出脈沖強度對黑曲霉生長和產糖化酶活性能力影響最突出,并且與脈沖個數互作效應明顯。因此,在對不同類型,不同發育時期等不同特點細胞進行PEF 處理時,對參數進行選擇和優化是首要任務,研究為該技術在今后工廠化應用中提供理論基礎。
4 結論
1)PEF處理黑曲霉孢子,不同的電場強度、脈沖寬度以及脈沖數會導致黑曲霉菌絲干質量和生產糖化酶的能力有所改變,電場強度和脈沖數及其相互作用是影響兩指標的主要參數。電場處理后的黑曲霉,其細胞膜通透性和酶活性會增大,代謝速度變快,可以更好的促進孢子萌發。脈沖強度為12.975 kV/cm、脈寬為54 μs、脈沖數66,獲得黑曲霉的菌絲干質量和糖化酶活性分別為28.05 mg和18.01 U/mL。
2)酯酶同工酶分析未發現PEF對黑曲霉有誘變的作用,只是通過刺激黑曲霉生長,影響細胞代謝而影響其生長和產糖化酶能力,可見PEF技術是不破壞黑曲霉本質,且處理過程簡單,生產效率高,是一種有前景的用于提高生物工程菌發酵效率的有力工具。
猜你喜歡
中學生數理化·中考版(2022年10期)2022-11-10 09:37:42
中學生數理化·八年級物理人教版(2022年12期)2022-02-14 07:08:42
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(2018年6期)2018-04-22 03:16:54
產品可靠性報告(2017年7期)2017-09-05 09:49:12
中學生數理化·八年級物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
