蘇云金芽胞桿菌SG3-7菌株的鑒定及殺蟲活性研究
2019-12-06 04:36:10代小娟秦培剛宋金東賀志有
陜西農業科學 2019年10期
關 鵬,代小娟,秦培剛,宋金東,賀志有
(1. 渭南職業技術學院 農學院,陜西 渭南 714026;2. 新疆醫科大學 基礎醫學部,新疆 烏魯木齊 830011)
蘇云金芽胞桿菌(Bacillusthuringiensis, Bt)是在土壤、塵埃、植物表面、昆蟲尸體以及污水等介質中廣泛存在的一種革蘭氏陽性細菌,由于其在芽胞形成過程中產生球形、立方體形、橢球形、菱形以及不規則形狀等伴胞晶體[1-3],又稱殺蟲晶體蛋白,對多種農林及衛生害蟲具有特異性殺蟲活性,而對人類及家畜無毒害,且不污染環境,因此成為全世界廣泛應用的微生物殺蟲劑之一。
隨著Bt菌株及其殺蟲功能基因在農林和衛生害蟲生物防治領域的大量應用,害蟲耐藥性問題日益嚴重,生物防治效率顯著降低,因此,害蟲耐藥性問題已成為Bt資源應用的一個瓶頸[4-5]。近些年來,廣大科研工作者通過不斷挖掘廣譜高效殺蟲活性的野生型Bt菌株[6],克隆大量高毒力的Bt殺蟲基因[7-8],以及通過不同Bt殺蟲基因進行片段置換或共表達等分子生物學手段解決害蟲耐藥性問題[9-10],提高Bt生物防治的效果,因此Bt菌株資源和殺蟲功能基因的研究成為Bt研究的熱點之一。
Bt SG3-7菌株是一株從秦嶺山區灌木叢土壤中分離的,能產生大量不規則伴胞晶體的野生菌株,本研究擬對該菌株的伴胞晶體形態、伴胞晶體編碼基因的類型以及殺蟲活性等方面進行分析,為該菌株在農業害蟲生物防治中的應用提供理論基礎。
1 材料與方法
1.1 材料
1.1.1 實驗菌株 野生型Bt SG3-7菌株是從秦嶺山區灌木叢土壤中分離得到并保存于本實驗室。
1.1.2 測試昆蟲 甜菜夜蛾(Spodopteraexigua)和棉鈴蟲(Helicoverpaarmigera)2齡期幼蟲通過本實驗室人工飼料進行飼養;菜青蟲(Pierisrapae)2齡期幼蟲通過新鮮甘藍喂養。
1.1.3 培養基 1/2LB、LB固體和液體培養基配置方法參見文獻[11]。
1.2 方法
1.2.1 Bt SG3-7菌株的活化及培養 吸取10 μL-20 °C保存的SG3-7菌株,接種在100 mL LB液體培養基中,30 °C,180 r·min-1振蕩培養10~12 h,然后吸取2 mL培養液,按1:100比例轉接入1/2 LB液體培養基中,在30 °C,180 r·min-1條件下振蕩培養48~72 h,最后離心收集胞晶混合物。
1.2.2 Bt SG3-7菌株顯微觀察 取少量上述胞晶混合物先后用預冷的1 mol·L-1NaCl洗滌3次,預冷的無菌水洗滌2次,最后加入適量預冷的無菌水制成懸液。吸取2-3 μL懸液在載玻片上涂布均勻,經染色后進行光學顯微鏡觀察,最后對懸液中胞晶混合物進行掃描電鏡觀察。
1.2.3 Bt SG3-7菌株中伴胞晶體蛋白的電泳分析 上述懸液經超聲波破碎后,離心獲得細胞裂解物,采用Sch?gger 等人方法對Bt SG3-7菌株中伴胞晶體蛋白進行SDS-PAGE電泳分析[12]。
1.2.4 Bt SG3-7菌株質粒DNA提取及殺蟲基因型鑒定 參考關鵬等人方法對Bt SG3-7菌株中質粒DNA進行提取[11]。以該質粒DNA為PCR擴增模板,利用PCR-RFLP方法對該菌株中伴胞晶體蛋白編碼基因的類型進行鑒定,引物序列參見文獻[13-14]。擴增產物經與pMD19-T克隆載體連接后,轉入大腸桿菌DH5α中,經藍白斑篩選后,對陽性單克隆進行測序分析。運用NCBI數據庫中 Blast工具對測序結果進行分析。
1.2.5 Bt SG3-7菌株的殺蟲活性分析 將制備好的Bt SG3-7菌株胞晶混合物先用預冷的1 mol·L-1NaCl洗滌3次,再用預冷的無菌水洗滌一次,最后胞晶混合物經稱重后,加入10 mL無菌水后震蕩混勻,通過梯度稀釋制成10~100 μg·mL-1等 6個不同濃度的測試樣品組。對棉鈴蟲、甜菜夜蛾幼蟲生物活性測定的方法參見文獻[11]。對菜青蟲幼蟲的生物活性測定:選取新鮮的甘藍制成大小相同的小塊,然后分別均勻地浸泡在不同的測試樣品中,然后取出,陰干,放入滅菌的培養皿中[15]。每個培養皿中放入2頭幼蟲,每個測試樣品組做3個重復,以無菌水浸泡的甘藍作為陰性對照。室溫培養3~5 d,然后統計結果并用SPSS10.0軟件分析計算LC50。
2 結果與分析
2.1 Bt SG3-7菌株的芽胞及伴胞晶體形態觀察
通過顯微觀察發現,Bt SG3-7菌株在1/2 LB液體培養基中培養72 h后,有大量不規則形狀的伴胞晶體及橢圓狀芽胞產生,且芽胞與晶體呈分離狀態。(圖1-A,圖1-B)。
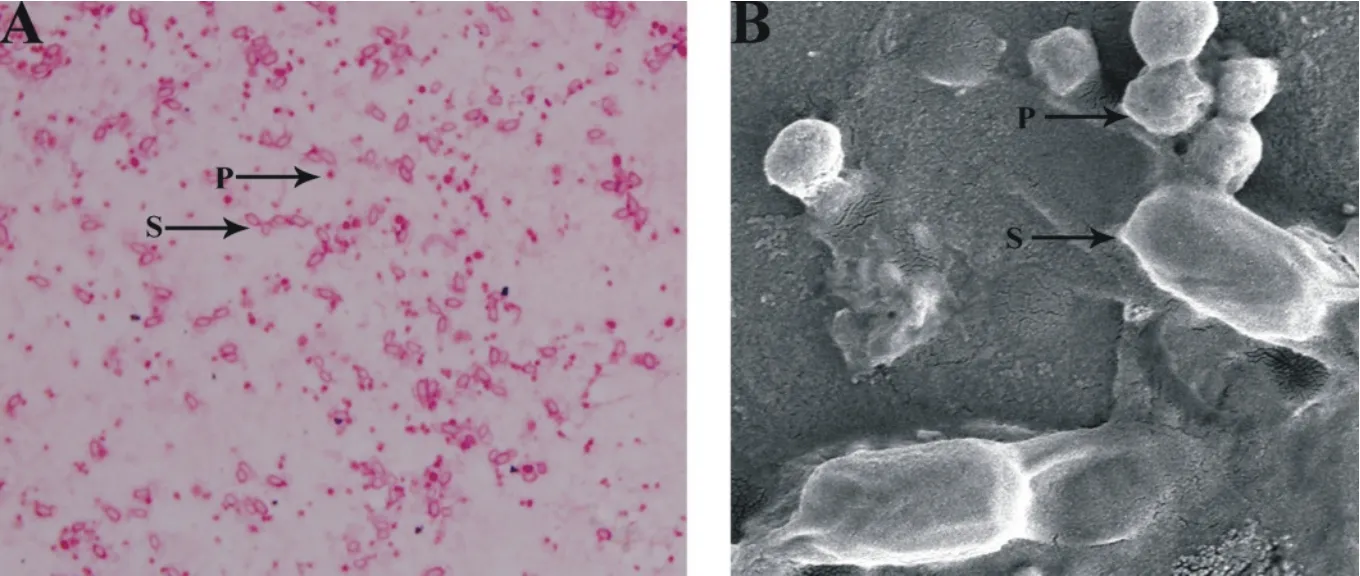
圖1 BtSG3-7菌株芽胞和伴胞晶體的顯微觀察
A:Bt SG3-7芽胞和伴胞晶體的光學顯微鏡觀察;B:Bt SG3-7芽胞和伴胞晶體的掃描電鏡觀察;P:伴孢晶體;S:芽胞。
2.2 Bt SG3-7菌株的伴胞晶體蛋白的電泳分析
SDS-PAGE電泳分析表明,Bt SG3-7菌株主要表達130 kDa和60 kDa分子量大小的伴胞晶體蛋白(圖2)。

圖2 BtSG3-7菌株伴胞晶體蛋白的SDS-PAGE電泳分析
M:蛋白質分子量標準;1. Bt SG3-7菌株表達的伴胞晶體蛋白。
2.3 Bt SG3-7菌株的伴胞晶體蛋白編碼基因的類型分析
采用PCR-RFLP方法對Bt SG3-7菌株的伴胞晶體蛋白編碼基因的類型分析表明,該菌株質粒DNA中攜帶cry2Ab、cry4A、cry9Ea等3個cry基因片段,以及1個cyt1Aa型基因片段。
2.4 Bt SG3-7菌株的殺蟲活性分析
Bt SG3-7菌株的胞晶混合物對鱗翅目的棉鈴蟲、甜菜夜蛾和菜青蟲幼蟲的殺蟲活性分析表明,該菌株對甜菜夜蛾和菜青蟲幼蟲具有較高殺蟲活性,其LC50分別為29.31 μg·mL-1和47.11 μg·mL-1,但對棉鈴蟲幼蟲無殺蟲活性(見表1)。

表1 BtSG3-7菌株胞晶混合物對三種鱗翅目害蟲的殺蟲活性測定
N:無殺蟲活性。
3 討論
經PCR-RFLP方法鑒定Bt SG3-7菌株內生質粒中攜帶有cry2Ab、cry4A、cry9Ea、 cyt1Aa型殺蟲晶體蛋白基因。根據以前的研究報道,cry2Ab基因表達約70 kDa分子量大小的蛋白,對鱗翅目的小菜蛾(Plutellaxylostella)、棉紅鈴蟲(Pectinophoragossypiella)等幼蟲有顯著殺蟲活性[16-17]。cry9Ea型基因編碼分子量約為130 kDa的蛋白,對粉紋夜蛾(Trichoplusiani)、小菜蛾等鱗翅目幼蟲具有特異性殺蟲活性[18-19]。cry4A 和cyt1Aa型基因分別編碼135 kDa和27 kDa的殺蟲蛋白,兩者均對雙翅目的蚊蟲具有特異性殺蟲活性[20-21]。SDS-PAGE電泳分析表明,Bt SG3-7菌株主要表達130 kDa和60 kDa伴胞晶體蛋白,因此,可推測Bt SG3-7菌株的伴胞晶體可能由多個Cry蛋白混合形成,其次,cry2Ab和cyt1Aa型基因在該菌株中表達量極低,或者該菌株中攜帶的這兩個基因可能不完整而未表達。另外,在Bt SG3-7菌株中鑒定得到的殺蟲基因均不表達60 kDa伴胞晶體蛋白,因此這表明該菌株中可能含有其他新型殺蟲基因未被鑒定。
鱗翅目的甜菜夜蛾和菜青蟲對農業中多種蔬菜危害嚴重,而人們一直采用高毒性的化學農藥對這些害蟲進行防治,其結果造成環境污染,蔬菜農藥殘留,以及害蟲耐藥性的產生,這不僅威脅到人類的健康,而且大大增加的害蟲防治的成本。本研究中Bt SG3-7菌株對甜菜夜蛾和菜青蟲幼蟲具有特異性殺蟲活性,因此該菌株可作為備用Bt菌株資源,在農業生產上對甜菜夜蛾和菜青蟲害蟲特異性生物防治中應用。
在后續的研究中,將對Bt SG3-7菌株中這些殺蟲基因進行克隆和表達研究,明確其殺蟲活性。另外,采用其他方法對該菌株中其他新的殺蟲基因進行鑒定和克隆,為該菌株在農業生產中的應用提供理論基礎。
