2株副溶血弧菌不同鹽度下致病性和毒力基因差異分析
2020-01-09 01:59:40馬清揚房文紅
海洋漁業 2019年6期
黃 倞,王 元,馬清揚,英 娜,趙 姝,吳 越,房文紅
(1.中國水產科學研究院東海水產研究所,農業部東海漁業資源開發利用重點實驗室,上海 200090;2.上海海洋大學水產與生命學院,上海 201306)
凡納濱對蝦(Litopenaeus vannamei)對鹽度適應能力強,不僅可以在海水中養殖,經淡化后也可以在淡水中養殖[1],2017年我國淡水養殖凡納濱對蝦產量達到59.1萬 t,占其養殖總產量35.5%。然而隨著養殖規模的擴大,病害成為制約凡納濱對蝦養殖的重要因素之一,其中由副溶血弧菌(Vibrio parahaemolyticus)引起的急性肝胰腺壞死癥,給對蝦養殖造成巨大的經濟損失[2]。副溶血弧菌是一種廣泛存在于海水、半咸水環境中的革蘭氏陰性嗜鹽菌,不僅感染魚蝦貝等水生動物[3],給水產養殖造成極大損失,而且也是重要的食源性病原菌,可導致腹瀉、嘔吐等急性腸胃炎[4]。副溶血弧菌可產生耐熱性直接溶血素(TDH)、TDH相關溶血素(TRH)和不耐熱性溶血素(TLH)3種重要溶血素,其在感染致病過程中發揮重要作用[5]。研究發現,Ⅲ型分泌系統(T3SS)和Ⅵ型分泌系統(T6SS)是與副溶血弧菌致病性密切相關的毒力因子[6-7]。T3SS主要由蛋白復合物形成跨膜分泌管道,似針狀結構刺入宿主細胞將效應蛋白直接注入細胞質中,發揮細胞毒性[7]。副溶血弧菌兩條染色體上各有一套T3SS系統,在染色體Ⅰ上的命名為T3SS1,在染色體Ⅱ上的命名為T3SS2。T3SS1系統由結構蛋白 (如 VscA1-VscY1、VcrD1、VcrG1、VcrR1、VcrV1)和效應蛋白 (VopQ、VopR、VopS和VPA0450等)構成,主要發揮對宿主細胞的細胞毒性、介導宿主細胞自噬凋亡等作用[8-9]。T3SS2在染色體II的毒力島上,由類似T3SS1的結構蛋白和7個效應蛋白(VopA/P、VopL、VopC、VopT、VopV、VopZ和VPA1380)構成,主要引起細菌腸毒性和一定的細胞毒性[10-11]、致病性副溶血弧菌不僅出現在海水養殖動物中,在淡水養殖的凡納濱對蝦也有檢出[12],但針對不同鹽度環境中的副溶血弧菌毒力的研究鮮有報道。本研究選取了2株不同鹽度養殖環境中分離的副溶血弧菌,比較2株副溶血弧菌在不同鹽度下的致病性、毒力基因攜帶情況,探索了鹽度對它們的毒力基因表達的影響,以期為不同鹽度下副溶血弧菌毒力機制研究提供理論依據。
1 材料與方法
1.1 菌株來源
實驗用副溶血弧菌海水菌株383分離自海南瓊海海水養殖發病凡納濱對蝦,淡水菌株V9分離自上海奉賢淡水養殖發病凡納濱對蝦,鑒定后保藏于本實驗室。細菌分離采用TCBS選擇性培養基,采用gyrB[13]和 16SrDNA[13]基因對分離細菌進行分子鑒定,生化鑒定結果參照《常見細菌系統鑒定手冊》。
1.2 實驗試劑
TCBS培養基購自青島海博公司;胰蛋白胨和酵母提取物購自Oxoid公司;細菌DNA和RNA提取試劑盒分別購自上海生工生物工程股份有限公司和天根公司;逆轉錄試劑盒和熒光定量PCR試劑盒購自Takara公司;非發酵G-桿菌鑒定試劑盒購自梅里埃公司。基因測序由上海瑞迪生物公司完成。
LB液體培養基配方:稱取胰蛋白胨10 g,酵母提取物5 g,加純水定容至1 L,氯化鈉隨鹽度變化計算添加量。
1.3 不同鹽度下弧菌生長速率實驗
從TCBS平板挑取菌落,用無菌生理鹽水制成菌懸液。移取菌懸液分別接入液體培養基,使2株弧菌懸濁度相近,液體培養基鹽度分別為2、10、20、30、40、50、60、70和80,設2個平行。于 28℃、轉速 180 r·min-1下培養。取 0、4、12、24、48 h的菌液在酶標儀(Spark 10 m,Tecan)上測定OD600時的吸光值。
1.4 弧菌感染實驗
實驗在東海所實驗基地進行,感染用凡納濱對蝦為實驗基地養殖,體長5~7 cm,體質量4~5 g。實驗在30 L塑料桶里進行,每個桶放養對蝦10尾,海水鹽度22,水溫控制在24~26℃,充氣增氧,投喂適量顆粒飼料,經3 d適應養殖后用于感染實驗。
菌株383和菌株V9分別接種于鹽度25和2的液體培養基中,28℃恒溫培養16 h,配置5個細菌攻毒濃度梯度,采用肌肉注射方式進行攻毒,注射量50μL·尾-1,對照組注射生理鹽水和煮沸滅活的菌體。攻毒組與對照組各設置2個平行組,每天記錄死亡量,連續觀察4 d。半致死濃度計算采用 Bliss法[34]。
1.5 弧菌毒力基因檢測
采用 PCR(Mastercycler pro,Eppendorf)對 2株副溶血弧菌的毒力基因進行檢測。檢測的毒力 基 因 有tdh[14]、trh[14]、tlh[15]、T3SS1[16]、T3SS2[16]、pirA[17]、pirB[17]、orf8[18]、toxR/S[19],其方法和引物參照相應毒力基因的文獻,引物由上海生工生物工程股份有限公司合成。
采用熒光定量 PCR(QuantStudio,ABI)檢測vcrD1[20]、vopS[20]、vopD1[20]和pirA[17]基因 表 達量,內參基因為pvuA[21],定量PCR引物由上海生工生物工程股份有限公司合成。
1.6 鹽度對副溶血弧菌T3SS1毒力基因表達的影響
1.6.1 弧菌培養
將2株副溶血弧菌分別接種于LB液體培養基中,28℃搖床過夜培養,將過夜培養的細菌以1%的比例分別接種于鹽度2、10、20和30的LB液體培養基,于180 r·min-1搖床培養。根據生長曲線估計培養時間,在OD600達到0.6時,取菌液4℃離心取沉淀。每個試驗組各設2個平行。
1.6.2 總 RNA提取與反轉錄
總RNA提取:使用TIANGEN細菌RNA提取試劑盒提取副溶血弧菌總RNA,操作在紫外線滅菌的獨立RNA實驗室中進行。采用1%瓊脂糖凝膠電泳檢測總RNA的完整性,同時用微量紫外分光光度計(ND5000,百泰克)測定RNA濃度。
反轉錄:采用DNA水解酶去除總RNA中殘留的DNA;采用Takara反轉錄試劑盒將RNA反轉錄成單鏈cDNA,反轉錄產物cDNA分裝后置于-80℃保存。
1.6.3 熒光定量PCR反應體系與條件
反應體系為10μL,其中cDNA 2μL,2×TB Green Premix ExTaqⅡ 5μL,ROX Reference DyeⅡ(50×)0.2μL,10μmol·L-1上下游引物各0.4μL,無菌雙蒸水 3μL。反應條件:95℃ 3 min,預變性:95℃ 10 s,55℃ 30 s,72℃ 30 s,40個循環,72℃實時檢測熒光信號,同時進行ROX值校正。最后進行RT-PCR溶解曲線分析。
2 結果與分析
2.1 鹽度對兩株副溶血弧菌生長速率的影響
分析2株弧菌在不同鹽度培養基下的生長速率(圖1),可以看出:海水菌株383在低鹽度下(培養基鹽度≤30時)生長速率快,培養12 h即接近生長穩定期;高鹽度(鹽度>30)時,其生長速率下降,24~48 h達到生長穩定期。淡水菌株V9除鹽度80外,在其他鹽度下都是在24 h達到生長穩定期。海水菌株383可以在鹽度高達80的培養基中生長,而淡水菌株V9在此鹽度下幾乎不生長;2株弧菌生長達到生長穩定期時,淡水菌株V9濃度明顯高于海水菌株383。
2.2 弧菌回感實驗結果
感染發病的凡納濱對蝦肝胰腺顏色變淡呈淺白色,萎縮變小,邊緣模糊。2株副溶血弧菌感染凡納濱對蝦48 h死亡情況見圖2,經計算,海水菌株383和淡水菌株V9的48 h半致死濃度(LD50)分別為1.73×103cfu·mL-1和1.32×105cfu·mL-1,由此可見海水菌株383的毒力明顯高于淡水菌株V9。在本實驗中,注射滅活菌液和生理鹽水對照組未見對蝦死亡。
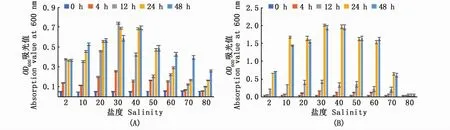
圖1 副溶血弧菌在不同鹽度下生長速率(n=2)Fig.1 Growth rate of Vibrio parahaemolyticus at different salinities
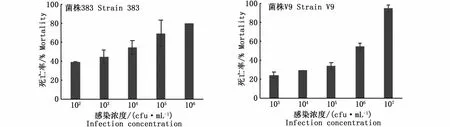
圖2 副溶血弧菌肌肉注射感染凡納濱對蝦48 h死亡率(n=2)Fig.2 48 h mortality rate of Litopenaeus vannamei infected by Vibrio parahaemolyticus after intramuscular injection
2.3 弧菌毒力基因攜帶情況
采用PCR方法對2株副溶血弧菌分別進行tlh、tdh、trh、T3SS1、T3SS2、pirA、pirB、toxR/S、orf8基因檢測,檢測結果見表1。2株副溶血弧菌tlh、T3SS1、pirA和pirB為陽性,tdh、trh、T3SS2、toxR/S和orf8為陰性。
2.4 鹽度對2株弧菌毒力基因表達的影響
對檢測結果為陽性的T3SS1基因和pirA基因進行鹽度影響的基因表達分析。
不同鹽度對2株副溶血弧菌4個毒力基因表達的影響見圖3。從4個毒力基因來看,vcrD1表達量較高,其次是pirA,vopD1表達量最低。進一步分析,不同菌株以及鹽度對毒力基因表達有較大的影響。海水菌株383的pirA在不同鹽度均有表達,在鹽度20時表達量最高;而淡水菌株V9的pirA僅在鹽度2時有表達,其表達量分別為菌株383在鹽度2和鹽度20下的54.7%和9.3%。2株菌株vopD1基因在不同鹽度下都有表達,均在鹽度2時表達量最高。淡水菌株V9的vopS基因在不同鹽度都有表達,而海水菌株383在鹽度10、20和30有表達,其表達量均低于淡水菌株。在鹽度20時海水菌株383的vcrD1基因超高表達,是其他鹽度的8.5~17.5倍;淡水菌株V9在鹽度2時有表達,在其他鹽度下表達量較低。從2個菌株來看,差別最大的毒力基因是pirA和vcrD1。
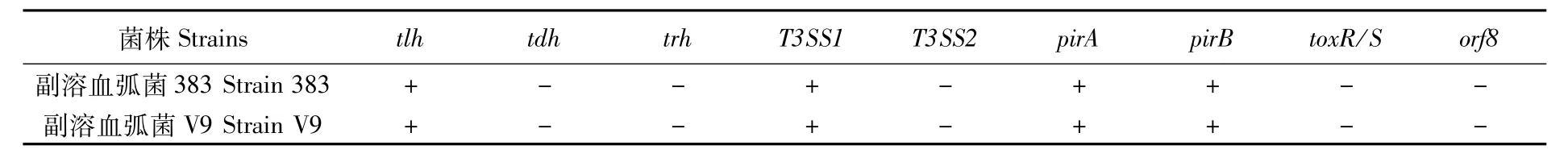
表1 2株副溶血弧菌毒力基因檢測結果Tab.1 Detection results of virulence genes of two strains of Vibrio parahaemolyticus
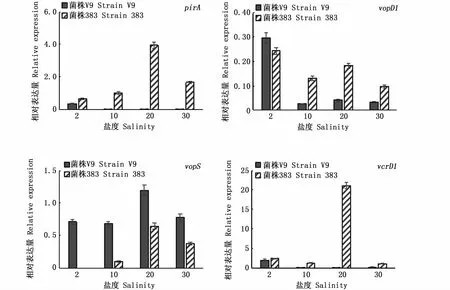
圖3 不同鹽度下2株副溶血弧菌毒力基因表達量(n=2)Fig.3 Expression of several virulence genes in Vibrio parahaemolyticus at different salinities
3 討論
3.1 淡水養殖凡納濱對蝦為副溶血弧菌進入淡水環境帶來風險
副溶血弧菌為嗜鹽菌,多出現在海水和半咸水環境,在淡水及淡水產品中檢出率較低[12]。為了提高淡水養殖凡納濱對蝦成活率,在蝦苗放養前往往向淡水中添加海水鹽提高水的鹽度,因此部分弧菌能得以生存且存在致病性[23]。本研究中的淡水菌株正是從該養殖方式下的一例病蝦內分離獲得的病原菌。從本研究結果來看,淡水菌株適應了低鹽度環境后在低鹽度下生長速率快于海水菌株,同時對高鹽度的耐受力有所下降。由此可見,淡水中養殖對蝦為副溶血弧菌侵入淡水養殖品種提供了輸入途徑,咸化的淡水為副溶血弧菌提供了生存條件。淡水養殖海水對蝦,可以導致病原向新的水體輸入,應引起一定的重視。
3.2 鹽度影響副溶血弧菌毒力基因的表達
環境因素如溫度、鹽度、酸堿度等都會影響毒力基因的表達,且不同的環境條件對不同毒力基因表達的影響是不一致的[20,23-24]。弧菌是條件致病菌,在外界環境條件穩定時一般不會引起發病,但一些環境因子的改變,例如溫度、鹽度等均可以調控其致病性相關蛋白的表達,從而影響細菌的致病性[23-25]。溶藻弧菌(V.alginolyticus)在溫度28℃、鹽度20、培養基初始pH為7時,其T3SS轉運蛋白VscO基因表達量最高[25]。遲緩愛德華菌在25℃時分泌的EvpA、EvpC、EseB和EseD蛋白明顯多于37℃時;注射感染37℃培養的遲緩愛德華菌,90%的線足鱸(Trichogaster trichopterus)成活,而注射25℃培養的遲緩愛德華菌,則有70%線足鱸死亡[26]。雖然該菌在37℃條件下生長快,但其T3SS輸送器蛋白表達較低;在28℃條件下T3SS輸送器蛋白表達最高,而在20℃條件下則沒有檢測到T3SS輸送器蛋白的表達。同等溫度下,中性和堿性環境比酸性環境更適合細菌生長和T3SS輸送器蛋白的表達[27]。本研究中,鹽度對副溶血弧菌pirA基因和T3SS1內膜蛋白vcrD1基因表達有較大的影響,不僅存在菌株間的差異,而且同一菌株的不同毒力基因受到的影響也是不同的。鹽度對副溶血弧菌毒力基因表達的影響機制還有待進一步研究,以為揭示副溶血弧菌致病機制提供理論依據。
3.3 副溶血弧菌海水菌株和淡水菌株的毒力分析
pirA和pirB最早是在副溶血弧菌中發現的、由質粒介導的重要毒力因子[28],由其引起的急性肝胰腺壞死病(acute hepatopancreatic necrosis disease,AHPND)對全球的對蝦產業造成了巨大的損失[28]。患病對蝦肝胰腺顏色變淡呈淡白色,部分患病對蝦肝胰腺明顯萎縮,質地變硬,腸胃變空。2018年有報道,pirA和pirB毒力基因出現在多種弧菌中,均能引起對蝦AHPND[29]。
T3SS作為副溶血弧菌的主要毒力因子之一,通過傳遞與致病有關的毒性因子來發揮病原菌的毒性,而鹽度是影響細菌基因表達以及其他生命活動的關鍵生態因子。副溶血弧菌T3SS1主要是對宿主細胞產生細胞毒性,介導宿主細胞的自體吞噬作用,最后導致細胞死亡。T3SS1在感染過程中與宿主細胞的凋亡有關,對多種細胞系如 HeLa細胞、Raw細胞均具有細胞毒性[1,30]。T3SS2又分為T3SS2α和T3SS2β兩種,且兩者不能存在于同一株菌中[31];T3SS2β僅存在于 KP+分離株中,與兔回腸袢結扎實驗模型中的腸毒性有關[32]。本文2株副溶血弧菌只檢測到T3SS1,未能檢測到T3SS2。
vcrD1編碼的內膜蛋白是組成T3SS1分泌裝置基體的一部分,在此基礎上針管狀結構蛋白才能繼續生長。vopD1編碼的轉運蛋白能在宿主細胞膜表面形成孔洞,效應蛋白沿該通道進入宿主細胞,具有HeLa細胞毒性和接觸溶血活性,轉運蛋白vopD1高表達有利于T3SS分泌通道的完善,釋放分泌蛋白,表現細菌毒性[33]。vopS編碼的效應蛋白可導致細胞變圓并通過抑制RhoB家族的鳥苷三磷酸酶導致肌動蛋白細胞骨架崩潰,并且vopS可以阻止Rho家族的鳥苷三磷酸酶與下游的效應蛋白反應,抑制感染細胞內肌動蛋白的重排[34]。
比較分析本研究中2菌株的毒力與pirA、T3SS1效應蛋白基因表達量之間的關系,發現兩株菌的pirA和vcrD1表達量相差10倍左右,結合半致死濃度(LD50),兩菌株的LD50相差近2個數量級。因此認為,2株副溶血弧菌的毒力強弱可能與pirA和vcrD1的表達量有關。
