橡膠樹白粉菌(HO-73)啟動子WY172 不同長度片段的克隆及表達活性分析
2020-01-14 06:59:04殷金瑤王義徐良向朱利王晨劉文波繆衛國
生物技術通報 2020年1期
殷金瑤 王義 徐良向 朱利 王晨 劉文波 繆衛國
(海南大學植物保護學院 熱帶農林生物災害綠色防控教育部重點實驗室,海口 570228)
啟動子是一段位于結構基因5′端上游的DNA序列,提供RNA 聚合酶識別和結合位點,是調控基因表達強弱的重要元件,通過參與基因的轉錄環節來調控基因表達的水平、部位及方式[1-2]。研究表明,通過自身啟動子在植物中對外源基因進行功能表達,為植物生物技術中有益轉基因的應用提供了可能,如提高植物脅迫耐受性等[3]。
目前,最經典的啟動子是花椰菜花葉病毒(CaMV)35S 啟動子[4]、玉米泛素(Ubiquitin)啟動子[5]和水稻肌動蛋白(ACT1)啟動子[6]等。但隨著應用不斷深入,上述幾個啟動子的缺陷也不斷暴露出來,如因高效表達需要產生的大量能耗會對植物生長產生影響,引起植物生長緩慢[7]。同時因其表達具有持續性,不隨時間和空間的變化而變化,在植物體內表達不具有特異性[8],因而人們開始尋求更加高效、優質的啟動子。
有研究表明,使用絲狀真菌中的強啟動子可以高效啟動目的基因的轉錄。在絲狀真菌中利用自身啟動子起始轉錄外源基因的轉錄效率通常很高,但是一些外源蛋白的產量遠遠低于內源蛋白的產量[9]。有文獻報道,啟動子活性、翻譯效率、蛋白糖基化、目標基因的拷貝數以及菌株自身蛋白酶的降解能力等均會影響蛋白的產量,其中以啟動子轉錄水平的高低影響最為關鍵,使用強啟動子可以實現目的基因的高效表達[10-11]。目前已報道的絲狀真菌啟動子主要是曲霉葡糖淀粉酶基因(glaA)啟動子,構巢曲霉的3-磷酸甘油醛脫氫酶基因(gpdA)啟動子,米曲霉的a-淀酶基因(amyB)啟動子,瑞氏木霉纖維二糖水解酶基因(cbh1)啟動子[12]。
橡膠樹白粉菌(Oidium heveae)引起的橡膠樹白粉病是橡膠生產中重要病害之一[13]。在前期研究中,我們首次報道了橡膠樹白粉菌基因組[14],構建了適于快速檢測橡膠樹白粉菌菌株HO-73 啟動子的探針載體[15],并通過將啟動子與CaMV35S 啟動子及ACT1 啟動子的活性比較,初步鑒定了WY7[16]、WY51[17]、WY193[18]等能夠用于調控雙子葉植物和單子葉植物中的外源目的基因表達的橡膠樹白粉菌內源啟動子。本研究中通過對其基因組進行分析,預測篩選了一些疑似內源啟動子的核心區域,并通過克隆得到了其中1 個疑似新的內源啟動子WY172(BankIt2158599,另文發表)。本研究以WY172 上游2K 序列為研究對象,通過利用生物信息學軟件對其進行檢索并預測轉錄因子結合位點,通過構建不同長度疑似具有啟動子活性片段的表達載體,觀察報告基因的表達情況,探討WY172 及4 個不同長度缺失片段的表達活性,旨在尋求更好的可服務于基因工程、應用于生產實際的優秀啟動子。
1 材料與方法
1.1 材料
橡膠樹白粉菌菌株HO-73 篩選、保存于本實驗室,培養于橡膠苗中感品種熱研7-33-97[19]上,培養室內溫度為25℃恒溫,光周期為14 h/10 h(光/暗),濕度范圍88%左右。模式生物三生煙(Nicotiana tabacumcv.Xanthi nc)作為研究對象,培養于上述培養室中。
1.2 方法
1.2.1 WY172 啟動子生物信息學分析 Promoter Scan 預測橡膠樹白粉菌全基因組DNA,得到若干橡膠樹白粉菌疑似內源啟動子的核心區域,將其中一個命名為WY172(BankIt2158599),并利用在線數據庫EPD 確認所驗證的啟動子是否為未報道的新啟動子。利用在線數據庫PROMO 對WY172 上游2K序列進行轉錄因子預測分析,利用在線生物信息學軟件JASPAR 檢索相應的轉錄因子結合位點,根據轉錄因子結合位點在序列上的分布位置對WY172上游2K 序列進行漸變缺失突變,用引物設計軟件Primer Premier 5 設計5′端不同缺失的上游引物和3′端下游引物。同時通過plantCARE 軟件對WY172 啟動子上游2K 序列的作用元件進行分析。
1.2.2 WY172 啟動子及不同長度片段的克隆 提取橡膠樹白粉菌基因組DNA(OMEGA,D3390),以此為模板,通過上游特異引物172F、172Q1F、172Q2F、172Q3F、172QF 和共同的下游引物172NR進行PCR 擴增,獲得5 種不同長度的啟動子片段。
將PCR 產物經1.0%的瓊脂糖凝膠電泳檢測并切膠回收。將回收產物與T-easy 載體連接,進行TA克隆,并轉化Trans5α 感受態細胞,涂布于Amp 抗性的LB固體培養基(100 μg/mL),37℃過夜培養16 h。挑取單菌落于Amp 抗性的LB 液體培養基中,37℃搖床震蕩培養12 h 后進行菌液PCR 驗證,驗證正確的陽性克隆菌株送華大生物公司測序。測序正確的菌株用80%的甘油保菌。
1.2.3 WY172 啟動子及不同長度片段瞬時表達載體的構建 選擇HindIII/BamH I 分別酶切并回收pBI121 載體和1.2.2 中的含不同長度片段的克隆載體,進行連接,并轉化Trans5α 感受態細胞,涂布于Kan 抗性的LB 固體培養基(100 μg/mL),37℃培養16 h。挑取單菌落于Kan 抗性的LB 液體培養基中,37℃搖床震蕩培過夜后進行菌液PCR 驗證,正確的陽性克隆菌株送華大生物公司測序,測序正確的菌株用80%的甘油保菌。
1.2.4 三親雜交法轉化農桿菌菌株 挑選含有目的基因的大腸桿菌菌株,在含有50 μg/mL 卡那霉素的LB 液體培養基中37℃震蕩培養(帶有CaMV35S 啟動子的pBI121 載體作為陽性對照),至OD600nm為0.5時,與LBA4404、HB101 感受態等體積混勻,涂布于不含任何抗生素的LB 固體培養基上,28℃過夜培養。用接種針將長出的菌落轉移到含有100 μg/mL利福霉素和50 μg/mL 卡那霉素的固體LB 培養基上,28℃培養3-4 d,長出單菌落。將長出的單菌落再次轉移到含有100 μg/mL 利福霉素和50 μg/mL 卡那霉素的LB 固體培養基上,挑選單斑在含有50 μg/mL卡那霉素和100 μg/mL 利福霉素的LB 液體培養基中37℃震蕩培養,進行菌液PCR 檢測,正確的陽性克隆菌株送華大生物公司測序,測序正確的菌株用80%的甘油保菌。
1.2.5 GUS 染色 利用農桿菌介導的轉化法(ATMT)侵染三生煙,將侵染后的煙草葉盤28℃共培養2 d后取樣,每個片段啟動子取6 片葉盤浸泡于GUS 染色液[20]中,37℃過夜培養,后將樣品轉入70%乙醇中脫色至葉片組織無色,觀察。
1.2.6 GUS 酶活性測定 取各樣品0.1 g 液氮研磨,提取植物總蛋白(CWBIO,Plant Protein Extraction Reagent),-80℃保存。GUS 蛋白濃度的測定采用BCA 法(Solarbio,BCA Protein Assay),使用多功能酶標儀Infinie 200 PRO 測定樣品在562 nm 處的吸光值,最后根據標準曲線計算樣品蛋白濃度。采用Jefferson 等[21]報道的方法測定GUS 蛋白活性,結果以生成的4-甲基傘形酮(4-methylumbelliferon,4-MU)的量與總蛋白的量和時間的比值(pmol 4-MUG/mg protein/min)來表示。實驗設置3 次重復。最后使用SPSS 軟件統計結果。
2 結果
2.1 WY172啟動子生物信息學分析
在線數據庫EPD 確認未發現與WY172 同源的啟動子。對轉錄因子的預測分析結果顯示WY172啟動子上游2K 序列上共有轉錄因子結合位點17 個(表1),運用IBS 軟件繪制基因圖譜(圖1),直觀地展示出各個轉錄因子結合位點在基因上的位置,為后續缺失突變位置的確定提供參考。根據轉錄因子結合位點在序列上的分布位置,以不切斷轉錄因子結合位點為原則,設計了4 個缺失突變片段,分別 為WY172Q(2250 bp)、WY172Q1(1551 bp)、WY172Q2(1281 bp) 和WY172Q3(871 bp)( 圖2)。結合前期研究得到的WY172 啟動子(251 bp),通過Primer Premier 5 軟件設計了6 個引物(表2)。PlantCARE 軟件分析結果(表3)顯示,其上游2K序列含有豐富的作用元件,包括光響應元件、脫落酸響應元件、生長素響應元件、茉莉酸甲酯響應元件、創傷誘導元件、厭氧感應元件、參與柵欄葉肉細胞分化的元件及玉米蛋白代謝調節的順式作用元件等。
2.2 WY172啟動子及不同長度片段的克隆
通過PCR 及產物膠回收,成功獲得5 個帶有酶切位點的不同長度片段,并構建了Teasy-WY172、Teasy-WY172Q、Teasy-WY172Q1、Teasy-WY172Q2、Teasy-WY172Q3 共5 個載體,菌液PCR 結果顯示出與目的片段大小基本一致的條帶(WY172Q 為2300 bp 左右,WY172Q1 為1500 bp 左 右,WY172Q2 為1300 bp 左 右,WY172Q3 為800 bp 左 右,WY172為250 bp 左右)。并經過華大生物公司測序及NCBI比對結果顯示序列相似性為100%,說明成功克隆了WY172 啟動子及不同長度片段。

表1 WY172 啟動子上游2K 序列含有的轉錄因子結合位點
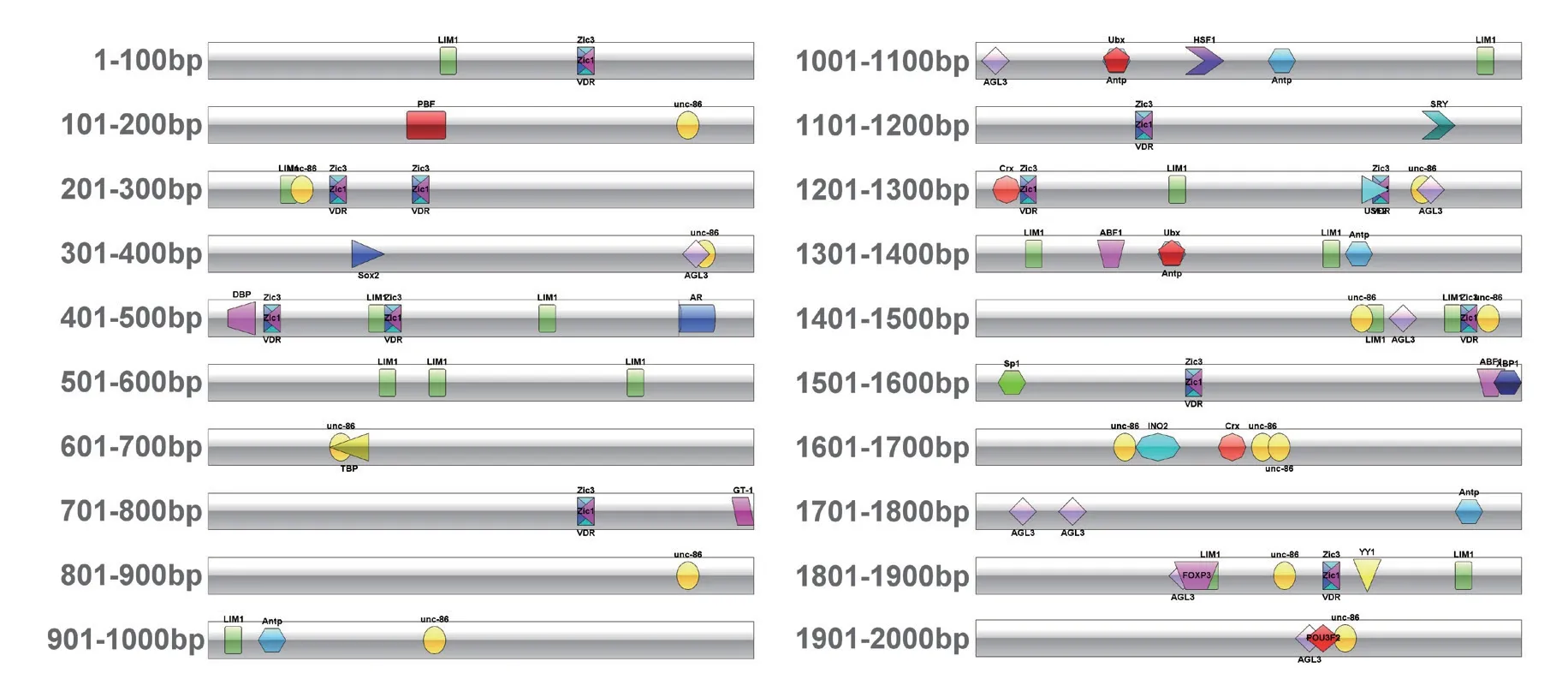
圖1 WY172 啟動子上游2K 序列轉錄因子分布圖

表2 引物序列
2.3 WY172啟動子及不同長度片段瞬時表達載體的構建
陽性克隆菌株的菌液PCR 結果顯示獲得與目的條帶大小一致的片段,華大公司測序結果經過比對顯示DNA 相似性為100%,獲得了重組的植物表達 載 體pBI121-WY172、pBI121-WY172Q、pBI121-WY172Q1、pBI121-WY172Q2、pBI121-WY172Q3(以pBI121 載體作為陽性對照),同時通過Vector NT1繪制了植物表達載體示意圖(圖2-A),并利用IBS軟件繪制了WY172 啟動子不同長度片段特征示意圖(圖2-C)。將5 個重組植物表達載體通過三親雜交法轉化農桿菌,菌液PCR 檢測結果顯示出與目的基因大小一致的特異片段(圖2-B)(WY172Q 為2300 bp 左右,WY172Q1 為1500 bp 左右,WY172Q2 為1300 bp 左 右,WY172Q3 為800 bp 左 右,WY172為250 bp 左右),經過華大生物公司測序及NCBI 比對結果顯示序列相似性為100%,說明獲得了含有各缺失片段啟動子的農桿菌菌株。

表3 WY172 啟動子上游2K 序列含有的作用元件

圖2 不同長度啟動子片段及載體構建
2.4 GUS蛋白染色
GUS 染色結果(圖3)顯示,所有菌株瞬時表達的葉盤都成功染出了藍色,其中陽性對照菌株(pBI121 載體)侵染的葉盤染色最淺,而WY172 不同片段菌株侵染的葉盤染色深度均強于陽性對照。pBI121-WY172Q3 葉盤染色最深,顏色由深至淺排列是:pBI121-WY172Q3> pBI121-WY172Q2> pBI121-WY172Q1> pBI121-WY172> pBI121-WY172Q,總體呈現隨長度增長GUS 染色顏色逐漸變淺的趨勢,但WY172Q3 片段出現藍色顯著加深現象。結果表明,WY172 啟動子的5 個不同長度片段均能啟動GUS基因的表達,其中WY172Q3 的表達活性最強。

圖3 不同長度啟動子片段煙草瞬時表達的GUS 染色
2.5 GUS酶活性檢測
為測定不同長度片段的酶活性,繪制了BSA 標準曲線和4-MU 標準曲線,R2值分別為0.9995 和0.9998,達到了測定要求(圖4)。SPSS 軟件統計酶活性分析結果(圖5)顯示,野生型幾乎檢測不到酶活性,而CaMV35S 啟動子和5 個WY172 不同長度片段啟動子均具有驅動GUS基因表達的酶活性,GUS 酶活性趨勢為:WY172Q3> WY172Q2>WY172Q1> WY172> WY172Q> CaMV35S,總體呈現隨長度的增長活性逐漸遞減趨勢,但在WY172Q3處酶活性出現顯著回升。酶活性檢測結果與GUS 染色結果一致,再次證明WY172 啟動子的5 個不同長度片段均具有啟動子活性,WY172Q3 表達活性最強。
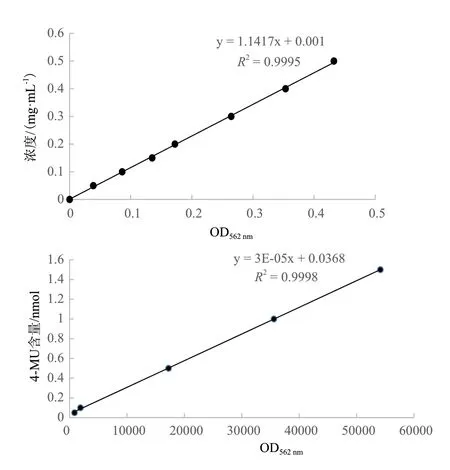
圖4 BSA 標準曲線和4-MU 標準曲線
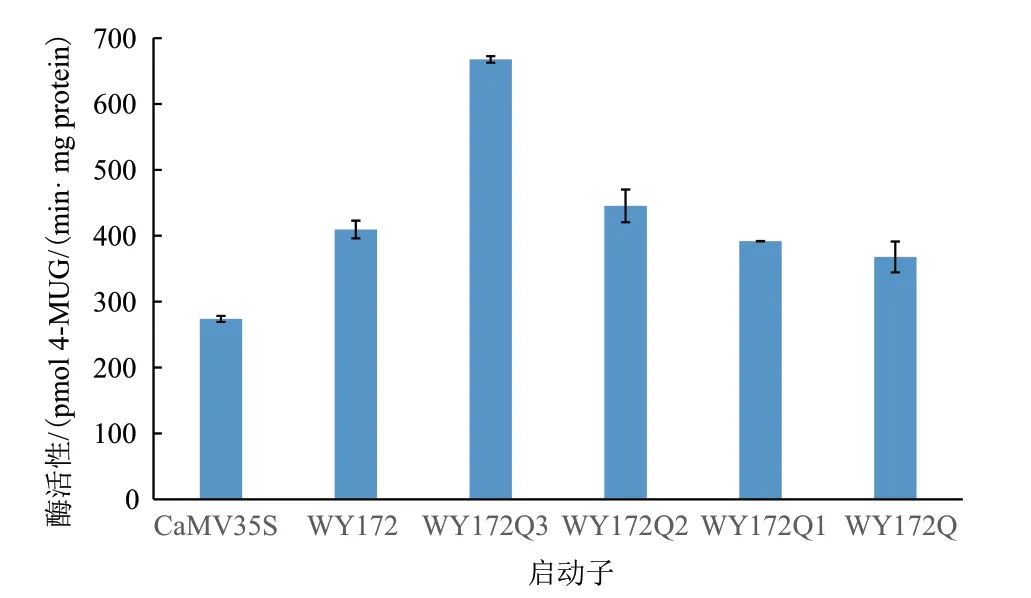
圖5 不同長度啟動子片段煙草瞬時表達的GUS 酶活性
3 討論
通過對不同長度啟動子的表達活性進行分析可以得知啟動子發揮作用的能力[22-23],為充分解讀其對基因的調控表達過程及后續應用奠定基礎。橡膠樹白粉菌是一種專性寄生菌,分子研究嚴重落后,啟動子方面的研究更是少之又少[24]。目前,已見報道的研究多集中于啟動子的篩選和鑒定。本實驗室前期通過不同卡那霉素濃度下啟動子的耐受性大小來篩選強啟動子,并以橡膠樹白粉菌基因組為研究對象,鑒定出多個在單子葉和雙子葉植物中均能表達的強啟動子。在此研究的基礎上,本研究聚焦于WY172 啟動子不同長度片段驅動外源基因表達的能力,以WY172 啟動子上游2K 序列為研究對象,進行漸變缺失突變,分析不同片段驅動GUS基因的表達活性。
本研究經過初步分析發現WY172 啟動子上游2K 序列含有豐富的順式作用元件,其中有參與創傷反應的WUN-motif 作用元件,厭氧感應的ARE 順式作用元件,參與葉肉細胞柵欄組織分化的HD-Zip1元件,參與玉米蛋白代謝調節的O2-site 順式作用元件,以及參與晝夜控制的circadian 順式作用元件等功能作用元件,還有茉莉酸甲酯響應、赤霉素響應、生長素響應等激素作用元件,此外還有低溫響應、光響應元件等非生物因素的作用元件。這表明WY172 啟動子很可能在植物生長的各個階段都能發揮作用,多樣性的元件也為下一步高效表達外源基因的啟動子的開發提供了可能,為WY172 啟動子的進一步研究和開發奠定了基礎。
GUS 染色結果及酶活性檢測表明,在調控GUS基因表達方面,WY172 啟動子的不同長度片段都具有比CaMV35S 啟動子強的啟動子活性。CaMV35S啟動子是目前國際上應用最廣、活性最強的啟動子之一,但隨著研究的深入,該類啟動子在應用上的不足已經顯現地越來越明顯[25]。WY172 啟動子的不同長度片段均具有比CaMV35S 啟動子更強的活性,這為基因工程工具的開發提供另外一個可能性。在后續實驗中將會對WY172 啟動子的具體類型及應用范圍等進行分析,以便為基因工程開發一個更有效的啟動子工具。
順式作用元件的種類和數量的不同,以及轉錄因子結合位點的差異,都有可能影響基因的表達效果。活性分析結果顯示,WY172 不同片段的活性強弱關系為WY172Q3> WY172Q2> WY172Q1>WY172> WY172Q> CaMV35S。WY172Q3 段 啟 動 子表達活性升高可能是由于在251 bp-821 bp 區段含有轉錄激活元件,使得該區段具有顯著增強效應;WY172Q2 段啟動子區段也有增強效應但表達活性低于251 bp-821 bp 段的原因,一個可能是821 bp-1281 bp 區段含有的激活元件種類和數量都少于WY172Q3 段,也可能是821 bp-1281 bp 區段同時含有抑制表達元件,與251 bp-821 bp 區段中的一部分激活元件的功能相抵了;WY172Q1 段啟動子及WY172Q 段啟動子表達活性降低,可能是由于1281 bp-2250 bp 區段含有抑制表達元件,導致了抑制效應。未來,我們將會對具體的抑制、激活元件進行定位及探究,從而研究該啟動子不同情況下的表達量變化。
我們的啟動子來源于絲狀真菌,這類真菌表達外源蛋白的量比較低[26],為實現外源基因的高效表達,通常需要強大的啟動子來驅動基因的轉錄,但目前針對絲狀真菌啟動子的研究與應用比較少,主要是木霉屬(Trichoderma)和黑曲霉屬(Aspergillus niger)的一些啟動子[27]。除此之外,絲狀真菌啟動子大部分都只能應用于自身菌體中外源基因的表達,而來源于橡膠樹白粉菌的WY172 啟動子不僅具有絲狀真菌啟動子的優點,還可以應用于植物,這為其推廣和應用提供了更大的可能。為后續對于其下游基因和調控方式的確定提供方向、奠定基礎,同時從一定程度上也促進了橡膠樹白粉菌的分子系統發育和形態學分析研究工作。
4 結論
本研究克隆了WY172 啟動子及其4 個不同長度的缺失片段,并構建了植物表達載體,通過GUS染色和酶活性檢測,對啟動子表達活性進行了定性和定量分析。證明以上WY172 啟動子5 個不同長度片段均具有啟動子活性,均能驅動GUS基因的表達。GUS 染色結果顯示WY172 啟動子不同長度片段的藍色強度均優于陽性對照CaMV35S 啟動子,其中WY172Q3 驅動GUS基因表達的酶活性最高。且酶活性檢測也與GUS 染色結果相符。不同片段的缺失對表達活性的影響不同,pBI121-WY172Q3 啟動子表達活性最強。
