高低應答CO2水稻品種苗期根系對高碳環境的響應①
2020-01-06 07:51:10吳晶晶施衛明
土壤 2019年6期
關鍵詞:水稻
吳晶晶,施衛明
高低應答CO2水稻品種苗期根系對高碳環境的響應①
吳晶晶1,2,施衛明1*
(1 土壤與農業可持續發展國家重點實驗室(中國科學院南京土壤研究所),南京 210008;2 中國科學院大學,北京 100049)
本文利用水培試驗和瓊脂板培養試驗研究了高CO2條件下產量響應存在顯著差異的兩個水稻品種:II優084(高響應)和武運粳23(低響應),在幼苗期根系形態對高C的響應差異。水培試驗結果表明,在幼苗時期,高應答品種II優084在低氮條件下地上部生物量在高CO2下增加28.5%,根系干物質量對高CO2響應顯著,增幅為28.5%,而其不定根數目沒有顯著增加,對干物質量響應貢獻較大的為總根長。II優084的總根長在高CO2下增幅為26.3%,不同根粗的根長均有高響應。低應答品種武運粳23低氮下地上部和根系響應不顯著,而在正常氮和高氮下則不同。正常氮條件下,地上部對高CO2響應不顯著,而根系生物量在高CO2下顯著增加76.0%,不定根數目增加25.8%,同時總根長增加45.0%,不同根粗的根長均有高響應,II優084則沒有顯著響應。在高氮條件下,武運粳23地上部生物量在高CO2下增加35.5%,根系生物量增加80.3%,不定根數目增加38.5%,根系平均直徑增加16.7%,總根長無響應,而II優084生物量在高氮下無顯著差異。同時,武運粳23在正常氮和高氮下的根系表面積和體積對高CO2響應也較II優084顯著。瓊脂板培養試驗的結果與水培結果一致,武運粳23根系形態對高濃度蔗糖的響應普遍高于II優084。試驗結果說明品種對高C環境的響應特征不隨培養條件的變化而變化。與植株生長后期不同,在幼苗期正常氮條件下低應答品種武運粳23的根系生物量和各形態指標對高C的響應明顯高于II優084,說明水稻苗期生長響應參數與后期產量響應參數不一定一致,可能是由于苗期生長高響應的品種在營養生長期旺長,反而不利于后期生殖生長,從而導致后期產量的低響應。
高C環境;培養條件;品種差異;根系形態
根據NOAA (National Oceanic and Atmospheric Administration)的最新數據,2017年全球大氣CO2平均濃度已達到0.793 mg/L,且仍然處于上升的趨勢[1]。除溫室效應導致的全球氣候變化對植物的間接影響外,大氣CO2濃度升高還直接影響植物的生長發育[2]。水稻是重要的糧食作物,其在CO2濃度升高條件下的生長發育和產量的響應已被國內外學者廣泛關注和報道[2-7]。CO2濃度升高,水稻光合速率短期內顯著增加[8-9],氣孔導度變小[10],分蘗和有效穗數明顯增多,生物量和產量也相應增加[11-15]。
根系是植物吸收水和營養元素的重要通道。植物對其所處環境的適應性,在很大程度上取決于植物根系的形態、結構特性和生理特點。許多研究表明,高濃度CO2條件下,植物除地上部的生長受到影響外,根系的長度、密度、粗細、生物量、根冠比、生理特性以及根系分泌物等也會發生明顯變化[16-21],從而有助于植物從土壤中獲取更多的水分和養分,更好地適應CO2濃度升高的環境。Kim等[4]報道在FACE條件下水稻品種Akitakomachi的根系較對照有明顯增加,且根系生物量與氮素吸收呈正相關關系。陳改蘋等[18-19]報道指出,在水稻各生育期,CO2濃度升高都極顯著增加了干物質量,且主要增加根粗為2.0 ~ 2.5 mm的部位,根冠比也顯著增加。Yang等[22]報道武香粳14的高CO2條件下不定根數和根長分別比對照條件下增加25% ~ 31% 和25% ~ 37%。我們前期的研究也表明FACE條件下產量高響應的品種II優084的根系干物質量在分蘗期后各生育期增幅均大于30%,而產量響應不明顯的品種武運粳23的根系生物量增幅較小或者不顯著[23]。
由于田間試驗是在水稻苗播種約1個月移栽后才開始進行高CO2處理,因此對兩水稻品種幼苗期在高C環境的響應是否與中后期的表現相一致,我們并不清楚。水稻品種II優084在高CO2下穗重產量和生物量的響應明顯高于品種武運粳23[23],結合前人篩選的其他水稻品種的產量響應數據[24],在本文中將II優084歸類為高應答水稻品種,將武運粳23歸類為低應答水稻品種。為了探究高、低應答兩個水稻品種的根系在幼苗期對高C的響應,本試驗分別用水培和瓊脂板兩種方法為幼苗地上部提供高C環境,比較兩個品種苗期根系形態的應答差異。同時比較它們在幼苗期與生長中后期響應的特點。
1 材料與方法
1.1 田間試驗
水稻FACE(Free-air CO2enrichment)基地位于揚州市江都區小紀鎮良種場(119°42'0"E,32°35'5"N)。平臺共有3個FACE試驗圈和3個對照圈,FACE圈與圈以及FACE圈與對照圈之間間隔大于90 m,以減少CO2釋放對其他圈的影響。通過FACE圈周圍管道向圈內噴射純CO2氣體。通過計算機網絡對圈內CO2濃度進行監測和控制,使水稻生長時期內圈內CO2保持比大氣背景CO2濃度高200 μmol/mol。對照沒有安裝管道,其他條件與FACE圈保持一致。同時每個圈內設有不施氮肥和正常施氮肥兩個區域。本試驗供試水稻品種為II優084(IIY)和武運粳23(WYJ)。于2014年5月21日播種于大氣CO2條件下,6月20日移栽于FACE和對照圈內的兩個氮肥處理區域。施肥水平與當地常規水平一致,分別是總氮 22.5 g/m2,總磷 9 g/m2,總鉀 9 g/m2。磷鉀肥作為基肥一次性施加;氮肥分3次施加:基肥40%、分蘗肥30%、穗肥30%,分別于6月20日、6月28日、8月1日施加。田間管理和病蟲害防治等為大田常規管理。
1.2 試驗設計
室內水培試驗在可控CO2濃度的兩臺植物光照培養箱中進行。箱內配有進口CO2紅外傳感器,將信號傳入控制器以控制箱內CO2濃度。培養箱中CO2濃度調節通過外源接鋼瓶通入CO2和每天更換CO2吸附劑(氫氧化鈣)進行。當箱內CO2濃度低于設定值時,控制器會自動打開通氣閥通入CO2直到箱內達到設定值;當箱內CO2濃度高于設定值時通氣閥關閉,箱內多余的CO2會被吸附劑緩慢吸附。本試驗設置兩培養箱內CO2濃度分別為0.746 mg/L(對照)和1.492 mg/L(處理)。光周期設置為12 h白天/12 h夜晚,相對濕度設置為80%,溫度為30 ℃白天/25 ℃夜晚。
水培營養液選用國際水稻研究所(IRRI)常規營養液配方加以微調,添加 Na2SiO3以保持營養液中的 SiO2為 0.1 mmol/L。常規營養液配方為:1.25 mmol/L NH4NO3,0.3 mmol/L KH2PO4,0.35 mmol/L K2SO4,1 mmol/L CaCl2·2H2O,1 mmol/L MgSO4·7H2O,0.5 mmol/L Na2SiO3·9H2O,9 μmol/L MnCl2·4H2O,0.39 μmol/L Na2MoO4·2H2O,20 μmol/L H3BO3,0.77 μmol/L ZnSO4·7H2O,0.32 μmol/L CuSO4·5H2O,20 μmol/L EDTA-Fe。正常N 處理(MN)采用1.25 mmol/L NH4NO3,低N處理(LN)采用0.25 mmol/L NH4NO3,高N處理(HN)采用3.75 mmol/L NH4NO3。
供試水稻同樣用II優084和武運粳23,挑選大小均勻、顆粒飽滿的種子用10% 的H2O2消毒30 min,用去離子水沖洗5次后30 ℃下在 0.5 mmol/L CaSO4中浸種1 d,后黑暗催芽48 h。然后挑選萌發的種子播于網篩上,1/4全營養液培養3 d后,1/2全營養液培養至10 d后挑選長勢一致的苗進行處理,期間2 d換一次全營養液,處理3周后將植株分為地上部和根系,地上部樣品直接105 ℃殺青30 min,80 ℃烘干后測干物質量,根系樣品對其進行根系形態統計后殺青、烘干測其干物質量。
瓊脂板培養,先將營養液中加入瓊脂粉制普通板和上下分離板留存,其中上板中加入不同含量蔗糖(0、10、50、100、200 g/kg)處理。在超凈工作臺上將挑選的已去殼的種子先用無水乙醇浸泡1 min,無菌水清洗3 ~ 5次后加入20% 次氯酸鈉和1滴吐溫80輕搖、浸泡20 min后用無菌水清洗5 ~ 10次。將消過毒的種子放在滅菌的密封容器中,加入適量無菌水,30 ℃黑暗浸種、催芽48 h后播于普通瓊脂板上置于光照培養箱中培養2 ~ 3 d后,將幼苗去胚乳移至上下分離板處理1周后取樣監測兩品種根系的響應。
根系形態分析采用根系掃描儀掃描,根系分析軟件WinRHIZO2012分析獲得數據。
1.3 數據處理
數據均采用SPSS分析軟件進行方差分析和多重比較。
2 結果
2.1 水培條件下,兩水稻品種苗期地上部對高CO2的響應差異
幼苗時期,II優084的地上部生物量在不同N水平下均高于武運粳23,而兩品種地上部生物量對高CO2的響應卻因N水平不同而不同。如圖1A所示,在LN處理下,II優084的地上部生物量在高CO2下增加了28.5%,武運粳23則無顯著變化;在MN條件下,高CO2對兩品種地上部生物量的增加效應均不顯著;而在HN處理下,高CO2對低應答品種武運粳23的增幅高達35.5%,對高應答品種II優084的增幅卻不明顯。
高CO2對早期分蘗影響情況統計顯示,武運粳23在LN時分蘗略有增加,但是增加不顯著;MN時高 CO2未增加武運粳23分蘗;HN時,分蘗在高CO2處理下增加了17.7%。而對II優084來說,LN下分蘗增加不顯著,MN及HN下,CO2增加分蘗達28.6%和25.8%,均高于同條件下的武運粳23(圖1B)。
2.2 水培條件下,兩水稻品種根系生物量對高CO2的響應差異
如圖2所示,與地上部相似,II優084的根系生物量也普遍較武運粳23大。LN下,高CO2顯著增加II優084根系,增幅達31.8%,武運粳23增加不顯著(圖2)。在MN和HN條件下,高CO2對武運粳23根系生物量的增幅為別為76.0% 和80.3%,而對II優084的根系生物量增幅不顯著(圖2)。
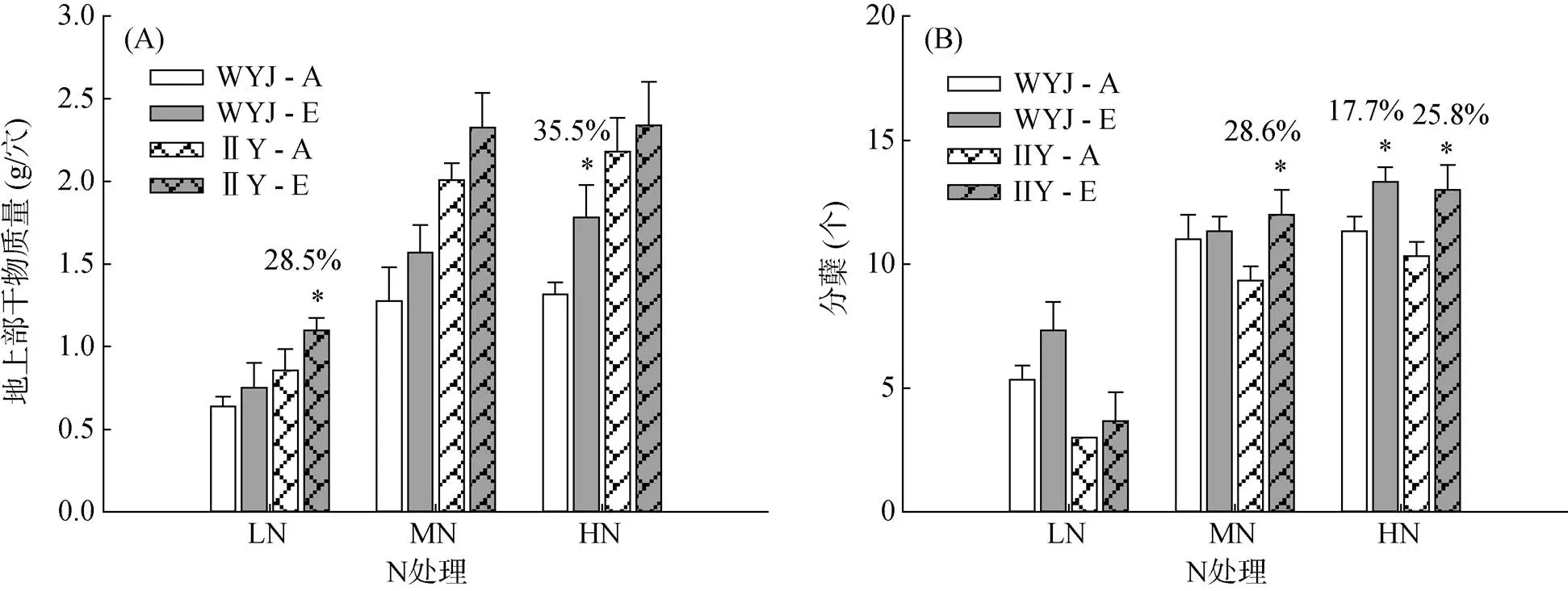
(WYJ:武運粳23;IIY:II優084;CO2兩個濃度水平:0.746 mg/L (A)、1.492 mg/L (E);*代表同一水稻品種同一N處理下不同CO2處理間差異顯著(P < 0.05);百分數值來源于(E–A)/A;下圖同)

圖2 兩水稻品種苗期在CO2和N的6種處理組合下的根系干物質量
2.3 水培條件下,兩水稻品種根系形態對高CO2的響應差異
我們從不定根數目、總根長、直徑小于0.5 mm的根長、直徑大于0.5 mm的根長、根平均直徑、根表面積和根體積等方面考察兩水稻品種的根系形態對高CO2的響應。由圖3A可知,LN條件下,兩品種不定根數目對高CO2無明顯響應;在MN和HN下,武運粳23的根系不定根數目在高CO2下分別增加了25.8% 和38.5%,而II優084無明顯響應。由圖3B可知,II優084的總根長僅在LN條件下對高CO2有明顯響應,增幅為26.3%;而武運粳23的總根長僅在MN下對高CO2有顯著響應,增幅達45.0%。進一步對根系生長進行分析,由圖3C可知,在LN條件下,II優084直徑小于0.5 mm的根長增幅為25.9%;MN條件下,武運粳23直徑小于0.5 mm的根長增幅為43.5%。圖3D為直徑大于0.5 mm的根長,武運粳23在3種N水平下響應均顯著,增幅分別為49.5%、63.5% 和50.8%,而II優084僅在LN下表現出顯著增加,增幅為30.2%。
兩品種根系平均直徑對高CO2響應比較如圖3E所示,LN條件下,兩品種間及CO2處理間均無明顯響應;MN和HN下,II優084根系直徑均大于武運粳23。CO2響應只在HN下的武運粳23中表現,增幅為16.7%。根系表面積對高CO2響應比較如圖3F所示,LN條件下,兩品種CO2處理間均無明顯響應;MN和HN下,II優084無響應,而武運粳23對高CO2響應顯著,分別增加53.1% 和31.0%。根體積與表面積響應相似,II優084的根體積在3個N水平下對高CO2均無明顯響應;武運粳23在LN下無響應,在MN和HN條件下明顯增加,增幅分別為65.8% 和55.4% (圖3G)。

圖3 兩水稻品種苗期在CO2和N的6種處理組合下的根系形態
2.4 瓊脂培養條件下,兩水稻品種根系形態對高C的響應差異
瓊脂板培養條件下,用蔗糖模擬高C處理水稻苗期地上部,觀察根系表型。總根長統計顯示(圖4A),兩品種根長在100 g/kg蔗糖處理時響應最顯著,相對不添加蔗糖對照來說,武運粳23在10、50、100 g/kg 蔗糖處理下的響應都明顯高于II優084,增幅明顯。此外,直徑小于0.2 mm的根長和直徑大于0.2 mm的根長在不同濃度蔗糖下的響應趨勢一致(圖4B、4C)。
根系平均直徑統計顯示(圖4D),10、50、100 g/kg 蔗糖處理時II優084的根系平均直徑明顯大于武運粳23。但是兩品種根系表面積(圖4E)隨蔗糖濃度的變化趨勢與總根長趨勢一致(圖4A);各蔗糖處理下兩水稻品種間根體積差異不大(圖4F)。
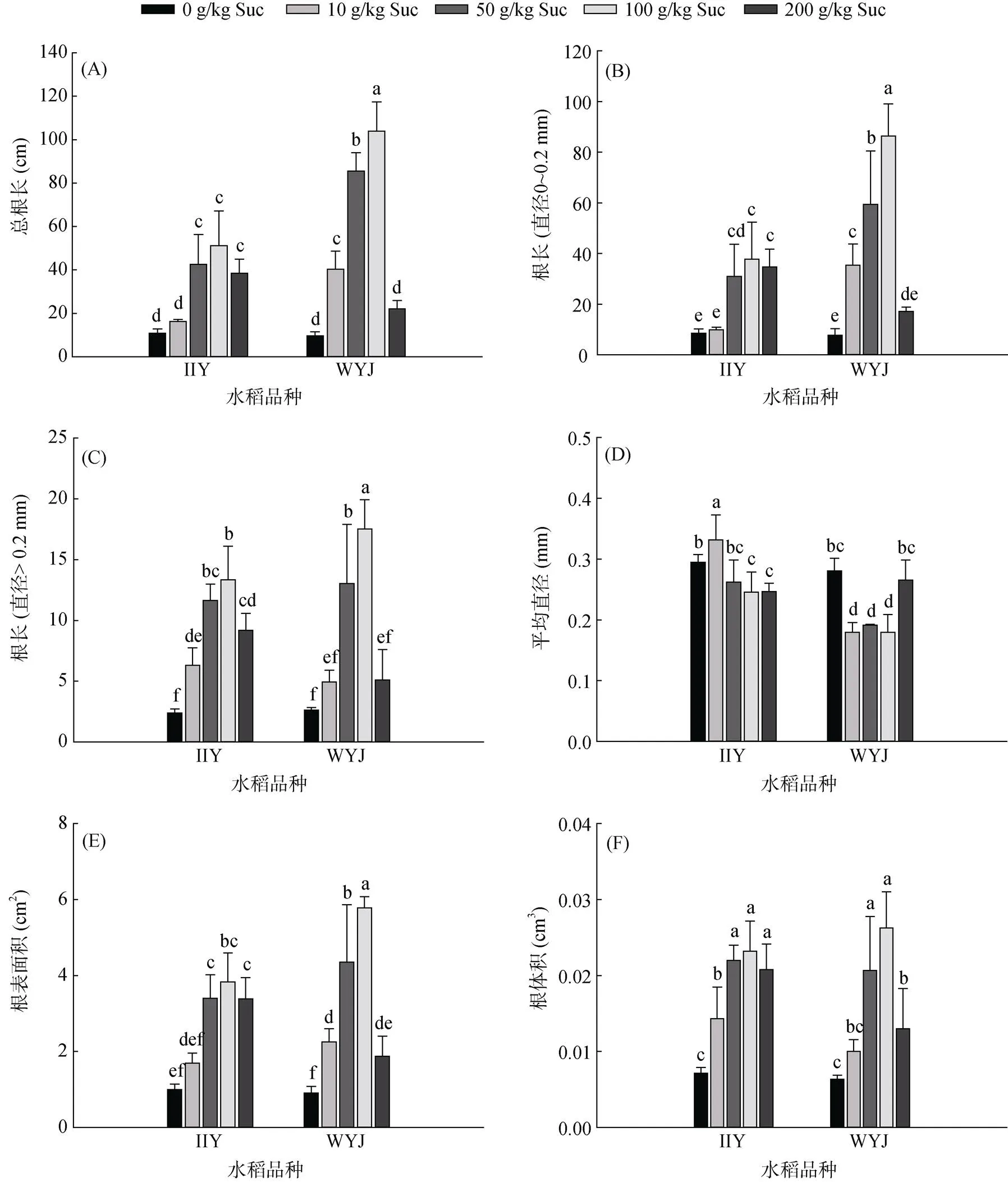
(圖中小寫字母不同表示不同水稻品種不同蔗糖處理間差異顯著(P < 0.05))
3 討論
目前關于高濃度CO2條件下水稻響應的研究大多集中于單一品種的生長響應和產量方面[4-5],在高CO2條件下,光合作用增強致碳水化合物增多,從而促進生物量和產量的提高。同時高CO2也影響水稻根系的生長和營養元素的吸收[21-22, 25-26]。CO2濃度升高對水稻生物量的影響,因供試品種、光照、溫濕度、地理位置等因素的不同而有不同。不同品種間響應差異的報道相對較少,尤其是根系響應的比較。
Zhu等[6]報道了高、低應答水稻品種的光合參數和CO2固定的相關基因的表達,發現高應答品種比低應答品種擁有更大的庫源比,使其在生育期維持更高的光合能力和產量。武慧斌等[27]利用Minirhizon-trons微根管技術監測高CO2條件下4個水稻品種根系的生長發育,發現根長密度、根表面積、根體積和根數都有不同程度的增加。本試驗運用了室內水培和瓊脂板培養兩種方法,監測田間FACE下產量響應差異明顯的兩品種在苗期對高C環境的響應,以更全面分析高、低應答水稻品種響應差異的表現和原因所在,為水稻適應高CO2環境的深入研究做鋪墊。
從生長情況和對高C環境的響應來看,在植物光照培養箱中提供高濃度CO2和在分離的瓊脂培養板上對地上部供應蔗糖兩種方法均能夠促進水稻幼苗根系的生長和根系形態的相應變化,說明兩種方法研究地上部供應高C對水稻幼苗影響的可行性。且相對來說,后者在設備要求和培養時間上更靈活,為以后高C環境的研究提供了思路。
我們的研究結果顯示,高C環境確實能夠促進水稻生長發育和根系形態的高響應,這與前人的結果相一致。同時我們更注重品種間的差異比較。低應答品種武運粳23苗期地上部生物量在高N(HN)下響應顯著,而高應答品種II優084地上部僅在低N(LN)下響應顯著(圖1A),說明水稻苗期地上部兩品種對高CO2的響應與N供應密切相關。分蘗情況顯示II優084對高CO2的響應更明顯(圖1B),分蘗響應與地上部生物量響應不一致,說明地上部生物量響應不依賴于分蘗的響應。相較于地上部生物量,根系生物量響應更顯著,且在正常N(MN)下武運粳23有明顯響應,而II優084無響應,說明在水稻苗期MN下武運粳23的根系較地上部響應早,推測高CO2下葉片產生的額外的光合產物首先用于根系的生長(圖2)。
由圖3可知,LN條件下II優084根系生物量的高響應(圖2)主要源自于總根長而非根系的不定根數目,且總根長在直徑0 ~ 0.5 mm和大于0.5 mm兩個水平均有明顯響應(圖3C、D);MN下武運粳23根系生物量的高響應表現在不定根數目(圖3A)、總根長(圖3B),且總根長在直徑0 ~ 0.5 mm和大于0.5 mm兩個水平均有明顯響應(圖3C、D);而HN條件下,武運粳23根系生物量的高響應則主要表現在不定根數目(圖3A)和根系平均直徑(圖3E)。說明高、低應答品種的根系響應特征隨施N水平而變化:在LN水平下,II優084根系優先增加根系長度,以增強根系對營養物質的吸收和增加生物量,不定根數目無明顯響應,武運粳23的不定根數目和總根長的響應均不顯著;在HN水平下,武運粳23則優先增加不定根數目,而總根長無明顯響應,II優084的不定根數目和總根長的響應均不顯著;而在MN水平下,武運粳23同時增加不定根數目和總根長以響應高CO2環境,II優084則兩者均無顯著響應。
不同N水平下,水稻的高CO2響應會表現出不同[5-12]。Kim等[12]的田間FACE試驗表明,LN下水稻的高CO2響應會受限制,而HN下的響應并沒有預想中的高。本試驗中,兩品種苗期在不同的供N環境下的響應差異也不同。就地上部生物量而言,武運粳23在LN和MN下無響應,在HN下有顯著響應,II優084則不同,在LN下高響應,在MN和HN下無顯著響應。就根系生物量而言,武運粳23在LN下無響應,在MN和HN下高響應,而II優084同樣是在LN下高響應,MN和HN下無顯著響應。總體來說,武運粳23更傾向于在MN和HN下對高CO2有響應,而II優084則更傾向于在LN下有顯著響應。相似的結果也表現在根系的不定根數目、總根長、根表面積和根體積等方面。對武運粳23而言,低N條件可能限制了C的同化利用,因此各生長指標無顯著響應,這與前人的FACE試驗結果相一致[5-12]。對II優084而言,N充足時無明顯高CO2響應,可能與其苗期生長速率較快有關,機體能夠有效利用有限的C資源用于自身生長,C源可能不是其生長的主要限制因子,而在LN下,機體能夠高效利用有限的N,此時C源成為主要限制因子,因此II優084能夠高響應。推測II優084有更強的C、N協同能力。
瓊脂板培養數據顯示,除200 g/kg蔗糖處理外,其余處理下武運粳23根系對高蔗糖響應均大于II優084,且100 g/kg蔗糖處理時最顯著。同時,總根長的響應差異主要表現在根系直徑位于0 ~ 0.2 mm之間的根系長度。在10、50、100 g/kg 蔗糖處理下,雖然武運粳23總根長相對II優084來說有明顯的響應優勢,但其根系平均直徑卻偏低(圖4D),導致雖然表面積仍有較大趨勢但根系體積卻與II優084相當。反過來說,雖然兩者的根體積相同,但武運粳23能分配更多的能量用于生長根系的長度而非讓根變得粗壯。這樣使根系變得細長,增加根系吸收面積而更有利于養分的吸收和生物量的積累。
我們之前的田間試驗結果顯示,在水稻的營養生長后期以及生殖生長期,高應答品種II優084無論是地上部還是根系生物量對高CO2的響應均明顯高于低應答品種武運粳23,而本文中試驗結果顯示低應答品種武運粳23在幼苗期對高CO2和高濃度蔗糖的響應更顯著,說明在水稻生長發育的不同時期,不同品種對高C環境的響應也不盡相同。
根系對植物生物量積累起著重要的作用,根系發育程度的好壞直接影響養分的吸收利用。高CO2條件下水稻根系的生長和形態變化的研究直接關系未來高CO2條件下的糧食安全問題。CO2濃度升高及其與其他環境因素的互作對水稻根系的影響,及不同基因型水稻根系對高CO2的響應差異機制與養分吸收的關系,還有待進一步研究。
4 結論
本文同時運用兩種方法對水稻苗期的高C環境進行模擬,且獲得的結果一致。比較兩個產量對高CO2響應有顯著差異的水稻品種的根系在幼苗時期對高C環境的響應。結果證明,苗期水稻根系對高CO2的響應與氮素供應濃度有關。低氮條件下,產量高應答品種II優084根系響應更顯著;而正常氮和高氮條件下產量低應答品種武運粳23根系對高C的響應更顯著,說明在水稻生長發育的不同時期,不同品種對高C環境的響應也不盡相同。
[1] NOAA/ESRL. Trends in atmospheric carbon dioxide[OL]. https://www.esrl.noaa.gov/gmd/ccgg/trends/global.html. Acces-sed 2018
[2] Ainsworth E A, Long S P. What have we learned from 15 years of free-air CO2enrichment (face)? A meta-analytic review of the responses of photosynthesis, canopy[J]. New Phytologist, 2005, 165(2): 351–371
[3] Bloom A J, Smart D R, Nguyen D T, et al. Nitrogen assimilation and growth of wheat under elevated carbon dioxide[J]. Proceedings of the National Academy of Sciences of the United States of America, 2002, 99(3): 1730–1735
[4] Kim H Y, Lieffering M, Miura S, et al. Growth and nitrogen uptake of CO2-enriched rice under field conditions[J]. New Phytologist, 2001, 150(2): 223–229
[5] Yang L X, Huang J Y, Yang H J, et al. The impact of free-air CO2enrichment (face) and N supply on yield formation of rice crops with large panicle[J]. Field Crops Research, 2006, 98(2/3): 141–150
[6] Zhu C W, Zhu J G, Cao J, et al. Biochemical and molecular characteristics of leaf photosynthesis and relative seed yield of two contrasting rice cultivars in response to elevated [CO2][J]. Journal of Experimental Botany, 2014, 65(20): 6049–6056
[7] Ziska L H, Bunce J A, Caulfield F A. Rising atmospheric carbon dioxide and seed yield of soybean genotypes[J]. Crop Science, 2001, 41(2): 385–391
[8] Ainsworth E A, Rogers A, Nelson R, et al. Testing the “source-sink” hypothesis of down-regulation of photo-synthesis in elevated [CO2] in the field with single gene substitutions in glycine max[J]. Agricultural and Forest Meteorology, 2004, 122(1/2): 85–94
[9] Zhu C, Ziska L, Zhu J, et al. The temporal and species dynamics of photosynthetic acclimation in flag leaves of rice (oryza sativa) and wheat (triticum aestivum) under elevated carbon dioxide[J]. Physiol. Plant, 2012, 145(3): 395–405
[10] 卞景陽, 張文忠, 張佳華, 等. CO2濃度增高對水稻葉片微觀結構的影響[J]. 沈陽農業大學學報, 2013, 44(6): 733–737
[11] Ainsworth E A, Beier C, Calfapietra C, et al. Next generation of elevated [CO2] experiments with crops: A critical investment for feeding the future world[J]. Plant Cell and Environment, 2008, 31(9): 1317–1324
[12] Kim H Y, Lieffering M, Kobayashi K, et al. Effects of free-air CO2enrichment and nitrogen supply on the yield of temperate paddy rice crops[J]. Field Crops Research, 2003, 83(3): 261–270
[13] Liu H J, Yang L X, Wang Y L, et al. Yield formation of CO(2)-enriched hybrid rice cultivar shanyou 63 under fully open-air field conditions[J]. Field Crops Research, 2008, 108(1): 93–100
[14] Long S P, Ainsworth E A, Leakey A D B, et al. Food for thought: Lower-than-expected crop yield stimulation with rising CO2concentrations[J]. Science, 2006, 312(5782): 1918–1921
[15] Long S P, Ainsworth E A, Rogers A, et al. Rising atmospheric carbon dioxide: Plants face the future[J]. Annual Review of Plant Biology, 2004, 55: 591–628
[16] Drake B G, GonzalezMeler M A, Long S P. More efficient plants: A consequence of rising atmospheric CO2[J]? Annual Review of Plant Physiology and Plant Molecular Biology, 1997, 48: 609–639
[17] Jongen M, Fay P, Jones M B. Effects of elevated carbon dioxide and arbuscular mycorrhizal infection on trifolium repens[J]. New Phytologist, 1996, 132(3): 413–423
[18] 陳改蘋, 朱建國, 龐靜, 等. CO2濃度升高對水稻抽穗期根系有關性狀及根碳氮比的影響[J]. 中國水稻科學, 2006, 20(1): 53–57
[19] 陳改蘋, 朱建國, 謝祖彬, 等. 開放式空氣 CO2濃度升高對水稻根系形態的影響[J]. 生態環境, 2005, 14(4): 503–507
[20] 馬永亮, 王開運, 孫卿, 等. 大氣CO2濃度升高對植物根系的影響[J]. 生態學雜志, 2007, 26(10): 1640–1645
[21] 牛耀芳, 宗曉波, 都韶婷, 等. 大氣CO2濃度升高對植物根系形態的影響及其調控機理[J]. 植物營養與肥料學報, 2011, 17(1): 240–246
[22] Yang L X, Wang Y L, Kobayashi K, et al. Seasonal changes in the effects of free-air CO2enrichment (FACE) on growth, morphology and physiology of rice root at three levels of nitrogen fertilization[J]. Global Change Biology, 2008, 14(8): 1844–1853
[23] Wu J J, Kronzucker H J, Shi W M. Dynamic analysis of the impact of free-air CO2enrichment (FACE) on biomass and N uptake in two contrasting genotypes of rice[J]. Functional Plant Biology, 2018, 45(7): 696–704
[24] Zhu C W, Xu X, Wang D, et al. An indica rice genotype showed a similar yield enhancement to that of hybrid rice under free air carbon dioxide enrichment[J]. Scientific Reports, 2015, 5(12710). DOI: 10.1038/srep15312
[25] 孫會峰, 朱建國, 謝祖彬, 等. 稻田CO2排放對大氣CO2濃度升高的響應[J]. 土壤, 2012, 44(6): 933–940
[26] 王亮, 朱建國, 曾青, 等. 大氣CO2濃度升高對水稻氮代謝影響的研究進展[J]. 土壤, 2010, 42(3): 344–351
[27] 武慧斌, 宋正國, 沈躍, 等. 水稻根系生長發育對CO2濃度升高的響應及其品種間的差異[J]. 生態環境學報, 2014, 23(3): 439–443
Response of Root Morphology in Seedling Stage of Different Rice Cultavars to High Carbon Condition
WU Jingjing1,2, SHI Weiming1*
(1 State Key Laboratory of Soil and Sustainable Agriculture, Institute of Soil Science, Chinese Academy of Sciences, Nanjing 210008, China; 2 University of Chinese Academy of Sciences, Beijing 100049, China)
Two rice cultivars, Wuyunjing23 (WYJ) and IIyou084 (IIY), have a different response to elevated [CO2] in their seed yield. It is not clear how their seedling root respond to high [CO2]. In this study, hydroponic culture and agar plate culture were utilized to explore the difference in WYJ and IIY response to elevated C condition in seedling root morphology. The results of hydroponics experiment showed that the elevated [CO2] increased shoot biomass of IIY by 28.5% under low nitrogen condition, and the dry weight of root was significantly responsive to high [CO2] at seedling stage. The number of adventitious roots was not increased significantly, and the total root length played a more important role in root biomass response. The total root length of IIY increased by 26.3% under high [CO2], and the root length in both levels of root diameter had high response. The response of shoot and root biomass of WYJ was not significant under low nitrogen level, while it is not the case under normal and high nitrogen levels. Under normal nitrogen condition, the response of WYJ shoots to high [CO2] was not significant, while its root biomass was increased significantly by high [CO2]. The number of adventitious roots was increased by 25.8%, while the total root length was increased by 45.0%. The root length in both levels of root diameter had high response. IIY did not respond significantly under normal nitrogen. At the high nitrogen level, the shoots biomass of WYJ was increased by 35.5% under high [CO2], the root biomass was increased by 80.3%, the number of adventitious roots was increased by 38.5%, and the root mean diameter was increased by 16.7%. The total root length had no response to elevated [CO2], and the biomass of IIY had no significant difference under high nitrogen condition. At the same time, WYJ showed higher response in the root surface area and volume to high [CO2] than IIY under normal nitrogen and high nitrogen conditions. The results of agar plate culture test were consistent with the results of water culture test, and response of WYJ root morphology to high concentration of sucrose was higher than that of IIY. The results showed that the different response to high C condition between cultivars would not change with different culture conditions. Different from the response in middle and late growth stage, response of seedlings root biomass and morphology to high [CO2] was more significant in WYJ than in IIY at normal nitrogen level, which shows that the response of rice yield are not always consistent with the response of seedling growth. The high response of seedling growth may not reflect the high response of later yield.
High C condition; Culture; Cultivars; Root morphology
國家自然科學基金項目(31430095)資助。
wmshi@issas.ac.cn)
吳晶晶(1989—),女,山西臨汾人,博士研究生,主要從事水稻分子生理研究。E-mail: jjwu@issas.ac.cn
S511
A
10.13758/j.cnki.tr.2019.06.002
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
