基于轉錄組測序的鐵皮石斛植物甾醇生物合成相關基因分析
2019-12-06 09:03:54林江波王偉英鄒暉戴藝民
熱帶亞熱帶植物學報 2019年6期
林江波,王偉英,鄒暉,戴藝民
基于轉錄組測序的鐵皮石斛植物甾醇生物合成相關基因分析
林江波,王偉英,鄒暉,戴藝民*
(福建省農業科學院亞熱帶農業研究所,福建 漳州 363005)
為了解鐵皮石斛()植物甾醇的生物合成途徑,利用Illumina HiSeq 4000高通量測序技術對莖和葉進行轉錄組測序,比較植物甾醇生物合成關鍵酶基因的表達。結果表明,共獲得43 085條Unigenes,其中24 459條在Nr、Swiss-prot、KOG和KEGG數據庫獲得注釋,7 333條獲得共同注釋。KEGG代謝途徑分析表明,鐵皮石斛植物甾醇生物合成分為3個階段,共有50個Unigenes (30種酶)參與。表達分析表明,和的表達量明顯高于和;成熟期莖、葉的表達量比生長期高,則生長期高于成熟期;同一時期,和在葉的表達量都比莖高。這為鐵皮石斛植物甾醇的開發利用和調控植物甾醇生物合成研究提供了科學依據。
鐵皮石斛;高通量測序;植物甾醇;基因; 表達;代謝
植物甾醇不僅是生物膜的重要組成成分,還是一類無色無味的甾體類活性成分,不溶于水,溶于有機溶劑,屬于三萜類化合物,含有28~29個C原子,主要骨架是4個環的環戊烷多氫菲,C-3羥基是活性基團[1-2],與羧酸形成植物甾醇酯,具有更好的水溶性和生理活性。植物甾醇以游離或結合甾醇的方式廣泛存在于植物體中,除常見的豆甾醇、-谷甾醇、菜油甾醇和菜籽甾醇外,已經報道了超過250種的衍生物,如甾體皂苷、甾體糖苷生物堿、植物甾酮和油菜素內酯等[3]。植物甾醇不僅具有抗氧化和延緩衰老的功能[4-5],還對多種腫瘤的發生和發展具有預防和治療作用[6-9]。
鐵皮石斛()是蘭科(Orchi- daceae)石斛屬多年生藥食兩用植物,具有增強免疫力、抗腫瘤、降血糖、抗氧化和抗疲勞等生理活性[10-11]。鐵皮石斛有效成分種類多樣,除了石斛類多糖和生物堿外,還有菲類、芪類、笏酮類、倍半萜類、甾體類及香豆素等[12]。楊柳等[13]利用正己烷蒸餾法提取的鐵皮石斛干燥莖揮發性成分中含有22,23-二氫豆甾醇、-谷甾醇、豆甾醇和油菜甾醇,相對含量分別為11.093%、4.126%、3.138%和1.648%,4種甾醇占揮發性成分的20.005%。陳曉梅等[14]從固體培養鐵皮石斛原球莖甲醇提取物中分離鑒定到豆甾醇和-谷甾醇。仝晶晶等[15]分析了6個產地鐵皮石斛干燥莖的豆甾醇和-谷甾醇含量,產于云南省普洱市景谷縣的豆甾醇含量最高,為86.399g g-1, 產于浙江省樂清市雙峰鄉的-谷甾醇含量最高,為344.683g g-1。
轉錄組(RNA-seq)測序能夠獲取植物特定組織或器官在特定時期的基因表達信息,有利于挖掘藥用植物次生代謝產物合成調控關鍵酶基因。吳超等[16]對鐵皮石斛葉片進行轉錄組測序,KEGG注釋結果表明有1 006個Unigenes與次生代謝物的生物合成有關。Guo等[17]對鐵皮石斛莖轉錄組數據分析表明,有69個Unigenes參與生物堿骨架的生物合成。Zhang等[18]對鐵皮石斛生長期和成熟期進行轉錄組測序,KEGG注釋結果表明有430個糖基轉移酶,89個纖維素合酶,135個Unigenes參與果糖和甘露糖代謝。
本研究以2個生長階段的鐵皮石斛莖、葉為材料,利用Illumina HiSeq 4000高通量測序技術進行轉錄組測序,以期獲得與植物甾醇合成相關的基因信息,分析鐵皮石斛植物甾醇代謝途徑, 為鐵皮石斛植物甾醇的代謝調控研究提供科學依據。
1 材料和方法
1.1 材料
鐵皮石斛()原種采自福建省龍巖市連城縣冠豸山崖壁,經過莖段擴繁后種植于福建省農業科學院亞熱帶農業研究所資源圃,分別于生長期(2016年8月15日)和成熟期(2016年12月15日)隨機采集3株新鮮莖、葉,分別標記為T1 (生長期莖)、T2 (生長期葉)、T3 (成熟期莖)和T4 (成熟期葉),經液氮速凍后用干冰寄到廣州基迪奧生物科技有限公司進行RNA提取、轉錄組測序和Unigene功能注釋。
1.2 植物甾醇生物合成途徑分析
依據植物甾醇生物合成途徑,結合轉錄組數據GO功能分類結果,分析植物甾醇生物合成途徑的相關Unigenes。
1.3 植物甾醇生物合成關鍵酶基因表達分析
用標準化后的Reads Per Kilobase per Million mapped reads (RPKM)值來表示樣品的表達量,分析植物甾醇生物合成關鍵酶基因在鐵皮石斛不同生育期的表達模式。
2 結果和分析
2.1 轉錄組測序和數據組裝
鐵皮石斛4個樣品的原始數據(NCBI SRA號:SRP181716)經過濾除雜處理后,分別獲得22 809 182、19 492 544、21 587 660和21 058 784 reads,分別包含有3 388 594 751、2 884 066 418、3 171 710 203和3 132 109 020個核苷酸序列信息。將獲得的高質量核苷酸序列進行拼接,共獲得長度大于200 bp的Unigenes片段43 085條,N50為1 628 bp,平均長度902 bp,最長Unigene片段為20 526 bp, 長度在200~299 bp的片段最多,有13 280條,其次是300~399 bp,有5 558條,≥3 000 bp的有1 701條(圖1)。
通過BLAST比對和ESTscan預測Unigene片段的編碼區(CDS),結果表明, 有25 928條Unigenes片段含有CDS (分別獲得24 266和1 662條),占Unigenes片段的60.18%,其中CDS長度大于1 000 bp的Unigenes片段有7 550條,占總數的17.52% (圖2)。
2.2 基因功能注釋和分類
將43 085條Unigenes片段在Nr、Swiss-prot、KOG和KEGG數據庫搜索比對序列相似性,共有24 459條獲得注釋,占總數56.77%:其中,在Nr數據庫注釋的有23 460條,占54.45%,Swiss-prot數據庫有18 514條,占42.97%;KOG數據庫有15 522條,占36.03%;KEGG數據庫有8 972條, 占20.82%;在4個數據庫都得到注釋的有7 333條, 占17.02%。
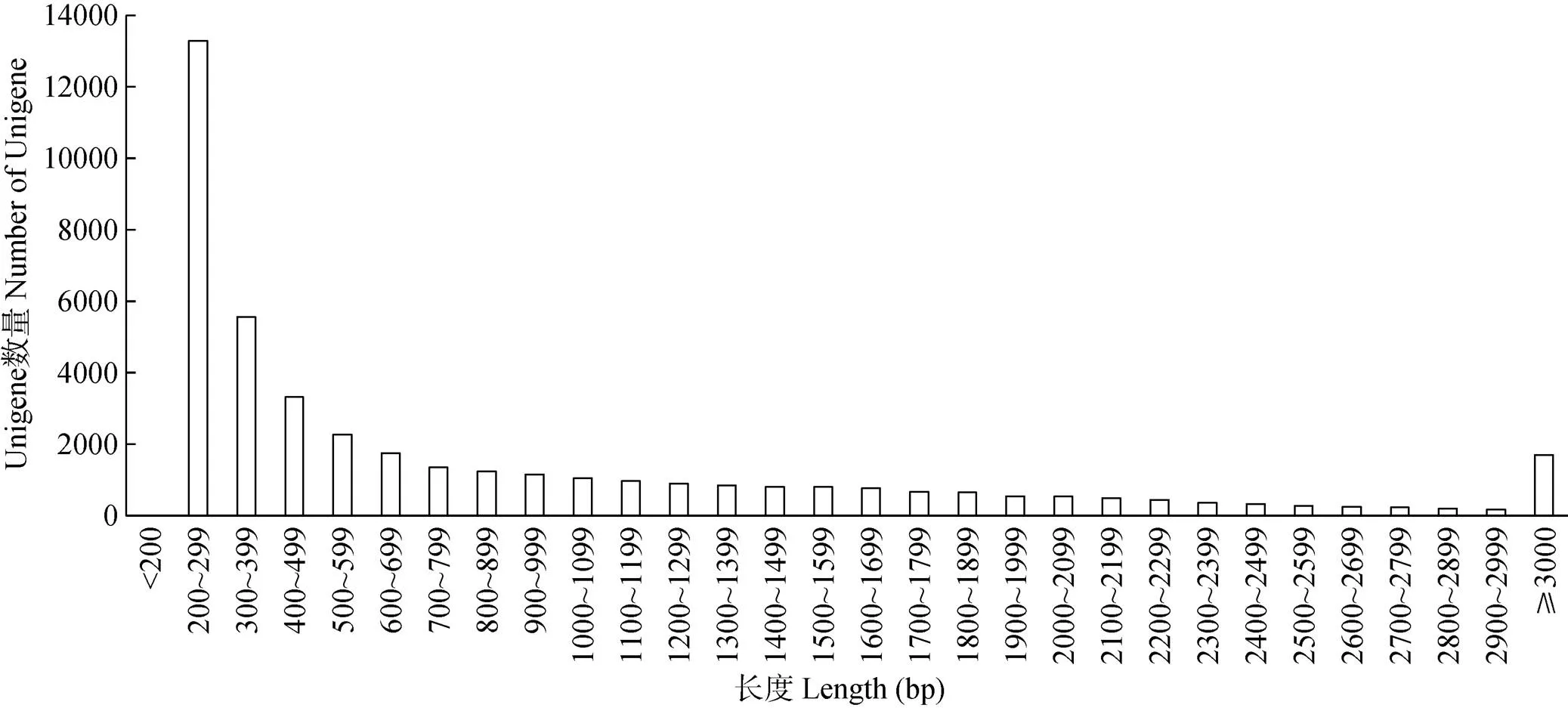
圖1 Unigene長度分布
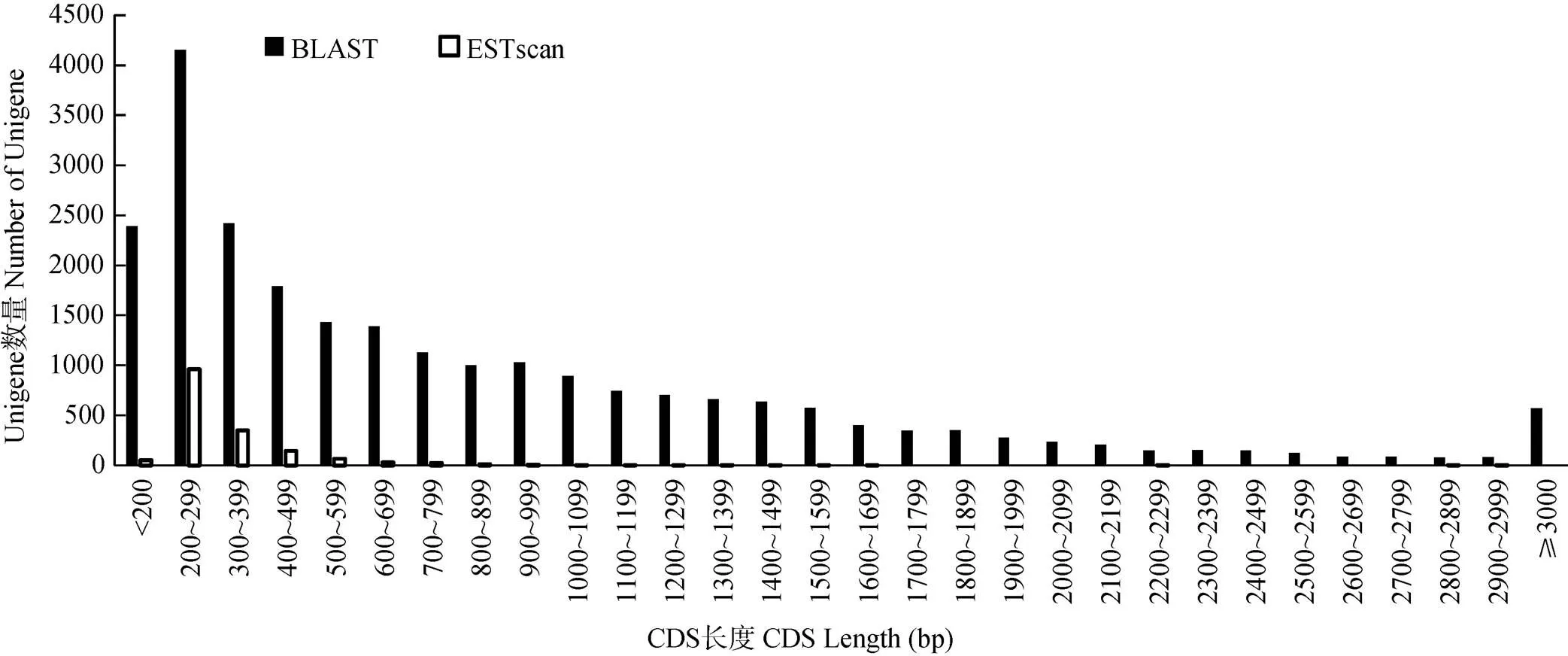
圖2 CDS長度分布
通過GO功能數據庫比對分析,共有13 710條Unigenes得到注釋和功能分類,分為3大類49個功能(表1),生物過程含20個功能,涉及31 519條Unigenes,其中代謝過程和細胞過程分別涉及7 617和7 200條;細胞組分含17個功能,涉及24 983條Unigenes,其中細胞和細胞組份分別涉及6 008和6 006條;分子功能含12個功能,涉及14 948條Unigenes,其中,催化活性有6 967條,結合的有6 448條。
KEGG注釋的8 972條Unigenes中有7 890條獲得Pathway注釋(表2),分為5大類19條代謝途徑。遺傳信息處理含有4條代謝途徑,其中翻譯的Unigenes數最多,為929條;環境信息處理含有信號轉導和膜運輸2條代謝途徑;細胞過程和有機系統各含有1條代謝途徑;代謝含有11條代謝途徑,以碳水化合物代謝最多,1 118條。

表1 鐵皮石斛Unigene GO功能分類

表2 KEGG注釋的代謝途徑
2.3 植物甾醇生物合成途徑
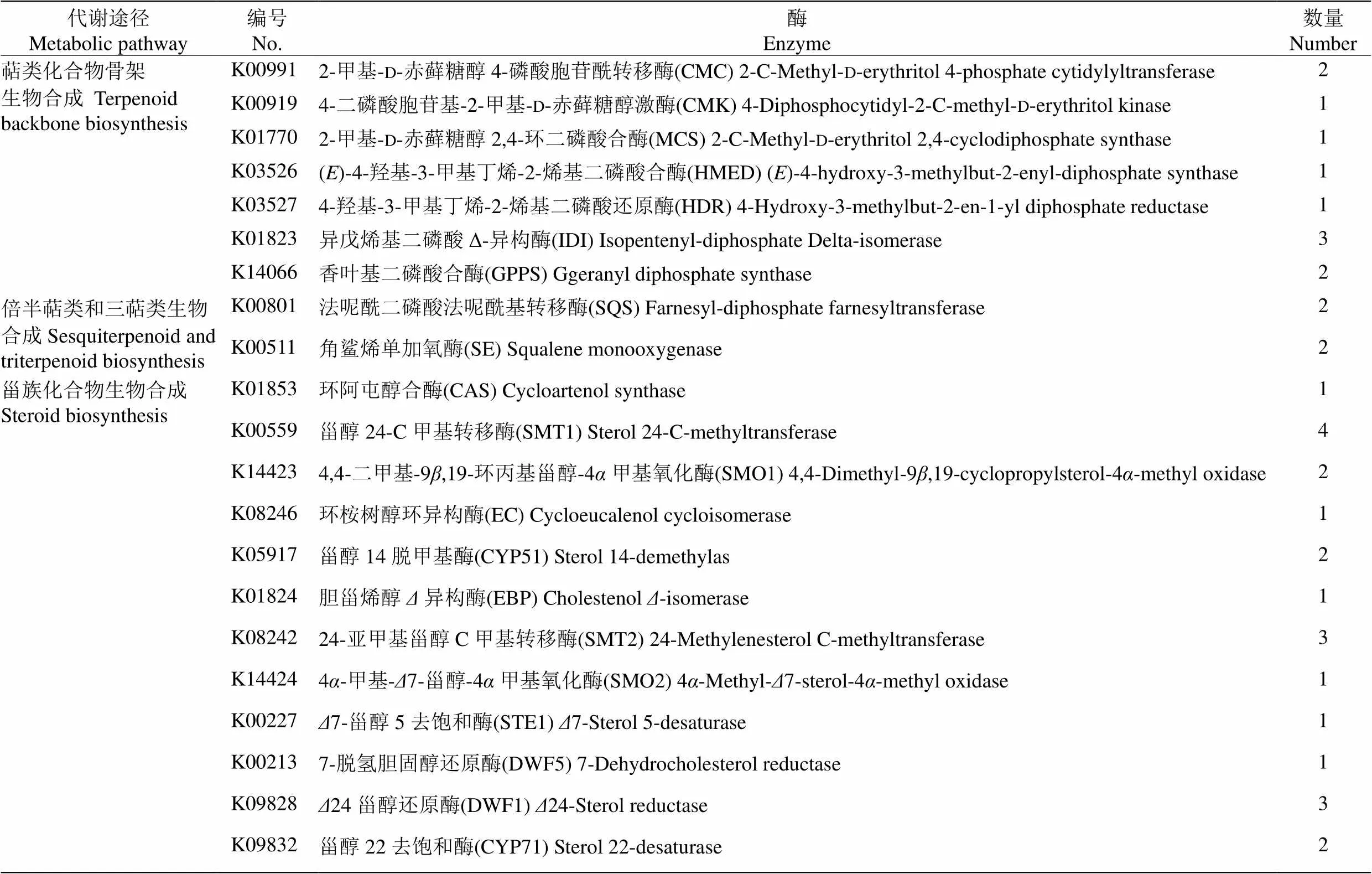
KEGG注釋結果表明,共有50個Unigenes (30種酶)參與鐵皮石斛植物甾醇的生物合成(表3)。植物甾醇生物合成途徑可分為萜類化合物骨架生物合成、倍半萜類和三萜類生物合成和甾族化合物合成3條代謝途徑。
根據Pathway分析結果預測鐵皮石斛植物甾醇生物合成路徑(圖3), 萜類化合物骨架生物合成有甲羥戊酸途徑(MVA)和丙酮酸途徑(DEP)兩種,MVA途徑從乙酰CoA到生成法尼基焦磷酸共7步7個酶(10個Unigenes)參與;DEP途徑從丙酮酸和甘油醛- 3-磷酸到生成法尼基焦磷酸共11步10個酶(15個Unigenes)參與。倍半萜類和三萜類生物合成從法尼基焦磷酸到2,3-氧化鯊烯共3步2個酶(4個Unigenes)參與。甾族化合物生物合成從2,3-氧化鯊烯到菜仔甾醇和豆甾醇生成共16步12個酶(22個Unigenes)參與。

表3 KEGG注釋的鐵皮石斛植物甾醇生物合成相關的Unigene數量
續表(Continued)
2.4 植物甾醇生物合成相關酶基因表達分析
選擇、、、、、和等7種關鍵酶基因,采用RPKM值分析他們在鐵皮石斛生長期和成熟期莖、葉中的表達量。從圖4可以看出,MVA途徑的和表達量比DEP途徑的和表達量低。在生長期和成熟期的表達量差異不大,但在莖的表達量比葉片稍高。在成熟期莖、葉的表達量比生長期莖、葉高,則生長期表達量高于成熟期;同一時期,和在葉片表達量都比莖高。
3 結論和討論
本研究通過Illumina HiSeq 4000測序平臺對鐵皮石斛2個生育期的莖、葉進行轉錄組測序,經過拼接后獲得43 085條大于200 bp的Unigenes片段,其中24 459條在Nr、Swiss-prot、KOG和KEGG數據庫獲得注釋。根據KEGG注釋結果,鐵皮石斛植物甾醇生物合成分成3個階段,共有50個Unigenes (30種酶)參與。
植物甾醇合成第一階段是法尼基焦磷酸的合成,有MVA和DEP兩種途徑。通過比較兩種途徑的2個關鍵酶基因的RPKM值,DEP途徑的和表達明顯高于MVA途徑的和,說明鐵皮石斛合成法尼基焦磷酸存在兩種途徑,但DEP途徑強于MVA途徑,這可能與鐵皮石斛是兼性景天酸途徑植物[19]有關。
植物甾醇根據側鏈C-24位上烷基分為甲基甾醇和乙基甾醇,通過甾醇C-24甲基轉移酶(SMT)在C-24位上分別通過1次和2次增加甲基的步驟完成[20-21]。SMT1把1個甲基從S-腺苷甲硫氨酸(SAT)轉移到環阿屯醇的C-24位上,形成C-24甲烯基環阿屯醇;SMT2則催化在C-24甲烯基環阿屯醇的衍生物24-亞甲基膽甾烯醇的C-24甲基上再添加1個甲基,形成乙基甾醇[22-23],因此,SMT1是甲基甾醇和乙基甾醇合成限速酶,SMT2是乙基甾醇合成限速酶。在煙草植株中過表達,菜油甾醇和谷甾醇分別提高80%和50%,而環阿屯醇和膽固醇含量分別下降53%和34%。過表達顯著降低了膽固醇和C-24甲基甾醇含量,而谷甾醇含量顯著提高[24-25]。本研究發現相同生育期鐵皮石斛的和在葉片中的表達量都明顯高于莖,推測葉片中植物甾醇的合成比莖高。8月份的葉表達量明顯高于,12月份則相反, 推測8月份葉片合成乙基甾醇多,而12月份則合成甲基甾醇多,這有待于對鐵皮石斛和功能的進一步鑒定,分析不同生長階段植物甾醇組分和含量。

圖3 鐵皮石斛植物甾醇生物合成途徑
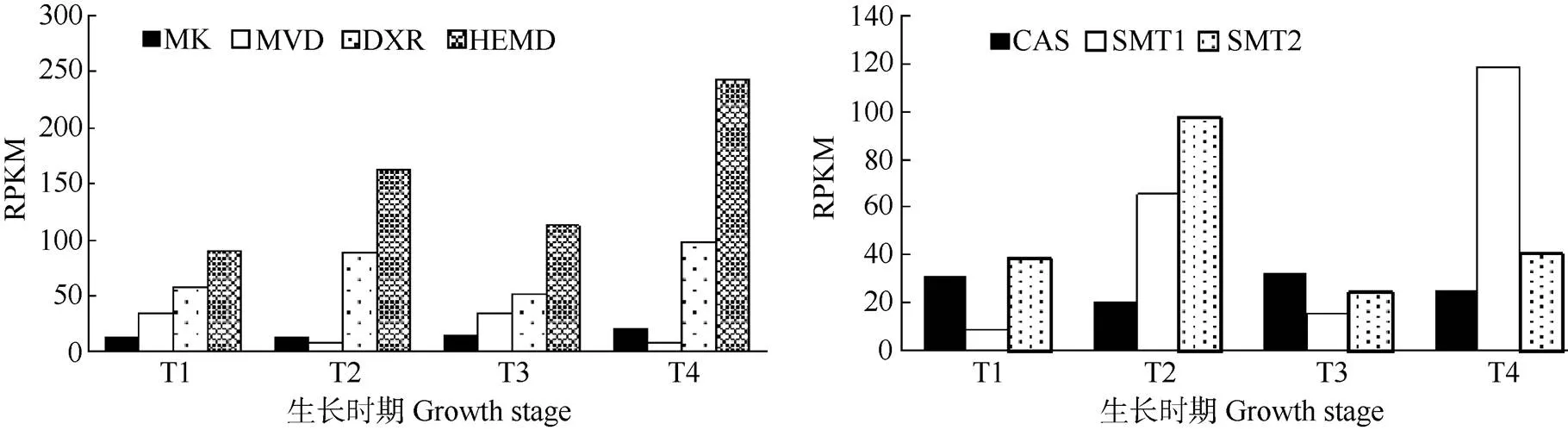
圖4 鐵皮石斛7個植物甾醇生物合成相關基因的表達模式。T1: 生長期莖; T2: 生長期葉; T3: 成熟期莖; T4: 成熟期葉。
[1] ZHANG X C, HAN Z H. Sterols biosynthesis and their physiological role in plants [J]. Plant Physiol Commun, 2001, 37(5): 452-457. doi: 10.13592/j.cnki.ppj.2001.05.033.張喜春, 韓振海. 植物體內甾醇的合成和生理作用[J]. 植物生理學通訊, 2001, 37(5): 452-457. doi: 10.13592/j.cnki.ppj.2001.05.033.
[2] DAI G H, WANG M. Property and function of phytosterol and its application in food [J]. J Kaifeng Inst Educ, 2016, 36(2): 261-262. doi: 10.3969/j.issn.1008-9640.2016.02.122. 代廣輝, 王敏. 植物甾醇性質、功能及其在食品中的應用[J]. 開封教育學院學報, 2016, 36(2): 261-262. doi: 10.3969/j.issn.1008-9640. 2016.02.122.
[3] Nes W D. Biosynthesis of cholesterol and other sterols [J]. Chem Rev, 2011, 111(10): 6423-6451. doi: 10.1021/cr200021m.
[4] WU S M, WU M C, MA L. Antioxidant effect of phytosterols in high grade rapeseed cooking oil (Ι): Antioxidant effect at normal temperature [J]. China Oils Fats, 2003, 28(4): 52-54. doi: 10.3321/j.issn:1003- 7969.2003.04.017.吳時敏, 吳謀成, 馬莉. 植物甾醇在菜籽高級烹調油中的抗氧化作用(I): 常溫下抗氧化作用的研究[J]. 中國油脂, 2003, 28(4): 52-54. doi: 10.3321/j.issn:1003-7969.2003.04.017.
[5] Rudkowska I. Plant sterols and stanols for healthy ageing [J]. Maturitas, 2010, 66(2): 158-162. doi: 10.1016/j.maturitas.2009.12.015.
[6] CAO M, OUYANG L. Study on development of anti-tumor effect of phytosterols [J]. Pract Pharm Clin Remed, 2015, 18(9): 1104-1107. doi: 10.14053/j.cnki.ppcr.201509026.曹玫, 歐陽露. 植物甾醇的抗腫瘤作用及其機制研究進展[J]. 實用藥物與臨床, 2015, 18(9): 1104-1107. doi: 10.14053/j.cnki.ppcr. 201509026.
[7] ZHOU L Y, XU C, WANG L Q, et al. Effect of-sitosterol on proli- feration and apoptosis of lung cancer cell line A549 [J]. Int J Lab Med, 2016, 37(7): 865-867. doi: 10.3969/j.issn.1673-4130.2016.07.001.周玲玉, 徐彩, 王樂琴, 等.-谷甾醇對人肺癌A549細胞增殖及凋亡的影響[J]. 國際檢驗醫學雜志, 2016, 37(7): 865-867. doi: 10. 3969/j.issn.1673-4130.2016.07.001.
[8] CHENG X A, ZHOU X W, ZHANG S M, et al. Effects of phytosterol on growth inhibiting and apoptosis induction of melanoma cells [J]. Guangdong Agric Sci, 2014, 41(10): 94-97. doi: 10.3969/j.issn.1004- 874X.2014.10.023.程杏安, 周曉武, 張淑明, 等. 植物甾醇對黑色素瘤細胞的生長抑制及凋亡誘導作用[J]. 廣東農業科學, 2014, 41(10): 94-97. doi: 10. 3969/j.issn.1004-874X.2014.10.023.
[9] ZHANG S, YUE W, WANG L, et al. Inhibitive effect of stigmasterol from hedyotis diffusa willd. on hepatoma cells&and its influence on transplanted H22 tumor cell’s multiplication cycle, apop- tosis [J]. Progr Mod Biomed, 2008, 8(11): 2016-2018. doi: 10.13241/ j.cnki.pmb.2008.11.017.張碩, 岳旺, 王蕾, 等. 白花蛇舌草豆甾醇對肝癌細胞的體內外抑制作用及對其增殖周期、凋亡的影響[J]. 現代生物醫學進展, 2008, 8(11): 2016-2018. doi: 10.13241/j.cnki.pmb.2008.11.017.
[10] LIAO J J, LI W J, LI J J, et al. Advances in the of constituents and pharmacological effects ofWall. ex. Lind. [J]. J Guangdong Ind Techn Coll, 2015, 14(3): 16-19,26. doi: 10.3969/j.issn. 1672-1950.2015.03.004.廖俊杰, 李文靜, 李進進, 等. 鐵皮石斛有效成分與功效研究進展[J]. 廣東輕工職業技術學院學報, 2015, 14(3): 16-19,26. doi: 10. 3969/j.issn.1672-1950.2015.03.004.
[11] CHEN S L, WU Z G, JIANG W, et al. Isolation and purification of three acidic polysaccharides fromand their antioxidant activities[J]. J Zhejiang Agric Sci, 2016, 57(6): 838-844. doi: 10.16178/j.issn.0528-9017.20160613. 陳松林, 吳志剛, 姜武, 等. 鐵皮石斛3種酸性多糖的分離純化及體外抗氧化活性[J]. 浙江農業科學, 2016, 57(6): 838-844. doi: 10. 16178/j.issn.0528-9017.20160613.
[12] SUN H, HU Q, JIN H, et al. Research advances in chemical consti- tuents and pharmacological activities ofcaulis [J]. Chin J Exp Trad Med Form, 2017, 23(11): 225-2348. doi: 10.13422/j. cnki.syfjx.2017110225.孫恒, 胡強, 金航, 等. 鐵皮石斛化學成分及藥理活性研究進展[J].中國實驗方劑學雜志, 2017, 23(11): 225-234. doi: 10.13422/j.cnki. syfjx.2017110225.
[13] YANG L, LIU S J, HU J M, et al. Determination of volatile components from stem ofby GC-MS [J]. Mod Chin Med, 2013, 15(5): 362-364. doi: 10.3969/j.issn.1673-4890.2013. 05.003.楊柳, 劉守金, 胡江苗, 等. GC-MS法檢測鐵皮石斛莖中揮發性成分[J]. 中國現代中藥, 2013, 15(5): 362-364. doi: 10.3969/j.issn. 1673-4890.2013.05.003.
[14] CHEN X M, YANG J S, GUO S X, et al. Studies on the chemical constituents of solid culturedprotocorms [J]. Mod Chin Med, 2014, 16(4): 265-268. doi: 10.13313/j.issn.1673-4890. 2014.04.001.陳曉梅, 楊峻山, 郭順星, 等. 固體培養鐵皮石斛原球莖化學成分研究[J]. 中國現代中藥, 2014, 16(4): 265-268. doi: 10.13313/j.issn. 1673-4890.2014.04.001.
[15] TONG J J, YU H, ZENG W B, et al. Study on functional components offrom different culturing areas [J]. J Yunnan Univ Trad Chin Med, 2017, 40(5): 81-84. doi: 10.19288/j.cnki.issn. 1000-2723.2017.05.018.仝晶晶, 虞泓, 曾文波, 等. 6個不同產地鐵皮石斛功效成分的分析研究[J]. 云南中醫學院學報, 2017, 40(5): 81-84. doi: 10.19288/j. cnki.issn.1000-2723.2017.05.018.
[16] WU C, PENG J, XIANG L, et al. Sequencing and analysis of the transcriptome of[J]. Mol Plant Breed, 2016, 14(12): 3334-3346. doi: 10.13271/j.mpb.014.003334.吳超, 彭娟, 向林, 等. 基于高通量測序的鐵皮石斛葉片轉錄組分析[J]. 分子植物育種, 2016, 14(12): 3334-3346. doi: 10.13271/j.mpb. 014.003334.
[17] Guo X, Li Y, Li C F, et al. Analysis of thetranscriptome reveals putative alkaloid biosynthetic genes and genetic markers [J]. Gene, 2013, 527(1): 131-138. doi: 10.1016/j.gene.2013. 05.073.
[18] Zhang J X, He C M, Wu K L, et al. Transcriptome analysis ofand its application to the identification of genes associated with polysaccharide synthesis [J/OL]. Front Plant Sci, 2016, 7: 5. doi: 10.3389/fpls.2016.00005.
[19] SU W H, ZHANG G F. The photosynthesis pathway in leaves of[J]. Acta Phytoecol Sin, 2003, 27(5): 631-637. doi: 10.17521/cjpe.2003.0091.蘇文華, 張光飛. 鐵皮石斛葉片光合作用的碳代謝途徑[J]. 植物生態學報, 2003, 27(5): 631-637. doi: 10.17521/cjpe.2003.0091.
[20] Diener A C, Li H X, Zhou W X, et al.controls the level of cholesterol in plants [J]. Plant Cell, 2000, 12(6): 853-870. doi: 10.1105/tpc.12.6.853.
[21] Schaeffer A, Bronner R, Benveniste P, et al. The ratio of campesterol to sitosterol that modulates growth inis controlled by[J]. Plant J, 2001, 25(6): 605-615. doi: 10.1046/j.1365-313x.2001.00994.x.
[22] BENVENISTE P. Biosynthesis and accumulation of sterols [J]. Annu Rev Plant Biol, 2004, 55: 429-457. doi: 10.1146/annurev.arplant.55. 031903.141616.
[23] Grebenok R J, Galbraith D W, Penna D D. Characterization ofendosperm C-24 sterol methyltransferase: One of two types of sterol methyltransferase in higher plants [J]. Plant Mol Biol, 1997, 34(6): 891-896. doi: 10.1023/A:1005818210641.
[24] Sitbon F, Jonsson L. Sterol composition and growth of transgenic tobacco plants expressing type-1 and type-2 sterol methyltransferases [J]. Planta, 2001, 212(4): 568-572. doi: 10.1007/s004250000417.
[25] HOLMBERG N, HARKER M, GIBBARD C L, et al. Sterol C-24 methyltransferase type 1 controls the flux of carbon into sterol biosyn- thesis in tobacco seed [J]. Plant Physiol, 2002, 130(1): 303-311. doi: 10.1104/pp.004226.
Analysis of Related Genes in Phytosterol Biosynthesis inBased on Transcriptome Sequencing Technology
LIN Jiang-bo, WANG Wei-ying, ZOU Hui, DAI Yi-min*
(Subtropical Agriculture Research Institute, Fujian Academy of Agricultural Sciences, Zhangzhou 363005, Fujian, China)
In order to understand the biosynthetic pathway of phytosterol in, the transcriptome sequence of stems and leaves at two growth stages was analyzed by using Illumina HiSeq 4000 high-throughput sequencing method, and the expressions of key enzyme genes for phytosterois synthesis were compared. The results showed that a total of 43 085 Unigenes were obtained by transcriptome sequencing, of which 24 459 Unigeneswere annotated in Nr, Swiss-prot, KOG and KEGG databases, and 7 333 were commonly annotated. KEGG metabolic pathway analysis showed that phytosterol biosynthesis ofcould be divided into three stages with 50 Unigenes (30 enzymes) involved. The expressions ofandwere significantly higher in stems and leaves than that ofand. The expression ofat mature stage was higher than that at growth stage, while the expression ofwas the opposite. The expressions ofandwere higher in leaves than that in stems at the same stage. These would be lay a foundation for the development and utilization of phytosterol and regulating phytosterol biosynthesis in.
; High-throughput sequencing; Phytosterol; Gene; Expression; Metabolism
10.11926/jtsb.4025
2018-12-03
2019-03-04
福建省自然科學基金面上項目(2018J01119)資助
This work was supported by the Natural Science Foundation of Fujian Province (Grant No. 2018J01119).
林江波(1976~ ),男,碩士,副研究員,從事農業生物技術研究。E-mail: 345953257@qq.com
Corresponding author. E-mail: dymttcn@163.com
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
航空世界(2020年10期)2020-01-19 14:36:20
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
