深圳壩光銀葉樹種群生命表及生存力分析
2019-12-19 02:13:14孫紅斌肖石紅蔡堅高常軍易小青吳琰魏龍王佐霖
熱帶作物學報 2019年11期
孫紅斌 肖石紅 蔡堅 高常軍 易小青 吳琰 魏龍 王佐霖
摘 ?要 ?以種群生命表和生存力分析理論為基礎,應用徑級法及勻滑技術,編制深圳壩光古銀葉樹(Heritiera littoralis)種群特定時間生命表,繪制其死亡率曲線、消失率曲線、存活曲線和生存率函數曲線,同時分析種群數量動態變化。結果表明:(1)壩光銀葉樹種群以幼苗和幼樹個體數量居多,老齡個體數量較少,表明現階段銀葉樹整體為增長型種群,考慮未來潛在干擾影響時種群仍處于穩定狀態;(2)銀葉樹種群存活曲線趨于Deevey-Ⅲ型,死亡率和消失率均在第Ⅰ齡級和第Ⅲ齡級出現2個峰值,曲線的變化趨勢基本一致;(3)種群生存分析結果顯示,生存率函數曲線呈單調上升趨勢,累計死亡率函數曲線單調下降,且前期上升或下降的幅度高于后期,種群死亡密度函數曲線較為平緩,危險率函數從第Ⅲ到第Ⅻ齡級隨齡級的增大不斷增大,死亡密度和危險率函數在第Ⅱ齡級最大。
關鍵詞 ?銀葉樹;種群生命表;種群動態;存活曲線;生存分析
中圖分類號 ?S718.5 ? ? ?文獻標識碼 ?A
Abstract ?The specific life table of Heritiera littoralis population in Baguang, Shenzhen was compiled based on the population life table and the survival analysis using the DBH (diameter at breast height) class method and smoothing out technique. The mortality rate curve, vanishing rate curve, survival rate curve and survival function curve were calculated, and the population dynamics were also analyzed. In H. littoralis population, the seedlings and young trees were in larger quantity, while the aged ones were fewer, indicating that the population was overall a growing population. The population was stable even taking potential future disturbance into account. The survival curve was more of Deevey-Ⅲ type. The mortality and vanishing curves both exhibited the same trend, existing two peaks in the lifespan. One was in the 1st age and the other was in the 3rd. Analysis of four survival functions of H. littoralis population showed that the survival function curve increased all long, while the cumulative mortality function curve decreased all the time. The increasing or decreasing range was higher at young stage than that at middle or mature stages. The death density function curve was relatively stable. The hazard rate function curve was increasing from the 3rd age to the 12th age, with the growth of age class. The death density and hazard rate function curves were highest during the 2nd age class.
Keywords ?Heritiera littoralis; population life table; population dynamics; survivorship curve; survival analysis
DOI ?10.3969/j.issn.1000-2561.2019.11.009
種群作為物種在自然界中存在的基本單位,是生物群落的基本組成單元,種群結構和動態是生態學研究的核心問題之一[1]。種群生命表是統計種群數量動態的有效方法之一,根據種群生命表所提供的數據,不僅能獲取種群的存活率、死亡率和消失率等種群特征重要參數,還能重建種群過去的干擾歷史,探索種群的進化演替歷程,預測種群未來生命趨勢及生存可能性,進而反映種群對環境的適應力[2-3]。植物種群動態是植物個體生存能力與外界環境因素相互作用的結果,對種群數量或大小在時間和空間上的變化規律有較好的解釋作用[4]。生存分析的4個函數可輔助種群生命表分析,能更好地闡明種群的生存規律[5]。
銀葉樹(Heritiera littoralis)是梧桐科(Sterculiaceae)屬于熱帶、亞熱帶海岸半紅樹植物,多分布于高潮線附近的海灘內緣、大潮或特大潮才能淹沒的灘地或海岸陸地,屬于典型的海陸兩棲的紅樹植物[6]。深圳壩光銀葉樹濕地園銀葉樹林歷史悠久,保存了種類豐富、數量眾多的古樹,是目前全國乃至全世界發現樹齡最長的天然古銀葉樹群落,具有重要的科研和經濟價值,也是珍貴的旅游資源[7]。國內外對銀葉樹的群落結構[8]、生物學特性[9]、藥用價值[10]、果實特征及萌芽技術[11]、鹽度和淹水對其苗木的影響[12-13]、遺傳多樣性[14-16]等方面做了相關研究,但對種群結構和生存力分析方面的研究較為缺乏。
本研究以種群生態學理論為基礎,對深圳壩光銀葉樹種群的數量特征進行初步分析,編制種群生命表,探討種群動態和生存狀態,以期為壩光古銀葉樹種群和群落的更新、生態系統的恢復和資源合理利用等提供科學依據。
1 ?材料與方法
1.1 ?研究區概況
壩光銀葉樹濕地園位于廣東省深圳市大鵬新區大鵬半島北端,地理位置:2237N~2239N,11430E~11432E,氣候屬亞熱帶季風氣候。年均氣溫22.1 ℃,歷年極端最高溫度36.6 ℃,極端最低溫度1.4 ℃,年平均相對濕度79%,年平均降水量1800.4 mm。研究區內銀葉樹林總面積約7.5 hm2,其中銀葉樹分布面積約0.7 hm2,現已定為國家珍稀植物(即銀葉樹)群落小區,該地銀葉樹林主要有2片:一片為小丘地(陸生環境)及內海灣灘地(海生環境),面積約為2500 m2,形成以銀葉樹為主的森林群落,內海灣灘地面積約2000 m2,銀葉樹常年被海水浸泡,周圍零星生長有少量秋茄(Kandelia candel)、鹵蕨(Acrostichum aureum)、老鼠簕(Acanthus ilicifolius)和木欖(Bruguiera gymnorhiza)等;另一片則在鄰海面沿海岸線間斷分布(海生環境),總長約200 m,面積約為1500 m2。
1.2 ?野外調查方法
于2017年10月全面調查深圳壩光銀葉樹濕地園內的銀葉樹個體,統計所有胸徑≥2.5 cm個體的樹高、胸徑、冠幅等以及所有胸徑<2.5 cm幼樹、幼苗的株數、高度等指標。
1.3 ?銀葉樹年齡的劃分
自然生長的銀葉樹個體的標準年齡較難準確確定,且其作為保護樹種,不能通過鉆取木芯的方式來確定其個體年齡,因此以銀葉樹徑級結構代替種群年齡結構分析種群動態,即根據銀葉樹胸徑的大小來劃分年齡結構級,按照每個年齡級5 cm的原則,將銀葉樹個體劃分為13個徑級:D<5 cm為Ⅰ級,5 cm≤D<10 cm為Ⅱ級,15 cm≤D<20 cm為Ⅲ級,以此類推,D≥60 cm為ⅩⅢ級。統計各齡級個體數,以獲取的數據編制銀葉樹種群靜態生命表,分析銀葉樹種群生命過程。
1.4 ?靜態生命表的編制
根據壩光銀葉樹濕地園銀葉樹種群不同年齡個體的分布情況編制靜態生命表。種群靜態生命表分析參照文獻[17-18]方法進行。由于研究區內銀葉樹種群為野生分布的自然林,調查的數據會出現后一個齡級高于前一個齡級的存活個體的情況,即死亡率為負值,為避免此情況的出現,采用勻滑技術[19]對調查得到的銀葉樹種群數據進行處理,編制銀葉樹的靜態生命表。
1.5 ?種群年齡結構數量動態變化
采用種群數量變化動態指數對不同齡級銀葉樹數量動態進行分析,計算自然狀態和考慮外部干擾時種群結構數量動態指數[20]。
自然狀態下種群數量變化動態指數Vpi:
考慮外部干擾時種群數量變化動態指數 :
式中,Vn為種群從n到n+1齡級的種群個體數量動態;Sn為第n齡級的種群個體數;Sn+1為第n+1齡級的種群個體數;max(Sn,Sn+1)和min(S1,S2,…,Sn)分別為數列極大值和極小值。Vpi和 大于0時,表示種群增長;小于0時,表示種群衰退;等于0時,表示種群穩定。
1.6 ?生存分析
引入生存分析中的種群生存率函數S(i)、累計死亡率函數F(i)、死亡密度函數f(ti)和危險率函數λ(ti)4個函數[2, 21],分析銀葉樹種群動態,闡明其生存規律,其計算公式如下:
式中,S(i)和hi分別為存活率齡級寬度。
2 ?結果與分析
2.1 ?銀葉樹種群靜態生命表
由銀葉樹種群靜態生命表(表1)可以看出,銀葉樹種群數量隨著齡級的增大波動較大。總體來說,銀葉樹幼苗、幼樹個體數量較豐富,個體數量高峰期出現在第Ⅰ~Ⅲ齡級。死亡率和消失率均在第Ⅰ和第Ⅲ齡級出現小高峰,出現小高峰的原因可能是由于銀葉樹幼苗和幼樹個體數量較多,而此階段生長和競爭能力相對較弱,無法適應強烈的環境而死亡;Ⅳ齡級時種群的死亡率和消失率最低。從第Ⅳ齡級開始,銀葉樹種群期望壽命值隨著齡級的增加逐漸呈降低趨勢,在第ⅩⅢ齡級時種群期望壽命僅為0.500,說明銀葉樹種群生命期望在成年階段較高。銀葉樹種群存活率在第Ⅰ齡級最低,僅為19.70%。
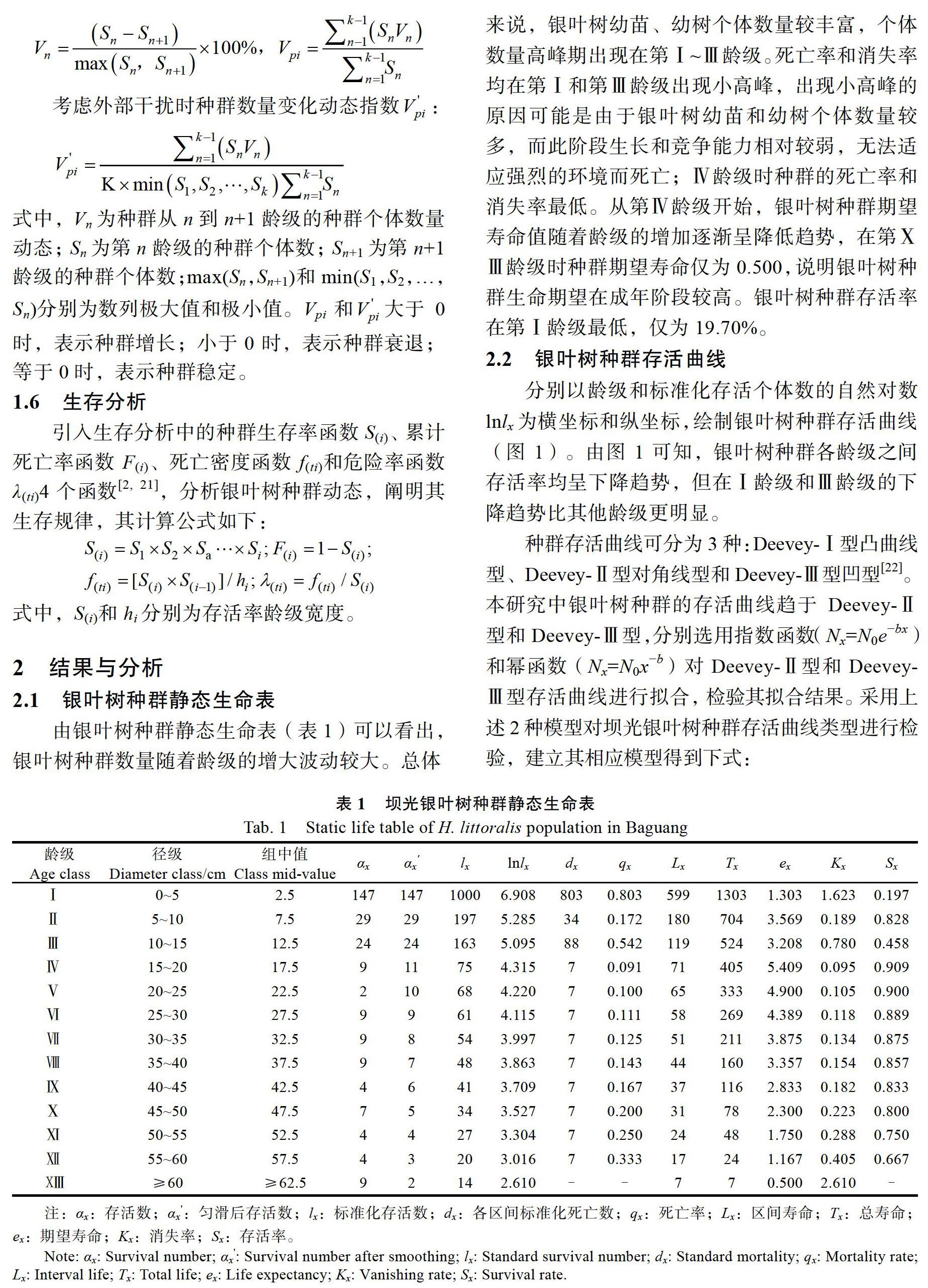
2.2 ?銀葉樹種群存活曲線
分別以齡級和標準化存活個體數的自然對數lnlx為橫坐標和縱坐標,繪制銀葉樹種群存活曲線(圖1)。由圖1可知,銀葉樹種群各齡級之間存活率均呈下降趨勢,但在Ⅰ齡級和Ⅲ齡級的下降趨勢比其他齡級更明顯。
種群存活曲線可分為3種:Deevey-Ⅰ型凸曲線型、Deevey-Ⅱ型對角線型和Deevey-Ⅲ型凹型[22]。本研究中銀葉樹種群的存活曲線趨于Deevey-Ⅱ型和Deevey-Ⅲ型,分別選用指數函數(Nx=N0e?bx)和冪函數(Nx=N0x?b)對Deevey-Ⅱ型和Deevey-Ⅲ型存活曲線進行擬合,檢驗其擬合結果。采用上述2種模型對壩光銀葉樹種群存活曲線類型進行檢驗,建立其相應模型得到下式:
從以上結果可以看出,2種模型均達到極顯著水平,相關性均較高,但冪函數擬合方程的相關系數比指數函數更高,因此本研究中銀葉樹種群存活曲線更趨向于Deevey-Ⅲ型曲線,即銀葉樹早期個體數量多,死亡量、死亡率和消失率高,生長到成熟時期時,種群內個體趨于穩定。
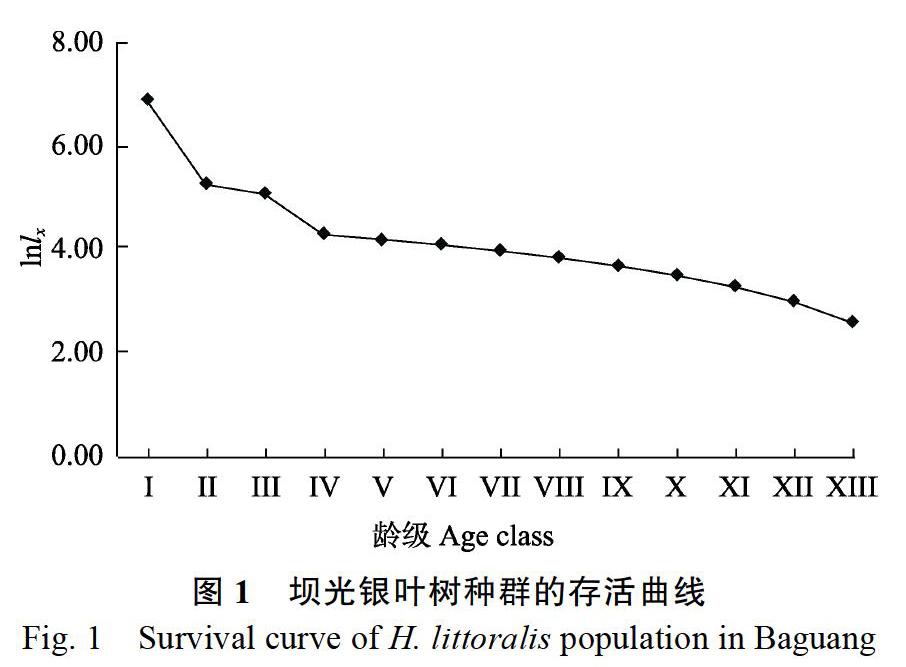
2.3 ?銀葉樹種群死亡率和消失率曲線
以齡級為橫坐標,死亡率qx或消失率Kx為縱坐標,繪制銀葉樹種群死亡率和消失率曲線(圖2)。如圖2所示,銀葉樹群落死亡率和消失率曲線均在第Ⅰ齡級和第Ⅲ齡級出現2個峰值,其變化趨勢基本一致,死亡率分別為80.30%和54.20%。整體而言,銀葉樹種群的死亡率和消失率曲線隨著齡級的增加呈現先減小后增加的趨勢。
2.4 ?銀葉樹種群年齡結構動態
深圳壩光銀葉樹種群結構動態的量化分析顯示,從第Ⅰ~XⅢ齡級的相鄰間個體數量變化動態指數(Vn)分別為80.27%、17.24%、62.50%、77.78%、?77.78%、0.00%、0.00%、55.56%、?42.86%、42.86%、0.00%和?55.56%。銀葉樹種群年齡結構的動態指數(Vpi)為57.40%,大于0,說明該種群為增長型種群。在考慮種群受到潛在外部環境干擾情況下,銀葉樹種群齡級結構的動態指數(V 'pi)為2.21%,接近0,即考慮潛在外部干擾情況時種群仍保持明顯的穩定結構。
2.5 ?銀葉樹種群生存分析
圖3和圖4為銀葉樹種群生存率函數曲線、累計死亡率函數曲線、死亡密度函數曲線和危險率函數曲線。銀葉樹種群的生存率隨齡級的增大大致呈遞減趨勢,相應的累計死亡率則呈遞增趨勢;生存率和累計死亡率曲線在第Ⅰ~Ⅲ齡級變化幅度均較大,之后變化緩慢。種群死亡密度函數曲線比較平緩,第Ⅰ~Ⅲ齡級死亡密度較大,而危險率函數則在第Ⅻ齡級最大,從第Ⅲ到第Ⅻ齡級隨著年齡的增大危險率呈增大的趨勢。
3 ?討論
本研究通過對深圳壩光銀葉樹的野外調查,用徑級法分析了銀葉樹種群的數量特征與動態。種群年齡結構、靜態生命表、存活曲線、死亡率曲線和消失率曲線能反映其生物學特性與環境因素相互作用關系[17]。深圳壩光銀葉樹種群整體上以幼苗和幼樹為主,定量分析種群數量變化動態指數>0,說明該種群為增長型種群,但考慮未來潛在干擾影響時數值接近0,說明種群仍保持穩定,這與陳曉霞等[8]通過銀葉樹群落高度級和立木級分布數據結果推測出該群落處于穩定生長型階段的結果趨于一致,盡管銀葉樹種群在一定時間內保持相對穩定,但隨著時間的推移,種群將趨于老化。
銀葉樹種群生命表顯示,種群的存活曲線趨向于Deevey-Ⅲ型,為衰退型種群。銀葉樹種群數量的動態變化趨勢能通過存活曲線、死亡率曲線、消失率曲線以及4個生存函數體現,種群具有前期數量下降較快,中后期數量相對穩定的特點。在整個生命周期內,銀葉樹出現2次死亡高峰,分別發生在第Ⅰ和第Ⅲ齡級,這主要是由于銀葉樹種群在幼苗和幼樹期數量較大,對資源的需求也較大,但生境中可利用的資源有限,種內競爭加劇,使得其死亡率增加,這也表明銀葉樹一旦在幼苗、幼樹經過環境的篩選后,大多數個體能存活到正常壽命。
種群在不同生長期的個體數、生存分析以及數量動態往往是由植物在生長周期各個階段所對應的性狀表現和環境因子共同作用決定的,體現了種群動態變化過程和群落的發展趨勢[23]。通過對深圳壩光銀葉樹種群生命表、種群動態和生存力的綜合分析表明,該種群在一定程度上具有相對穩定性,但由于幼苗、幼樹時期死亡率和消失率較高,加速了其走向衰退的風險,因此應及時加以有效的保護,為幼苗和幼樹的生長提供優越的存活空間。此外,可適當收集壩光銀葉樹種子,采用人工育種方式對其進行撫育,擴大分布區域,最終讓其擁有更穩定、平衡的發展條件。
參考文獻
[1] Gurevitch J, Scheiner S M, Fox G A. The ecology of plants[M]. Sunderland Massachusetts, USA:Sinauer Assocoates Inc, 2002.
[2] 張欽弟, 張金屯, 蘇日古嘎, 等. 龐泉溝自然保護區華北落葉松種群生命表與譜分析[J]. 應用與環境生物學報, 2010, 16(1): 1-6.
[3] 劉 ?丹, 劉士玲, 郭忠玲, 等. 水冬瓜赤楊種群生命表與時間序列分析[J]. 北京林業大學學報, 2017, 39(10): 62-69.
[4] 黨海山, 張燕君, 張克榮, 等. 秦嶺巴山冷杉(Abies fargesii)種群結構與動態[J]. 生態學雜志, 2009, 28(8): 1456-1461.
[5] 楊鳳翔, 王順慶, 徐海根, 等. 生存分析理論及其在研究生命表中的應用[J]. 生態學報, 1991, 11(2): 153-158.
[6] 王伯蓀, 張煒銀, 昝啟杰, 等. 紅樹植物之詮釋[J]. 中山大學學報(自然科學版), 2003, 42(3): 42-46.
[7] 林中大, 唐光大. 深圳市龍崗區古樹現狀調查研究[J]. 廣東林業科技, 2012, 28(2): 63-68.
[8] 陳曉霞, 李 ?瑜, 茹正忠, 等. 深圳壩光銀葉樹群落結構與多樣性[J]. 生態學雜志, 2015, 34(6): 1487-1498.
[9] 簡曙光, 韋 ?強, 唐 ?恬, 等. 深圳鹽灶銀葉樹種群的生物學特性研究[J]. 華南農業大學學報, 2005, 26(4): 84-87, 91.
[10] Christopher R, Nyandoro S S, Chacha M, et al. A new cinnamoylglycoflavonoid, antimycobacterial and antioxidant constituents from Heritiera littoralis leaf extracts[J]. Natural Product Research, 2014, 28(6): 351-358.
[11] Ye Y, Lu C Y, Wong Y, et al. Diaspore traits and inter-tidal zonation of non-viviparous mangrove species[J]. Acta Botanica Sinica, 2004, 46(8): 896-906.
[12] Paliyavuth C, Clough B, Patanaponpaiboon P. Salt uptake and shoot water relations in mangroves[J]. Aquatic Botany, 2004, 78(4): 349-360.
[13] Mangora M M, Mtolera M, Bj?rk M. Effects of waterlogging, salinity and light on the productivity of Bruguiera gymnorrhiza and Heritiera littoralis seedlings[J]. African Journal of Marine Science, 2017, 39(2): 167-174.
[14] Jian S G, Tang T, Zhong Y, et al. Conservation genetics of Heritiera littoralis (Sterculiaceae), a threatened mangrove in China, based on AFLP and ISSR markers[J]. Biochemical Systematics and Ecology, 2011, 38(5): 924-930.
[15] Das A B, Mukherjee A K, Das P. Molecular phylogeny of Heritiera, Aiton (Sterculiaceae), a tree mangrove: variations in RAPD markers and nuclear DNA content[J]. Botanical Journal of the Linnean Society, 2001, 136(2): 221-229.
[16] Hu W M, Wang S W, Jian S G, et al. Discovery and comparative assessment of SNPs and SSRs for genetic diversity analysis of wild Heritiera littoralis, Dryand (Sterculiaceae), an endangered mangrove tree species[J]. Conservation Genetics Resources, 2015, 7(3): 663-668.
[17] 李 ?豪, 張欽弟, 苗艷明, 等. 歷山自然保護區禿山白樹天然種群生命表[J]. 生態學雜志, 2013, 32(1): 52-58.
[18] 趙志剛, 郭俊杰, 曾 ?杰, 等. 廣西大明山格木種群的空間分布格局與數量動態特征[J]. 林業科學, 2014, 50(10): 1-7.
[19] 江 ?洪. 云杉種群生態學[M]. 北京: 中國林業出版社, 1992.
[20] 劉國軍, 張希明, 朱軍濤, 等. 準噶爾盆地東南緣梭梭種群結構與動態研究[J]. 西北植物學報, 2011, 31(6): 1250-1256.
[21] 蔡新斌, 吳俊俠. 甘家湖自然保護區白梭梭種群特征與動態分析[J]. 干旱區資源與環境, 2016, 30(7): 90-94.
[22] 鐘軍弟, 成夏嵐, 莫雨杏, 等. 雷州九龍山紅樹林國家濕地公園玉蕊種群動態[J]. 濕地科學, 2018, 16(2): 231-237.
[23] 張滋庭, 閆 ?明, 張滋芳. 山西霍山華北落葉松種群生態特征研究[J]. 山西師范大學學報(自然科學版), 2018, 32(2): 52-60.
