花生GAIP-B基因的生物信息學分析及表達研究
2019-12-09 01:52:35何佳霖趙凱周定緯李培培李忠峰馬興立張幸果殷冬梅
山東農業科學 2019年9期
何佳霖 趙凱 周定緯 李培培 李忠峰 馬興立 張幸果 殷冬梅

摘要:為研究DELLA蛋白在花生生長、育種過程中的功能,利用RT-PCR技術從花生栽培種品系花2014中克隆得到一個花生DELLA蛋白編碼基因AhGAIP-B(GenBank登錄號:XP_016191184)。序列分析結果表明,AhGAIP-B開放閱讀框(ORF)全長1 812 bp,只有一個外顯子,可編碼603個氨基酸。生物信息學分析發現,AhGAIP-B編碼的產物是一類典型DELLA蛋白,其三級結構與擬南芥GAI相似,親緣關系與大豆GmGAI1最近。進一步qRT-PCR結果顯示,AhGAIP-B基因呈組成型表達,其中在莖中表達量最高;同時,本研究還發現,外源GA3可以誘導AhGAIP-B基因上調表達,其表達量在處理后12 h最高。本結果將為進一步的花生DELLA功能研究提供理論基礎。
關鍵詞:花生;DELLA蛋白;AhGAIP-B基因;赤霉素;組織表達
中圖分類號:S565.2:Q786文獻標識號:A文章編號:1001-4942(2019)09-0028-07
Bioinformatics Analysis and Expression of GAIP-B in Peanut
He Jialin, Zhao Kai, Zhou Dingwei, Li Peipei, Li Zhongfeng, Ma Xingli, Zhang Xingguo, Yin Dongmei
(College of Agronomy, Henan Agricultural University, Zhengzhou 450002, China)
Abstract To study the function of DELLA protein in peanut growth and breeding, a peanut DELLA protein coding gene AhGAIP-B(GenBank accession number: XP_016191184) was cloned from peanut cultivar Hua 2014 by RT-PCR. Sequence analysis showed that the full length of AhGAIP-B open reading frame (ORF) was 1 812 bp with only one exon, and it encoded 603 amino acids. Bioinformatics analysis showed that the product encoded by AhGAIP-B was a typical DELLA protein, whose tertiary structure was similar to that of GAI in Arabidopsis thaliana, and its genetic relationship was the closest to GmGAI1 in soybean. qRT-PCR results showed that AhGAIP-B gene was expressed by the constitutive type in peanut with the highest expression level in stem. Exogenous GA3 could induce the up-regulation expression of AhGAIP-B gene, which reached the highest level at 12 hours after treatment. This results could provide a theoretical basis for further study of DELLA function in peanut.
Keywords Arachis hypogaea; DELLA protein; AhGAIP-B gene; Gibberellin; Tissue expression
赤霉素(Gibberellins,GAs)是一類由四環骨架構成的二萜類化合物,在植物生長發育中發揮著重要的調節作用[1-4]。目前,研究者們對擬南芥等模式植物中的GAs代謝途徑開展了深入研究,其中對于GAs信號轉導途徑的關鍵負調控因子DELLA蛋白研究較多[5]。迄今為止,擬南芥中共鑒定出五個DELLA蛋白,分別為GAI(GA Insensitive)、RGA(Repressor of GA1-3)、RGL1(REPRESSOR OF ga7-i-LIKE protein 1)、RGL2(REPRESSOR OF ga7-i-LIKE protein 1)、RGL3(REPRESSOR OF ga7-i-LIKE protein 1)[6-8]。已有研究表明,DELLA蛋白含有多個保守功能域:位于N端特異的DELLA和TVHYNP酸性保守域與GA信號感知區;C端的VHVID、RVER、Pyre及SAW結構域具有GA信號抑制功能;另外其中心區還含有核定位區域NLS[9]。當GAs存在時,可誘導 GID1蛋白構象變化,并與DELLA蛋白結合從而形成GA-GID1-DELLA三聚體;然后在26S蛋白酶泛素化作用下DELLA蛋白發生降解,解除其對植物的生長抑制作用[10]。不同DELLA蛋白在植物發育中具有不同作用,如擬南芥GAI和RGA對植物伸長生長和開花存在抑制功能[11],而RGL1和RGL2的主要功能在于調節花瓣、雄蕊和花藥的發育[12]。
在我國,花生(Arachis hypogaea)是一種重要的油料和經濟作物,但每逢夏季多雨年份,花生植株長勢過旺容易引起倒伏、影響機械化收獲,造成大幅減產。所以花生的矮化育種具有重要的現實意義,然而傳統育種方式獲得矮化優良品種周期較長。近年來基因工程技術的發展為我們提供了新的思路,通過轉入GAs抑制因子DELLA蛋白基因可調節植株的高度,從而獲得優良矮化品種[13],但在花生中還未見相關報道。
本研究從花生中克隆到一個DELLA蛋白編碼基因GAIP-B,對其進行了生物信息學分析,并進行了不同組織表達分析。研究結果可為深入研究花生DELLA蛋白的分子作用機制和分子矮化育種提供參考。
1 材料與方法
1.1 植物材料
本試驗材料為花生栽培種品系花2014。
不同組織部位取材:試驗于2018年5月1日至9月10日在河南農業大學毛莊科教園區進行,行距0.4 m。于2018年7—8月取花生的根、主莖、幼葉、花蕾和未成熟種子,3次重復,液氮速凍后-80℃保存。
赤霉素處理材料選取:花生種子催芽24 h后播種,在光周期為16L∶8D、溫度為26℃/24℃的人工氣候箱中培養,相對濕度60%,于花生三葉期(出苗后7 d)葉面噴施50 mg/L的赤霉素,在處理后0、2、4、8、12、24、48 h采集葉片樣品。每個時間點選定9株長勢一致的植株,每3株為1個重復,共設3次重復。取樣后用液氮速凍后-80℃保存。
1.2 總RNA提取和cDNA合成
取相應組織的材料在液氮中研磨,總RNA提取采用TransZOL Plant(TRANSGEN)試劑盒,cDNA的合成采用PrimeScriptTM RT reagent Kit with gDNA Eraser(TaKaRa)。引物由上海尚亞生物技術有限公司合成,其編號及序列見表1。
1.3 AhGAIP-B基因的克隆
根據NCBI(https://www.ncbi.nlm.nih.gov/)數據庫搜索到的AhGAIP-B基因設計特異性引物(表1),以花生栽培種花2014的cDNA為模板擴增目的片段AhGAIP-B。PCR反應體系為50 μL:cDNA模板2.5 μL,5×Q5 Reaction Buffer 10 μL,dNTPs 1 μL,F、R引物各25 μL,Q5 High-Fidely DNA Polymerase 0.5 μL,ddH2O補足50 μL。反應程序:98℃預變性30 s;98℃變性10 s,59℃退火30 s,72℃延伸2 min,33個循環;72℃延伸2 min。1%瓊脂糖凝膠電泳檢測PCR產物。凝膠回收試劑盒(ZOMANBIO)純化目的條帶,連接到克隆載體ZT4-Blunt上,轉化后篩選陽性克隆送至上海尚亞生物技術有限公司進行測序。
1.4 AhGAIP-B基因的生物信息學分析
用在線軟件GSDS 2.0(http://gsds.cbi.pku.edu.cn/index.php)繪制基因結構圖;利用MapInspect軟件進行染色體定位作圖;用SMART(smart.embl-heidelberg.de/)工具進行結構域的預測,最終確定DELLA基因家族序列;使用expasy工具進行蛋白質理化性質分析;用TMHMM Server v. 2.0工具(http://www.cbs.dtu.dk/services/TMHMM/)預測蛋白質跨膜結構域;蛋白質亞細胞定位用在線網站(http://www.csbi- o.sjtu.edu.cn/bio inf/plant-multi/)進行預測。
蛋白質的二級結構用在線軟件PRABI(http://ww-w.prabi.fr/)進行預測;根據在線軟件SWISS-MODEL(https://www.swissmode- l.expasy.org/interactive)分析蛋白質三級結構。
在PeanutBase(https://www.peanutbase.org/)和NCBI(https://www.ncbi.nlm.ni-h.gov/)利用在線Blast工具搜索花生AhGAIP-B蛋白的同源序列;利用本地軟件DNAMAN 6.0及在線weblogo 2.82網站(http://weblogo.berkeley.edu/)進行氨基酸多序列比對及同源性分析;使用MEGA 7.0選擇鄰接法(neighbor-joining,NJ)構建系統發育進化樹,參數設置Bootstrap=1 000。
1.5 實時熒光定量PCR分析
分別以不同組織材料總RNA反轉錄的cDNA為模板,以Actin為內參基因,擴增體系及程序參照TaKaRa公司的SYBR Premix Ex TaqTM Ⅱ試劑盒說明書操作。三次生物學重復,所用引物見表1,采用2-ΔΔCt計算相對表達量。
2 結果與分析
2.1 AhGAIP-B基因的克隆
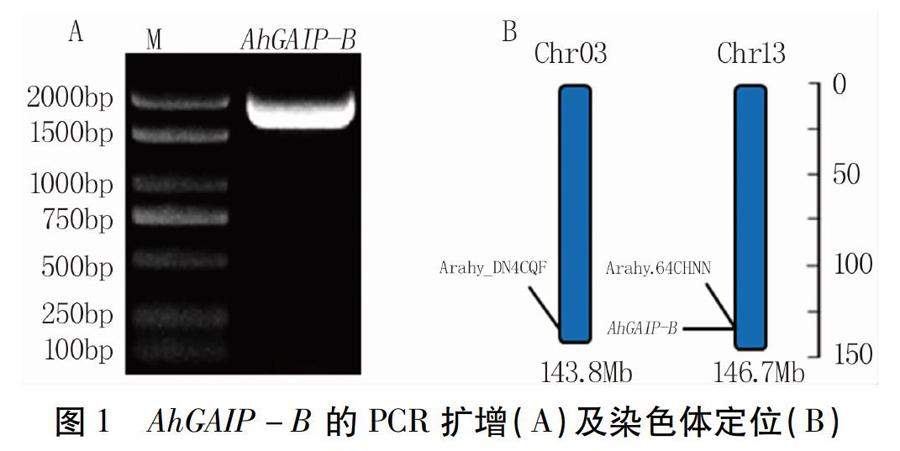
以栽培種花2014的cDNA為模板進行PCR擴增,獲得AhGAIP-B基因的cDNA序列,長1 812 bp(圖1A)。測序結果表明,該基因長度及核苷酸序列與參考基因組13號染色體上的基因Arahy.64CHNN預測序列完全一致,與同源染色體03號染色體上的參考基因Arahy.DN4CQF相似度為99.74%。
2.2 AhGAIP-B基因的生物信息學分析
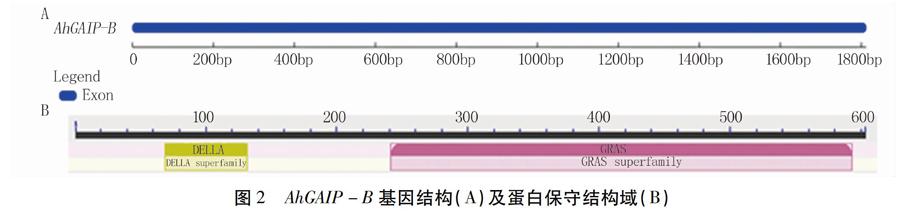
2.2.1 AhGAIP-B蛋白氨基酸序列分析 花生基因組測序結果顯示,AhGAIP-B位于花生13號染色體上(圖1B),包含一個外顯子,無內含子(圖2A),可編碼603個氨基酸。氨基酸理化性質分析結果表明,AhGAIP-B蛋白的化學分子式為C2889H4495N803O893S23,相對分子質量為65.50 kD,親水系數為-0.233,預測為親水性蛋白。同時,該蛋白無跨膜結構域,亞細胞定位在細胞核上。進一步保守域分析顯示,AhGAIP-B蛋白N端含有DELLA結構域,C端存在GRAS結構域,屬于GRAS家族基因(圖2B)。
2.2.2 AhGAIP-B蛋白結構分析 二級結構分析發現,AhGAIP-B由49.1%的α-螺旋、308%的無規則卷曲、11.6%的延伸鏈和8.5%的β-轉角共同組成(圖3A)。蛋白質三維結構分析發現AhGAIP-B蛋白質三級結構與AtGAI類似(圖3B),推測兩者可能具有相同的功能。
2.2.3 不同物種DELLA蛋白的保守結構域與系統進化分析 利用DNAMAN軟件將AhGAIP-B氨基酸序列與擬南芥(At)、大豆(Gm)、棉花(Gh)、水稻(Os)、芝麻(Si)、小麥(Ta)、油菜(Bn)及玉米(Zm)等物種的DELLA蛋白進行氨基酸多重比對,發現不同物種的DELLA蛋白在C端都含有GA信號阻遏結構域VHVID、RVER和SAW,中心區存在LZ(亮氨酸七肽重復序列)及核定位區域NLS,而N端則具有GA信號感知結構域DELLA和TVHYNP(圖4)。為進一步研究這些物種中DELLA蛋白的親緣關系,基于前述氨基酸比對結果構建系統發育進化樹。結果顯示,
前述花生等9個物種的DELLA蛋白被明確劃分為兩大類,其中,同為禾本科的單子葉植物水稻、小麥和玉米DELLA蛋白為Ⅰ大類,其余雙子葉植物則為Ⅱ大類(圖5)。Ⅱ大類里面根據遺傳距離又可細分幾個小類,如同為十字花科的擬南芥和油菜的DELLA蛋白可歸為一類,錦葵科的棉花和胡麻科芝麻的DELLA蛋白歸為一類;而同為豆科的花生和大豆的DELLA蛋白親緣關系則最近。
2.3 AhGAIP-B基因表達分析
熒光定量分析結果表明,AhGAIP-B基因在不同組織均有表達,其中莖中表達量最高,其次為根、種子、葉和花(圖6)。為進一步了解AhGAIP-B應答GA3的表達模式,對GA3處理不同時間基因的表達水平進行了分析,結果表明,AhGAIP-B基因的表達隨GA3處理時間延長先逐漸上升,在12 h時達到峰值,為處理前的26倍;此后逐漸下調,但處理48 h時仍高于處理前水平(圖7)。
3 討論與結論
本研究克隆了花生GAIP-B基因,并對其表達模式進行了分析,該基因在花生的根、莖、葉、花、種子中均有表達,與擬南芥GAI及水稻SLR[STBX]1的組織表達模式一致[14,15]。但并非所有植物的DELLA基因都是如此,擬南芥中的AtRGL1、AtRGL2和AtRGL3僅在花、萌芽的種子和角果中表達[14];蘋果GAI基因僅在幼果中表達[16];大豆GAI1a僅在葉、花和胚組織中表達[17];黃瓜GAIP基因在莖和雄花芽中表達[18],這表明DELLA基因在不同植物中可能存在不同的功能。模式植物擬南芥中的RGA和GAI結構相似,但RGA比GAI更具主導作用,敲除RGA和GAI基因后使擬南芥GA1-3缺陷型恢復為野生型或過表達GA型[19];而RGL1和RGL2共同參與種子萌發、葉片衰老和控制花形態發育等途徑[20-22]。Zhang等[23]利用黃瓜GAIP在擬南芥中異源表達,發現CsGAIP能夠恢復擬南芥rga-24/gai-t6突變體的花形態、雌蕊發育和株高,并且導致擬南芥雄蕊數量減少;此外,將蘋果RGL2a基因在擬南芥中超表達后,轉基因植株表現出類似GAI突變矮化型表型[24,25]。這些結果表明DELLA蛋白可能參與植株矮化、花發育等多個生物學功能。
本研究從花生中成功分離出一個DELLA蛋白編碼基因AhGAIP-B,其編碼產物具有典型的DELLA蛋白保守結構域,且進化上與同為豆科的大豆DELLA蛋白親緣關系最近。進一步研究發現,AhGAIP-B基因在花生多個組織部位均有表達,其中以莖中表達量最高;此外,外源GA3可以誘導AhGAIP-B基因上調表達。本研究結果有助于深入理解DELLA蛋白在花生生長發育中的調節機制,并為花生株型改良提供理論基礎。
參 考 文 獻:
[1]Itoh H,Tanaka-Ueguchi? M, Kawaide H, et al. The gene encoding tobacco gibberellin 3 β-hydroxylase is expressed at the site of GA action during stem elongation and flower organ development[J].The Plant Journal, 1999, 20: 15-24.
[2]Silverstone A L, Chang C, Krol E, et al. Developmental regulation of the gibberellin biosynthetic gene GA1 in Arabidopsis thaliana[J].The Plant Journal, 1997, 12: 9-19.
[3]Rebers M, Kaneta T, Kawaide H, et al. Regulation of gibberellin biosynthesis genes during flower and early fruit development of tomato[J].The Plant Journal, 1999, 17: 245-250.
[4]Yamaguchi S, Smith M W, Brown R G, et al. Phytochrome regulation and differential expression of gibberellin 3-hydroxylase genes in germinating Arabidopsis seeds[J].Plant Cell, 1998, 10: 2115-2126.
[5]Ikeda A, Tanaka S, Yamaguchi J, et al. Characterization of gibberellin signal transduction pathway in the slender mutant with constitutive gibberellin-response in rice[J].Sci. Rep. Fac. Agric. Meijo.Univ., 1999, 35: 15-22.
[6]Dill A, Jung H S, Sun T P.The DELLA motif is essential for gibberellin-induced degradation of RGA[J].Proceedings of the National Academy of Sciences, 2001, 98 (24): 14162-14167.
[7]吳紹華, 赫澤蕓, 張世鑫, 等.橡膠樹HbGAIP-B基因的克隆與表達分析[J].熱帶作物學報, 2016, 37(5): 881-887.
[8]King K E,Moritz T,Harberd N P. Gibberellins are not required for normal stem growth in Arabidopsis thaliana in the absence of GAI and RGA[J].Genetics, 2001, 159: 767-776.
[9]Itoh H, Ueguchi-Tanaka M, Sato Y, et al.The gibberellin signaling pathway is regulated by the appearance and disappearance of SLENDER RICE1 in nuclei[J].The Plant Cell, 2002, 14 (1): 57-70.
[10] Murase K, Hirano Y, Sun T P, et al.Gibberellin-induced DELLA recognition by the gibberellin receptor GID1[J].Nature, 2008, 456: 459-463.
[11] 吳建明, 黃杏, 丘立杭, 等. DELLA蛋白在植物中的研究進展[J].農業生物技術學報, 2016, 24(8): 1207-1215.
[12] Chen W H, Lee Y I, Yang C H. Ectopic expression of two FOREVER YOUNG FLOWER orthologues from Cattleya orchid suppresses ethylene signaling and DELLA results in delayed flower senescence/abscission and reduced flower organ elongation in Arabidopsis[J].Plant Molecular Biology Reporter, 2018, 36(5/6): 710-724.
[13] Fu X, Sudhakar D, Peng J, et al.Expression of Arabidopsis GAI in transgenic rice represses multiple gibberellin responses[J].The Plant Cell, 2001, 13(8): 1791-1802.
[14] Tyler L, Thomas S G, Hu J, et al. DELLA proteins and gibberellin-regulated seed germination and floral development in Arabidopsis[J].Plant Physiology, 2004, 135(2): 1008-1019.
[15] Ikeda A, Ueguchi-Tanaka M, Sonoda Y, et al. Slender rice, a constitutive gibberellin response mutant, is caused by a null mutation of the SLR1 gene, an ortholog of the height-regulating gene GAI/RGA/RHT/D8[J].The Plant Cell, 2001, 13(5): 999-1010.
[16] 梁美霞,祝軍,戴洪義. 柱型蘋果MdGAI基因的克隆及表達分析[J].園藝學報, 2011, 38(10): 1969-1978.
[17] 宋仙萍. 大豆DELLA基因GmGAI1α參與赤霉素信號轉導途徑的功能分析[D].哈爾濱: 東北農業大學, 2013.
[18] 張顏. 黃瓜中GA信號轉導因子CsGAIP和CsGAMYB1的克隆與功能分析[D].北京: 中國農業大學, 2014.
[19] Dill A, Sun T. Synergistic derepression of gibberellin signaling by removing RGA and GAI function in Arabidopsis thaliana[J].Genetics, 2001, 159: 777-785.
[20] Lee S C, Cheng H, King K E, et al. Gibberellin regulates Arabidopsis seed germination via RGL2, a GAI/RGA-like gene whose expression is up-regulated following imbibition[J].Genes and Development, 2002, 16(5): 646-658.
[21] Chen L G, Xiang S Y, Chen Y L, et al. Arabidopsis WRKY45 interacts with the DELLA protein RGL1 to positively regulate age-triggered leaf senescence[J].Mol. Plant, 2017, 10(9): 1174-1189.
[22] Cheng H, Qin L, Lee S, et al.Gibberellin regulates Arabidopsis floral development via suppression of DELLA protein function[J].Development, 2004, 131: 1055-1064.
[23] Zhang Y, Liu B, Yang S, et al. A cucumber DELLA homolog CsGAIP may inhibit staminate development through transcriptional repression of B class floral homeotic genes[J].PLoS ONE, 2014, 9(3): e91804.
[24] Peng J, Carol P, Richards D E, et al. The Arabidopsis GAI gene defines a signaling pathway that negatively regulates gibberellin responses[J].Genes and Development, 1997, 11(23): 3194-3205.
[25] Foster T, Kirk C, Jones W T, et al. Characterisation of the DELLA subfamily in apple (Malus×domestica Borkh.)[J].Tree Genetics & Genomes, 2007, 3(3): 187-197.
收稿日期:2019-07-24
基金項目:國家自然科學基金項目(U1704232);河南省產業技術體系(S2012-05-G03)
作者簡介:何佳霖(1995—),男,河南信陽人,碩士研究生。
通訊作者:殷冬梅(1972—),女,河南南陽人,博士,教授,主要從事花生遺傳育種工作。E-mail:yindm@126.com
