免疫細胞與外泌體相互作用機制的研究進展①
2019-12-27 06:18:20劉滿宇張文慧張林波
中國免疫學雜志 2019年22期
劉滿宇 付 璐 張文慧 張林波
(吉林農業大學生命科學學院,長春 130118)
外泌體作為一種功能廣泛的胞內囊泡,與免疫細胞的多種功能有著密切聯系。但外泌體在免疫細胞間及免疫細胞與其他細胞間如何發揮作用,以及外泌體的作用與機體免疫應答之間存在何種聯系,目前尚未完全闡明。探究外泌體對免疫細胞的影響及在免疫過程中的作用,對于進一步揭示免疫系統功能及其運作方式具有重要意義。
1 外泌體的形成機制及主要作用
外泌體(Exosome)是一種由磷脂雙分子膜與包裹在膜中的蛋白質、核酸、脂類、糖類等物質構成的囊泡樣結構,直徑在30~100 nm之間。20世紀80年代首次被發現于羊網織紅細胞的培養液中。一般認為,外泌體起源于細胞內的多囊體(Multivesicular body,MVB),在ESCRT復合物、脂質和四跨膜蛋白超家族等多種分子的參與下成熟,并在RAB蛋白家族的參與下與質膜融合,進而釋放到細胞外環境中(圖1),這一融合過程也可能與SNAREs蛋白有關。

圖1 細胞內外泌體產生和分泌的機制[1]Fig.1 Mechanism of production and secretion of cell internal and external secretory bodies[1]Note:ESCRT.Endosomal sorting complex required for transport;SNAREs.Soluble NSF-attachment protein receptor.
近年來的研究表明,免疫細胞、腫瘤細胞、干細胞、上皮細胞等細胞均能夠產生外泌體。這些外泌體能對巨噬細胞、樹突狀細胞(Dendritic cell,DC)、T淋巴細胞、B淋巴細胞、自然殺傷細胞(Natural killer cell,NK cell)等免疫細胞進行調控。同時免疫細胞也能產生外泌體參與免疫應答(圖2),發揮抗原遞呈、免疫激活、細胞殺傷及調節代謝等眾多作用。
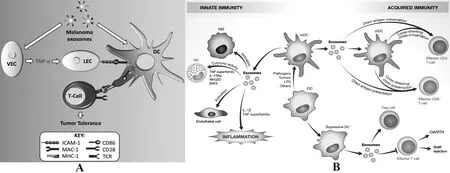
圖2 外泌體與DC的相互作用[2,3]Fig.2 Interaction between exosome and DC[2,3]Note:A.The stimulatory effect of tumor-derived exosomes to DC;B.The mechanism of DC regulating immune response through exosomes.
2 不同來源外泌體對免疫細胞的作用
目前的研究表明,來自腫瘤細胞、干細胞以及一些受到外源刺激的免疫細胞和組織細胞分泌的外泌體能夠靶向刺激免疫細胞, 對免疫系統的功能進行調控。同時,不同免疫細胞之間也會通過外泌體來協調免疫過程,這些外泌體對免疫細胞所引發的免疫調控機制也是免疫應答中的重要組成部分。
2.1外泌體影響巨噬細胞的極化 巨噬細胞作為非特異性免疫的重要組成部分,既是非特異性免疫的執行者,也是免疫信號的重要傳遞者。一般認為,在不同細胞因子的刺激下,組織內巨噬細胞可極化為M1和M2兩種亞群,分別起到介導炎癥反應和抑制炎癥的作用。隨著研究的深入,人們發現外泌體對巨噬細胞的功能調控主要通過控制其極化來實現。而let-7b/tLR 4通路可能是外泌體誘導巨噬細胞極化的重要方式[4]。體外實驗發現,來自抗原遞呈細胞的外泌體通過攜帶的脂多糖等抗原物質誘導極化[5];腫瘤細胞釋放的外泌體中則攜帶有血小板反應素1,能夠激活巨噬細胞并使其向M1型極化[6]。小鼠的背根神經節感覺神經元在外周軸突損傷后,會釋放miR-21-5p過表達的外泌體,來促進巨噬細胞的M1極化[7]。能夠促進巨噬細胞M2極化的外泌體也有許多。人臍帶間充質干細胞(Mesenchymal stem cells,MSC)能通過外泌體作用于體外培養的骨髓來源巨噬細胞,誘導其從M1型極化到M2型并下調炎癥因子的釋放,以利于脊髓損傷后的修復[8];受到脂多糖刺激的MSC分泌的外泌體對大鼠巨噬細胞的調節作用更為明顯,能夠促進大鼠的創傷愈合[4]。在小鼠體內腫瘤細胞的上皮間充質轉換過程中會釋放含有miR-21的外泌體,誘導巨噬細胞的M2極化[9]。此外腫瘤細胞也可以通過GP130/STAT3[10]、PTEN/PI3Kγ[11]等信號通路來促進極化。另外的一些研究表明,小鼠的脂肪干細胞可能通過miR-30D-5P在體內刺激巨噬細胞的M2極化作用[12]。在體外實驗中,來自氧化低密度脂蛋白誘導的內皮細胞中的外泌體可利用攜帶的轉移相關肺腺癌轉錄本1促進M2巨噬細胞極化,這可能與動脈粥樣硬化發生有關[13];暴露在酒精中的單核細胞釋放的外泌體利用攜帶的miR-27a與初始單核細胞溝通,誘導其極化為M2型巨噬細胞[14]。
這些研究表明,促進巨噬細胞向M1型極化的外泌體主要來源于抗原遞呈細胞、腫瘤細胞和神經元細胞等。而誘導巨噬細胞向M2方向極化的外泌體可來自MSC、腫瘤細胞、脂肪干細胞、上皮細胞和酒精處理過的單核細胞等。這些外泌體促進極化的作用與其攜帶的miRNA有著密切的關系。
2.2外泌體對T細胞分化與功能調控的影響 T細胞作為免疫系統的重要組成部分,一方面是免疫信息傳遞的重要環節,另一方面也能直接發揮免疫殺傷效應。T細胞包括眾多亞群,如細胞毒性T細胞(Cytotoxic T-lymphocyte,CTL)、抑制性T細胞(Suppressor T cell,Ts)、調節性T細胞(Regulatory T cell,Treg)和輔助性T細胞(Helper T cell,Th)等。這些亞群分化自T原細胞,其可在外泌體的刺激下分化成相應的功能性T細胞。
研究表明,小鼠體內MSC來源的外泌體能誘導T原細胞向Treg的分化,這一過程依賴抗原呈遞細胞的存在[15];DC來源的外泌體能夠刺激T原細胞PI3K/mTOR軸影響其分化,進而控制Treg和Th17細胞的平衡[16];受到致病菌刺激的上皮細胞也能夠通過外泌體促進Th17的增殖,進而促進結腸腫瘤生長[17]。從小鼠肥大細胞培養液中提取的外泌體可增強CD4+T細胞向Th2細胞分化[18]。在癌癥患者體內提取的腫瘤細胞中衍生的外泌體能夠體外調節T細胞產生細胞因子,并下調活化T細胞中CD3ζ與JAK3的表達,通過Fas/FasL(Fas配體)誘導活化的CD8+T細胞凋亡[19];也可通過影響Th1和Th17的分化與促進Treg產生來損傷T細胞功能[20]。
外泌體除參與T細胞分化外,也與T細胞功能調控有關。腫瘤細胞與DC來源的外泌體能夠介導抗原特異性T細胞活化和免疫活性改變[21]。被腫瘤細胞激活的DC產生的外泌體,能在小鼠體內以MHCⅠ依賴的方式促進CTL活化,使CD8+T細胞分化為CTL效應細胞,產生T細胞依賴性抗腫瘤免疫[22,23];卵清白蛋白(OVA)特異性T細胞分泌的外泌體能夠抑制小鼠體內OVA激活的樹突狀細胞,刺激CD8+CTL反應,進而抑制抗腫瘤免疫[24]。在一些體外實驗中發現,腫瘤細胞來源外泌體還會導致Treg強烈和持續地生成肌苷[25],并通過調控TGF-β和IL-10表達,來提高Treg的增殖能力,從而增加其對細胞凋亡的抗性[26,27]。Treg能將miRNA轉移到Th1中,抑制Th1增殖和細胞因子分泌[28]。此外,腫瘤來源外泌體還能夠改變CD4+或CD8+T細胞的炎性因子表達[20]。
從這些研究中我們可以看出,T細胞的分化可受到來自MSC、DC、腫瘤細胞、肥大細胞、上皮細胞等多種來源外泌體的影響。同時,來自腫瘤細胞、DC、Treg和被激活的T細胞能夠調控受體T細胞的狀態,包括調節細胞活性、CTL反應及改變細胞因子表達等。
2.3外泌體對B細胞狀態與功能的影響 作為機體免疫中重要的效應細胞,B細胞的細胞膜表面帶有多種類型的受體,在體液免疫過程中,B細胞能夠分化為漿細胞,并分泌特異性抗體。有研究顯示,外泌體能通過淋巴管迅速地從周邊傳輸到淋巴結,而淋巴結中的B細胞則是外泌體的主要攝取者之一[29]。
在體外共培養條件下,MSC細胞衍生的外泌體能夠抑制B細胞的炎癥作用,并影響B細胞的分化[30];骨髓基質細胞來源外泌體也可抑制慢性淋巴細胞白血病B細胞的自發凋亡,并增強其耐藥性和遷移能力[31];而DC來源的外泌體也能夠通過補體激活和抗原穿梭的方式參與介導T細胞和B細胞的激活,誘導抗腫瘤免疫[32]。腫瘤細胞來源的外泌體能夠通過脂質筏/膽固醇內吞途徑來刺激人B細胞[33],并增強B細胞的增殖與向漿細胞的分化[34]。
而當B細胞受到病毒的侵染時,外泌體還會作為載體,調控B細胞的狀態并參與物質運輸。如EB病毒感染的B細胞能夠被人口腔上皮細胞分泌的富含miR-200家族成員的外泌體觸發激活,并促進B細胞與口腔上皮細胞間的病毒交換[35]。
通過這些研究可知,來自MSC、DC、腫瘤細胞、上皮細胞等細胞的外泌體能夠改變B細胞的分化、激活、增殖和凋亡,并影響其耐藥性和遷移能力。
2.4外泌體對樹突狀細胞調控和表達的影響 樹突狀細胞(DC)分為髓樣DC和淋巴樣DC,是一種功能強大的抗原遞呈細胞,與腫瘤的產生和發展有著緊密聯系。在這一過程中,外泌體的作用也不容忽視。
小鼠體內的腫瘤細胞能夠分泌攜帶腫瘤抗原、LAMP1、OVA和pMHCⅠ等分子的外泌體,并轉移到DC中,引發DC的抗腫瘤免疫[22,36]。而DC的抗腫瘤免疫過程也是通過外泌體實現的。胰腺癌細胞株的體外實驗發現腫瘤細胞外泌體能夠將miRNA轉移到DC中,改變DC的miRNA表達并抑制調節因子X相關蛋白(rFXAP)產生,這可能是導致MHCⅡ分子抑制和免疫耐受的原因[37];同來源的外泌體也能下調DC中的TLR4和下游細胞因子[38]。
此外,DC也可以接受來自B細胞的外泌體并調控T細胞的分化;T細胞能夠利用攜帶含有基因組或線粒體DNA的外泌體,通過cGAS/STING胞質DNA傳感通路與IRF3依賴性干擾素調節基因的表達,誘導DC的抗病毒反應[39]。
以上研究表明,DC能夠被來自腫瘤細胞和其他免疫細胞的外泌體調控。這些外泌體能通過傳遞抗原、miRNA等多種物質,影響DC的增殖、活化以及細胞因子和miRNA的表達。
2.5外泌體對NK細胞腫瘤殺傷作用的影響 自然殺傷細胞(Natural killer cell,NK)是先天免疫細胞,其在機體防御的第一線尤其是腫瘤殺傷過程中起關鍵作用。而外泌體可顯著影響NK細胞殺傷作用。
眾多體外實驗表明,腫瘤細胞來源外泌體可通過攜帶的IL-15、IL-18等細胞因子以及4-1BBL蛋白類似物,促進NK細胞的活化與增殖,增強其敏感性與抗腫瘤效力[40,41];也可攜帶NF-κB-α激活激酶相關蛋白1(NAP-1),通過增加NK細胞中干擾素調節因子3(IRF-3)的表達及其磷酸化,來提升Ⅰ型干擾素基因和趨化因子配體基因的表達,促進NK細胞的功能[42]。而這些外泌體攜帶的TGFβ1,能夠下調NK細胞NKG2D受體表達,誘導NK細胞中Smad磷酸化[19];或利用miR210和miR-23α等miRNA,直接影響NK細胞中CD107a和IFNg的表達,降低其細胞毒性與抗腫瘤活性[43];腫瘤來源外泌體還能抑制NK細胞活化受體表達,來降低NK細胞的腫瘤殺傷作用[40]。DC則通過分泌表達白細胞介素-15Ra和NKG2D配體的外泌體,直接參與IL-15Ra與NKG2D依賴的NK細胞增殖和活化,促進分泌IFNc[44]。
由此可見,NK細胞能夠受到腫瘤細胞和DC來源的外泌體調控。其中腫瘤細胞來源的外泌體能起到雙向調控的作用。一方面,這些外泌體能夠激活NK細胞,促進其增殖,并提高對腫瘤細胞的殺傷;另一方面,也可通過抑制細胞毒性而逃避免疫監視。而DC來源的外泌體主要對NK細胞起到激活作用,并促進NK細胞的增殖。
3 免疫細胞來源外泌體對其他細胞的功能
外泌體除了能調控免疫細胞功能外,還是免疫細胞功能調控的執行者。免疫細胞來源的外泌體能夠調控包括免疫細胞、上皮細胞、腫瘤細胞等多種受體細胞并改變微環境。進而對整個免疫應答造成影響。
3.1巨噬細胞來源外泌體在機體免疫應答及調控過程中的作用 作為非特異性免疫的重要效應細胞和抗原呈遞細胞,巨噬細胞也可利用外泌體釋放信號。這些信號隨著其所在微環境和受到刺激的不同而改變,當受到結核分枝桿菌、沙門氏菌、弓形蟲、艾滋病毒等病原體的感染和一些外來抗原的刺激時,巨噬細胞釋放的外泌體能攜帶病原體相關的分子模式受體(PAMPs)[45],同時會導致外泌體中miRNA的種類與含量發生變化[46]。
一些體外實驗表明,從鼠傷寒沙門氏菌感染的巨噬細胞中分泌的促炎外泌體能觸發幼稚巨噬細胞和DC Toll樣受體4依賴性的TNF-α釋放,以及其他多種細胞因子的分泌,這可能與外泌體中包含的LPS有關[47];而LPS對巨噬細胞的激活也能通過影響其釋放的外泌體中miRNA與蛋白表達,引起脂肪細胞基因表達的改變。這些變化與補體活化、活性氧種類調節、白細胞遷移與活化、單核細胞趨化等炎癥反應有關,也對碳水化合物的分解代謝和細胞活化產生影響[48]。同時在利用病原菌感染小鼠的研究中發現,有的病原菌如結核分枝桿菌,其感染的巨噬細胞能夠通過外泌體的途徑呈遞抗原,并能協同ATP促進MHC-Ⅱ的釋放,通過外泌體向T細胞表達抗原肽,這在促進T細胞免疫的過程中起重要作用[49,50];而在病原體的刺激下,巨噬細胞產生的外泌體能夠促進TNF-α和IL-12的產生以及中性粒細胞和巨噬細胞的募集,這可能是免疫激活的一部分[45]。
關于巨噬細胞來源外泌體對上皮細胞的影響,體外實驗結果表明,巨噬細胞釋放的外泌體能夠刺激內皮細胞中的整合素向溶酶體轉運,誘導整合素蛋白降解,進而對內皮細胞的遷移起到負調節作用[51];在LPS誘導的膿毒性肺損傷小鼠模型中,被LPS激活的巨噬細胞釋放的外泌體中,促炎性miRNA(miRNA-155、miRNA-146a)表達上調,使上皮細胞中的促炎介質TNF-α與IL-6表達增加,進而破壞支氣管上皮細胞緊密連接蛋白的表達,對肺部的結構屏障造成破壞[52]。有研究者嘗試在體外將巨噬細胞來源的外泌體作為載體承載miRNA抑制劑,發現其能夠調控腫瘤細胞中的miRNA表達水平[53]。這可能會成為腫瘤治療的一種新方式。
以上結果表明,巨噬細胞來源的外泌體能作用于其他免疫細胞、脂肪細胞、上皮細胞與腫瘤細胞,發揮著抗原呈遞、免疫調節及代謝調控等作用。
3.2T細胞來源外泌體的腫瘤殺傷及免疫激活作用 在T細胞中,外泌體的釋放受到二酰基甘油和二酰基甘油激酶(Diacylglycerol kinase α,DGKα)的調節,DGKα能夠調控PKD1/2的亞細胞定位與激活,PKD1/2是外泌體分泌的關鍵調節分子,介導DGKα對外泌體分泌通道的影響[39]。而這些外泌體在體外能夠發揮多種作用。
大鼠體內OVA抗原特異性的T細胞外泌體能夠抑制DC對CD8+T細胞的刺激,進而抑制DC介導的大鼠CD8+CTL糖尿病和抗腫瘤免疫[24]。而在小鼠體內,Treg來源的外泌體還能將miRNA轉移到Th1細胞中,抑制Th1細胞增殖和細胞因子分泌[28]。體外實驗表明,CTL能通過外泌體來釋放TNF超家族,誘導腫瘤細胞死亡[54]。在脾和淋巴結部位衍生的Ts則能分泌抑制性外泌體,調節免疫炎癥反應外周組織部位抗原遞呈細胞和效應T細胞功能[55]。受到LPS刺激的Treg衍生的外泌體能攜帶miR1505p和miR-142-3p調控DC,使IL-10過表達并抑制IL-6表達來抑制免疫反應,這可能有助于治療自身免疫性疾病[56]。而在受到腺病毒基因刺激時,Treg會分泌含有獨特的miRNAs和iNOS分子的外泌體。其能誘導細胞凋亡,并促進T細胞向Treg的轉化[57]。
通過這些研究可知,T細胞來源外泌體能夠作用于DC、腫瘤細胞、抗原遞呈細胞和其他T細胞,其功能包括激活T淋巴細胞介導的細胞毒性和殺傷作用以及多種免疫調節過程。
3.3B細胞來源外泌體對免疫細胞的作用 與T細胞類似,在B細胞內MVB成熟和外泌體分泌的過程中,PKD1/2蛋白扮演著關鍵調節因子的角色[58],能夠影響細胞功能并造成微環境的改變。
體外研究表明,這些外泌體能夠傳遞miRNA-155來抑制肝細胞中丙肝病毒的活性[59],還能攜帶過敏原衍生肽以誘導T細胞增殖和Th2樣細胞因子的產生[60]。而一些病毒對B細胞的感染也會改變其外泌體成分與功能,使之對細胞代謝產生影響,進而改變腫瘤微環境[61]。同時,B細胞外泌體能在小鼠體內激活特異性T淋巴細胞、誘導體內的細胞毒性反應[62]。
B細胞淋巴瘤是一組淋巴組織的惡性腫瘤,其特征為B淋巴細胞的惡性轉化。B細胞淋巴瘤細胞來源的外泌體在腫瘤發生過程中起雙重作用。一方面,來自腫瘤細胞的外泌體能將腫瘤抗原和MHCⅠ類分子傳遞給樹突狀細胞,然后通過誘導CD8+T細胞依賴性的抗腫瘤免疫應答,直接抑制惡性腫瘤的發生[63];而另一方面也能誘導免疫細胞的凋亡,并通過抑制NKG2D受體介導的細胞毒性,從而損害NK細胞功能[64]。對外泌體在B細胞淋巴瘤中的作用研究,為淋巴瘤的抑制和治療提供了新思路。
通過對這些研究的總結可知,來自B細胞的外泌體可以作用于肝細胞、T細胞、DC及NK細胞,影響細胞的激活、代謝與凋亡。
3.4樹突狀細胞來源的外泌體及其功能 DC來源的外泌體主要通過激活其他免疫細胞來發揮功能,作為免疫調節的一種重要途徑,能夠在細菌、寄生蟲、病毒的感染和抗腫瘤過程中發揮作用。
通過對小鼠的研究發現,被腫瘤激活的DC釋放的外泌體,在體外和體內均能刺激CD8 T細胞增殖,并傳遞抗原物質與功能性MHC-肽復合物,有效地誘導特異性CTL反應、抗腫瘤免疫和CD8 T細胞記憶[36,65];而在寄生蟲感染過程中,受到抗原刺激的DC能夠通過外泌體的方式抵抗炎癥反應[66]。同時,DC來源的外泌體能利用功能性IL-15與NKG2D配體直接介導NK細胞的活化[45],或通過攜帶TNF超家族配體激活NK細胞TNF-γ的分泌,并能在腫瘤細胞表面表達TNF、FasL和TRAIL,從而觸發腫瘤細胞Caspase的活化和凋亡[67]。體外研究則表明,DC來源的外泌體也能夠促使NK細胞的增殖與分泌IFNc,抑制腫瘤的轉移[68];還能與細菌Toll樣受體配體結合,對未活化的DC有著強烈的激活作用[69]。此外,DC來源的外泌體也會幫助病原體入侵機體。HIV-1會通過刺激樹突狀細胞免疫受體,來抑制外泌體從DC中釋放,并改變外泌體內容物的成分,使之富含促凋亡分子DAP-3,以增加CD4+T淋巴細胞的凋亡[70]。此外,DC來源的外泌體也可能與朊病毒的體內傳播有關[71]。
DC來源的外泌體能對T細胞、B細胞、NK細胞以及未激活的DC起作用,其作用包括參與免疫激活、免疫調控與抗腫瘤作用。DC來源外泌體功能的多樣性也為研究者們提供了靶向給藥的新思路。
3.5NK細胞來源外泌體的抗腫瘤作用 NK細胞的殺傷作用主要有兩種機制:①釋放穿孔素和顆粒酶,通過增加凋亡小體的形成和caspase-3激活而誘導內在凋亡途徑;②涉及caspase-8a的外源性凋亡途徑。NK細胞主要通過表達FasL和穿孔素等殺傷蛋白發揮細胞毒性作用,參與抗腫瘤和免疫穩態的調節。而NK細胞在靜息和激活條件下都會釋放出外泌體,能夠作為FasL、顆粒酶B、穿孔素與TNF-α的載體,將其分泌到細胞外[72,73]。同時體外實驗表明,這些外泌體能攜帶NK細胞的標記蛋白CD56,可能參與促進外泌體與靶細胞的黏附,對不同的腫瘤靶細胞和活化的免疫細胞發揮細胞毒活性[74]。NK細胞分泌的攜帶TNF-α的外泌體還能夠在腫瘤處特異性聚集,在體外和體內均對膠質母細胞瘤細胞有靶向和抗腫瘤作用[72]。并且NK細胞也會利用外泌體來激活其他NK細胞的活性,并傳遞殺傷物質。
根據目前的報道,NK細胞作為殺傷腫瘤的重要細胞,其分泌的外泌體主要作用包括向腫瘤細胞靶向傳遞殺傷物質以及免疫激活,發揮抗腫瘤功能。
4 展望
外泌體作為一種廣泛存在于細胞間的信息傳遞方式,由于其迅速和直接跨膜的特性而受到廣泛關注。隨著研究的深入,外泌體在免疫細胞應答過程中的作用逐漸被揭示。外泌體在免疫細胞與腫瘤細胞、干細胞、其他免疫細胞及一些組織細胞間的信息傳遞中發揮作用,調控機體免疫和腫瘤抵抗過程,其中也不乏雙向調控的情況出現。此外,這些機制也與細胞代謝和干細胞分化有關。深入探討外泌體的免疫調節方式及相關機制,對進一步完善對機體免疫過程的認識具有重要意義,也能夠為相關研究提供參考。