花生VAMP基因家族全基因組鑒定及表達分析
2019-12-09 01:52:35陸曉東張浩苑翠玲孫全喜閆彩霞趙小波王娟李春娟鄭奕雄單世華
山東農業科學 2019年9期
陸曉東 張浩 苑翠玲 孫全喜 閆彩霞 趙小波 王娟 李春娟 鄭奕雄 單世華


摘要:植物囊泡結合膜蛋白(VAMP)是定位在囊泡上的運輸蛋白,在植物發育以及響應生物和非生物脅迫中發揮重要作用。本研究對花生VAMP基因進行了全基因組鑒定與分析,并對它們在22個組織中的表達模式進行了分析。結果表明,二倍體野生種花生Arachis duranensis基因組有18個VAMP基因,Arachisi paensis基因組有21個,栽培種有41個,剔除假基因后花生VAMP基因家族有62個成員;表達模式分析表明AdVAMP17與AiVAMP1在大部分組織中均有表達,AdVAMP18與AiVAMP21在雌蕊中表達量最高,可能有特異性表達。本研究為進一步分析VAMP家族成員,并深入探討其在花生中的生物學功能和進化模式奠定了基礎。
關鍵詞:花生;囊泡結合膜蛋白(VAMP)基因家族;囊泡運輸;表達模式
中圖分類號:S565.2:Q781文獻標識號:A文章編號:1001-4942(2019)09-0042-08
Genome-Wide Identification and Expression Analysis of Peanut VAMP Gene Family
Lu Xiaodong1, Zhang Hao2, Yuan Cuiling2, Sun Quanxi2, Yan Caixia2, Zhao Xiaobo2, Wang Juan2, Li Chunjuan2,Zheng Yixiong1, Shan Shihua2
(1. College of Agriculture and Biology, Zhongkai University of Agriculture and Engineering, Guangzhou 510225, China;
2. Shandong Peanut Research Institute, Qingdao 266100, China)
Abstract Vesicle associated membrane protein (VAMP) is a transport protein located on vesicles of plants, which plays an important role in plant development and response to biotic and abiotic stresses. In this study, the genome-wide of peanut VAMP gene was identified and analyzed, and the expression patterns in 22 tissues were analyzed. The results showed that there were 62 members in peanut VAMP gene family, 18 of which were the Arachis duranensis genomes, 21 of which were the A. ipaensis genomes and 41 of which were the cultivars. Expression pattern analysis showed that AdVAMP17[STBZ] and AiVAMP1 were expressed in most tissues, and AdVAMP[STBX]18[STBZ] and AiVAMP[STBX]21[STBZ] had the highest expression level in pistils, which might have specific expression. These results laid a foundation for further analysis of the VAMP family members and discussing their biological functions and evolutionary patterns in peanut.
Keywords Peanut; Vesicle associated membrane protein (VAMP) gene family; Vesicle trafficking; Expression pattern
花生是我國重要的油料作物,在我國國民經濟中占有重要地位,其生長過程遭受的各種生物與非生物脅迫對花生產量和品質造成了極大的影響[1]。培育抗逆品種是提高花生產量與品質的重要手段,抗逆基因的挖掘利用是培育抗逆品種的前提,花生全基因組測序工作的完成為挖掘抗逆基因奠定了基礎[2]。
VAMP基因在植物發育以及響應生物和非生物脅迫中發揮重要作用[3]。Levine等[4]研究發現了擬南芥AtVAMP7蛋白超量表達能阻斷BAX誘導酵母產生的程序性細胞凋亡(programmed cell death,PCD),從而改善細胞膜的組分與膜的完整性,阻止細胞凋亡,對膜起修復作用。薛媛[5]研究證明了擬南芥VAMP711參與了植物激素脫落酸(ABA)介導的質膜H+-ATPase活性調節機制,在干旱脅迫下,VAMP711基因通過與質膜H+-ATPase蛋白AHA1/AHA2互作,抑制質膜H+-ATPase的活性,促進氣孔的關閉從而減少植物體內水分的流失,應對干旱脅迫。Sugano等[6]研究OsVAMP714基因敲除和過表達水稻植株并進行抗病性試驗,證明了OsVAMP714參與了稻瘟病抗性且在水稻稻瘟病抗性中具有潛在的特異性,并對水稻植株生長發育有重要作用。李華一[7]的研究證明TaVAMP714基因參與了小麥條銹菌的感病反應,且ABA對TaVAMP714基因的誘導明顯,干旱和鹽脅迫都能誘導TaVAMP714上調表達。Kim等[8]的研究發現擬南芥VAMP721/722基因是將白粉病蛋白RPW82攜帶至植物-真菌界面這一激活抗性反應分泌機制的關鍵組分,能用于細胞外周的侵入前防御和EHM的侵入性防御,對致病性白粉病真菌起防御作用。方靜[9]研究發現熱處理誘導CitVAMP71基因表達量與翻譯水平上升,椪柑果實通過增厚細胞壁響應熱脅迫,進而提高了細胞的支撐作用,利于果實加強對外界病原菌侵染及其他生物脅迫的物理屏障。Sun等[10]研究發現GsVAMP72基因的過表達不僅能通過改變離子含量和下調應激反應基因的表達而降低大豆耐鹽性,還能增加植物對ABA敏感性并改變ABA響應基因的表達水平。
目前,關于VAMP基因家族在花生中的研究尚未見報道。本研究利用生物信息學方法從全基因組水平對花生VAMP基因家族進行系統挖掘,并對該基因家族的基因結構、染色體定位、組織表達模式等進行系統分析,以期為后續研究此類基因在花生中的生物學功能奠定基礎。
1 材料與方法
1.1 序列收集
在TAIR(TAIR release 10, http://www.arabidopsis.org/)數據庫下載擬南芥VAMP家族蛋白全長序列[11],Pfam數據庫(http://pfam.Xfam.org/)下載VAMP家族蛋白結構域序列,將兩類序列在花生基因數據庫Peanutbase(https://www.peanutbase.org/)和NCBI數據庫(http://www.ncbi.nlm.nih.gov/)中進行Blastp檢索(E值≤10-15),獲得花生VAMP家族類似序列,人工去除冗余序列,作為VAMP家族的候選序列并編好基因名稱。結合Pfam數據庫中VAMP結構域保守序列和SMART在線分析軟件(http://smart.embl-eidelberg.de/)對候選序列進行VAMP結構域分析,剔除不含有該結構域的蛋白質序列,得到花生VAMP基因家族全體成員序列。
1.2 系統發育進化樹構建與染色體定位分析
根據獲得的擬南芥與花生VAMP家族蛋白全長序列數據信息,將其氨基酸序列在MEGA 7.0上進行序列比對,采用非加權組平均法(unweighted pair-group method with arithmetic means, UPGMA)對比后獲得的序列構建系統發育進化樹,采用Bootstraping法對進化樹進行評估與分組。校驗參數: p-distance重復1 000次。
利用ExPASy-ProtParam(http://web.expasy.org/protparam/)對所得花生VAMP基因家族蛋白序列的分子量及等電點等理化性質進行分析。
從花生基因組中的gff3文件中提取VAMP基因家族的染色體起始和結束位置,利用MapInspect軟件進行染色體物理分布圖的繪制,獲得VAMP成員的染色體物理定位信息。
1.3 花生VAMP家族基因結構及保守基序分析
由Gene Structure Display Server 2.0 (http://gsds.cbi.pku.edu.cn/)在線網站提供基因結構預測的可視化工具[12]。將去除假基因的花生VAMP基因家族的CDS序列及基因組序列在GSDS程序中得到其基因結構示意圖,在基因結構圖上標注特定區域,得到VAMP基因家族基因結構圖。
利用MEME 4.12(http://meme-suite.org/tools/meme)[13]在線網站對VAMP家族基因的保守基序進行分析。
1.4 花生VAMP基因表達量分析
利用Heml軟件分析花生VAMP基因在花生22個組織中的表達情況。
2 結果與分析
2.1 花生VAMP基因家族鑒定與理化性質分析
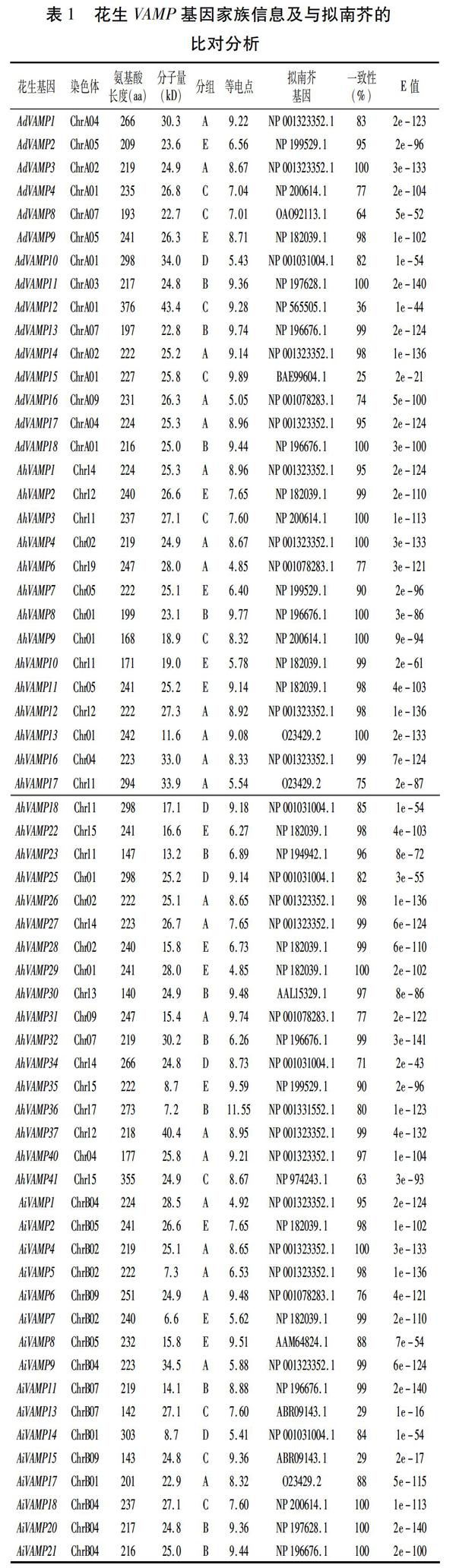
從兩個二倍體野生種花生A.duranensis和A.ipaensis基因組中分別得到18個和21個VAMP基因,在普通花生栽培種中得到41個VAMP基因家族成員,分別命名為AdVAMP1~18、AiVAMP1~21、AhVAMP1~41(表1)。利用Pfam、SMART等軟件檢驗這80個基因家族成員,發現其中有18個成員具有不完整的結構域,可能為假基因,剔除這18個成員,將擁有完整結構域的62個花生VAMP基因家族成員構建系統進化樹,根據其拓撲結構,將花生VAMP基因家族分為5組,分別命名為group A、B、C、D、E。
在進化樹中我們發現group A擁有最多的花生VAMP基因家族成員,為23個;其次是group E(13個)、group B(11個)、group C(10個)、group D(5個)(圖1)。由進化樹分析可知,花生VAMP蛋白家族形成兩大分支,共有24對旁系同源基因,其中有5對旁系同源對bootstrap值小于70,其他19對旁系同源對bootstrap值在70以上,其中有15對旁系同源對bootstrap值大于90,因此我們認為花生VAMP蛋白在進化上親緣關系較為接近。
理化性質分析發現,VAMP基因家族成員的氨基酸長度差異較小,其長度在140(AhVAMP30)~376 aa(AdVAMP12)之間,且大部分基因家族成員氨基酸長度在200~300 aa之間;花生VAMP蛋白分子量在6.6 kD(AiVAMP7)與43.4 kD(AdVAMP12)之間;等電點分布在4.85(AhVAMP6、AhVAMP29)與11.55(AhVAMP36)之間,有17個VAMP蛋白偏酸性,等電點小于7.0,其他45個VAMP蛋白呈中性或偏堿性。為了探究花生VAMP家族基因的生物學功能,將其與模式植物擬南芥進行了比對分析(表1)。結果表明,花生VAMP基因家族與擬南芥對應的基因之間有較好的同源關系(E-value<3.00e-141)。該結果為研究花生VAMP基因家族的功能提供了一定的借鑒。
2.2 花生VAMP基因家族的染色體分布
染色體定位分析有助于更直觀地了解花生VAMP基因家族在染色體中的分布情況,由圖2~圖4[JP]可知VAMP基因家族成員在染色體上并沒有均勻分布。在A.duranensis基因組染色體A06與A08上并無分布,而在染色體A01上分布最多,高達6個VAMP基因(圖2);在A.ipaensis基因組染色體B10上無分布,而在染色體B01及B02上均分布有4個成員(圖3);在栽培種染色體06、染色體08、染色體10、染色體16、染色體20上無分布,而在染色體01與染色體11上均分布有6個成員(圖4)。由此可見,花生VAMP基因家族成員在染色體上的位置分布并不均勻,且對于花生VAMP基因家族40條染色體的觀察發現多數基因家族成員分布在染色體的兩端,而靠近著絲粒部分的基因較少。
2.3 VAMP家族基因結構及保守性分析
使用MEME軟件對花生62個VAMP基因家族成員所編碼的蛋白進行保守基序分析,獲得6個motif的信息,分別命名為motif 1~motif 6。預測結果如圖5所示,有23個基因成員均含有motif 1、motif 2、motif 4、motif 5,有16個基因家族成員僅含有motif 3和motif 6兩個結構域,motif 1、motif 2、motif 4在VAMP基因家族中分別存在36、38、35個,由此,我們認為VAMP基因家族有較為保守的結構。此外,我們觀察到group A與group B大部分基因編碼的蛋白均含有motif 1、motif 2與motif 4,而group C基因成員編碼的蛋白則含有motif 1、motif 2、motif 4與motif 5。這說明group A與group B功能可能較為類似,group C與group A及group B親緣關系較近,基因可能有較為類似的功能。而group D與group E大部分基因編碼的蛋白僅含有motif 3與motif 6,說明這兩組基因家族成員與group A、group B、group C親緣關系較遠,基因編碼蛋白功能可能差異較大。我們還分析了62個花生VAMP基因家族的結構,發現5組基因家族成員之間結構差異較大,而組內的結構相對接近(圖5)。大部分基因家族成員都由外顯子(exon)、非編碼區(UTR)和內含子(intron)組成,僅有AiVAMP11、AiVAMP20、AhVAMP3、AhVAMP8、AhVAMP9、AhVAMP23這6個基因家族成員僅由內含子和外顯子組成。我們注意到,在VAMP基因家族中,5組基因家族的內含子外顯子結構分化嚴重,變化較大,其中group C的平均長度在五組里面最短,group A平均長度最長。
2.4 VAMP基因家族表達模式分析
基因表達模式分析有利于我們研究VAMP基因家族的功能。利用苗期葉片等22個組織的轉錄組數據,對花生A.duranensis與A.ipaensis基因組共31個成員在花生22個組織中的表達模式進行分析,由圖6可知,大部分VAMP基因家族成員在22個組織中有不同程度的表達,且在不同組織或器官中,不同基因家族成員的表達模式存在明顯差異。在本聚類圖中,用藍、綠、黃、紅深度表示基因表達的強弱情況,顏色越接近紅色表達情況越強,黑色則表示該成員在此組織中未見表達。AdVAMP17與AiVAMP1表達量最高,整體呈紅色,說明這兩個蛋白在花生22個組織中均有表達,且表達量是所有VAMP基因家族最高的;而AdVAMP18、AiVAMP21、AdVAMP14、AiVAMP5、AiVAMP13、AiVAMP15與AdVAMP8共7個基因家族成員在多數組織中未見表達或表達量較低,但在雌蕊(pistils)中,AdVAMP18與AiVAMP21兩個基因成員的表達量是所有VAMP基因家族成員中最高的,表明在雌蕊中AdVAMP18與AiVAMP21可能有特異性表達。
3 討論與結論
花生是我國重要的油料作物,生長過程中遇到的脅迫問題復雜而多樣,挖掘和利用抗逆基因具有重要意義。眾多學者的研究表明VAMP蛋白介導的囊泡運輸在植物對抗逆境脅迫中發揮重要作用,因此對于VAMP基因家族的研究不容忽視。近年來,關于植物VAMP基因家族克隆及功能方面的研究集中在擬南芥、水稻、麥類等植物,主要參與抗病、干旱、鹽脅迫等逆境反應[4-10,14,15]。因此,我們預測花生VAMP基因家族可能參與抗逆反應。
本研究基于花生全基因組和22個組織轉錄組測序數據,首次對花生VAMP基因家族進行了系統發育樹、保守域、保守基序、染色體定位、22個組織表達模式的分析,共鑒定到62個花生VAMP蛋白,并根據進化樹與基因結構將其分為5組。花生VAMP基因家族的保守基序非常明顯地將其分為了兩組,我們推測擁有motif 1、motif 2、motif 4的A、B、C組基因功能可能較為接近,擁有motif 3和motif 6的D、E組基因與前者可能有不一樣的功能。近年來關于植物功能基因家族的研究表明,在水稻、擬南芥、陸地棉等植物中,擁有相同保守基序或功能域的氨基酸序列在系統發育樹相同的組內是高度保守的,這些同一組內含有相同基序的蛋白或具有相似功能[16]。
基因的表達模式分析可以為未知基因功能的預測做一定的貢獻。我們分析了花生VAMP基因家族在22個組織中的表達模式,發現AdVAMP18和AiVAMP21在雌蕊中表達量最高,雄蕊次之,而這兩個基因在其他組織中幾乎不表達或者表達量很低,而AdVAMP14和AiVAMP5也在雌蕊中有較高表達,因此我們推測AdVAMP18、AiVAMP21、AdVAMP14和AiVAMP5在花及種子形成過程中可能發揮著重要作用。而AdVAMP8基因在花生種子中不表達,暗示其啟動子可能是個非種子表達啟動子,可以用于花生遺傳轉化,使轉化基因在花生種子中不表達,避免人們對轉基因的恐慌。同時,我們發現在花生VAMP基因家族染色體定位中,A.duranensis基因組的AdVAMP16基因與A.ipaensis基因組的AiVAMP6均在09號染色體,四倍體栽培種花生的AhVAMP31、AhVAMP6在09號及19號染色體上具有相似的位置,結合表達聚類圖中AdVAMP16基因與AiVAMP6基因在花生22個組織中相似的表達模式及在基因結構分析中相似的內含子、外顯子結構,我們認為這四個基因可能為同源基因,具有相同的功能。同理,A.duranensis基因組的AdVAMP18基因與A.ipaensis基因組的AiVAMP18基因均在1號染色體,四倍體栽培種花生的AhVAMP8、AhVAMP18等在1號染色體與11號染色體上也具有類似位置,也可能為具有相同功能的同源基因。具體基因功能的探索還需要進一步試驗驗證。
本研究首次系統分析了VAMP蛋白在花生中的情況,包括染色體定位、基因結構、保守結構域、表達模式等,為下一步深入研究此類蛋白提供了參考。
參 考 文 獻:
[1]Sharma S, Pandey M K, Sudini H K, et al. Harnessing genetic diversity of wild Arachis species for genetic enhancement of cultivated peanut[J].Crop Science, 2017, 57(3): 1121-1131.
[2]Zhuang W, Chen H, Yang M, et al. The genome of cultivated peanut provides insight into legume karyotypes, polyploid evolution and crop domestication[J].Nature Genetics, 2019, 51(5): 865.
[3]Fujii H, Zhu J K. Osmotic stress signaling via protein kinases[J].Cellular and Molecular Life Sciences, 2012, 69(19):3165-3173.
[4]Levine A, Belenghi B, Damari-Weisler H, et al.Vesicle-associated membrane protein of Arabidopsis suppresses Bax-induced apoptosis in yeast downstream of oxidative burst[J].Journal of Biological Chemistry, 2001, 276(49):46284-46289.
[5]薛媛. 擬南芥SNARE蛋白VAMP711通過調控質膜H+-ATPase響應干旱脅迫的機制研究[D].北京:中國農業大學,2019.
[6]Sugano S, Hayashi N, Kawagoe Y, et al. Rice OsVAMP714, a membrane-trafficking protein localized to the chloroplast and vacuolar membrane, is involved in resistance to rice blast disease[J].Plant Molecular Biology, 2016, 91(1/2):81-95.
[7]李華一. 小麥SYP71和VAMP714基因在小麥與條銹菌互作中的功能分析[D].楊凌:西北農林科技大學,2014.
[8]Kim H, OConnell R, Maekawa-Yoshikawa M, et al. The powdery mildew resistance protein RPW8.2 is carried on VAMP721/722 vesicles to the extrahaustorial membrane of haustorial complexes[J].The Plant Journal, 2014, 79(5):835-847.
[9]方靜. 熱處理誘導椪柑果實抗逆相關蛋白VAMP表達的初步研究[D].武漢:華中農業大學,2011.
[10]Sun X, Ji W, Ding X, et al. GsVAMP72, a novel Glycine soja R-SNARE protein, is involved in regulating plant salt tolerance and ABA sensitivity[J].Plant Cell Tissue and Organ Culture, 2013, 113(2):199-215.
[11]Poole R L. The TAIR database[M].Plant Bioinformatics, 2005.
[12]郭安源, 朱其慧, 陳新, 等. GSDS:基因結構顯示系統[J].遺傳, 2007, 29(8):1023-1026.
[13] Bailey T L, Johnson J, Grant C E, et al. The MEME suite[J].Nucleic Acids Research, 2015, 43(W1):W39.
[14]豆獻英. 稻瘟病菌Mgvam7基因的克隆與功能分析[D].南京:南京農業大學,2008.
[15]裴洪翠. 麥類作物抗銹病相關蛋白互作網絡的研究[D].泰安:山東農業大學,2014.
[16] Tang Y, Qin S, Guo Y, et al. Genome-wide analysis of the AP2/ERF gene family in physic nut and overexpression of the JcERF011 gene in rice increased its sensitivity to salinity stress[J].PLoS ONE, 2016, 11(3): e0150879.
收稿日期:2019-07-24
基金項目:泰山學者特聘專家(ts201712080);山東省農業良種工程項目(2017LZN033,2017LZGC003);山東省現代農業產業技術體系(SDAIT-04-02);國家自然科學基金青年科學基金項目(31601336);廣東省農業農村廳科技項目(2019KJ136);山東省重點研發計劃項目(2016GGH4518);青島市科技惠民計劃項目(17-3-3-49-nsh)
作者簡介:陸曉東(1994—),女,在讀碩士研究生,研究方向為花生遺傳育種。E-mail:xdongdonglu@163.com
通訊作者:鄭奕雄(1963—),男,教授,主要從事花生育種與栽培研究。E-mail: gdsscqs@163.com
單世華(1971—),男,研究員,主要從事花生種質創新及利用研究。E-mail:shansh1971@163.com
