2018年6月渤海大型水母分布特征*
2020-02-06 06:00:48王朋鵬
海洋與湖沼 2020年1期
王朋鵬 張 芳 孫 松,5① 楊 濤
(1.中國科學院海洋研究所 海洋生態與環境科學重點實驗室 青島 266071;2.青島海洋科學與技術試點國家實驗室 海洋生態與環境科學功能實驗室 青島 266071;3.中國科學院大學 北京 100049;4.中國科學院海洋大科學研究中心 青島266071;5.中國科學院海洋研究所 山東膠州灣海洋生態系統國家野外科學觀測研究站 青島 266071;6.中國水產科學研究院黃海水產研究所 山東省漁業資源與生態環境重點實驗室 青島 266071)
自1990年以來,全球很多海域中水母數量顯著增多(Arai,2001;Purcellet al,2001a,b;丁峰元等,2005),已給全球范圍內的經濟、社會和生態帶來了嚴重影響。我國近海暴發的大型水母種類主要有沙海蜇(Nemopilema nomurai)、霞水母(Cyaneaspp.)、海月水母(Aureliaspp.)、多管水母(Aequoreaspp.)等(Donget al,2010;Zhanget al,2012)。近二十年來,針對近海暴發的大型水母我國科學家在黃、東海開展了大量野外調查工作,在其時空分布及物理驅動機制、種群動態、水母旺發引發的漁業資源效應及生態效應等方面的研究卓有成效(李建生等,2009;王彥濤等,2012;Zhanget al,2012;Sunet al,2015;左濤等,2016a;張芳等,2017)。
我國渤海海域同樣出現水母暴發現象,萊州灣、遼東灣作為我國傳統意義上漁業生物重要的產卵、孵育和索餌場所,大型災害性水母暴發嚴重影響了該海域海洋漁業資源與漁業生產(董婧等,2013;左濤等,2016b);水母暴發在影響漁業資源的同時,更是危及到人類生命安全,浴場水母蜇傷事件的報道層出不窮(董婧等,2013;鄭向榮等,2014b)。因此,我國科學家在渤海海域同樣開展了大量水母自然生態學的研究工作,但這主要以遼東灣大型水母調查研究為主,例如,遼寧省海洋水產科學研究院對遼東灣大型水母的種群變動開展了長期監測,對該海域不同年份各階段大型水母的基礎生物學信息有了清楚的了解,并結合室內生態學實驗,在大型水母生活史、繁殖生物學等方面進行了比較系統的探討(Donget al,2006,2008;王彬等,2010,2012;董婧等,2012,2013;孫明等,2013)。鄭向榮等(2014a,b)、左濤等(2016b)分別對河北近海、萊州灣大型水母的分布特征進行過報道。到目前為止,我國對于渤海海域大型災害水母的監測結果較為分散,具有明顯區域性,缺少同步、整體性地調查,限制了大型水母在渤海種群動態的整體性分析,同時制約了渤海、黃東海大型水母自然生活史的關聯性探討,從而阻礙了對我國近海大型水母自然生活史的整體性理解。
本研究于2018年6月上旬,在渤海海域進行了較大范圍的定點調查,調查船舶為“中漁科102”漁業資源調查船,記錄了渤海內大型水母種類組成、漁獲密度及傘徑大小分布。通過本研究,以期達到如下目的:(1)系統性了解6月份整個渤海海域大型水母的分布格局,并探討渤海各海域水母之間潛在的連通性,推測渤海海域大型水母可能存在的暴發源頭;(2)結合前人研究結果,對渤海與黃東海大型水母進行了對比,探討渤海與黃東海大型水母生長的差異。
1 材料與方法
1.1 調查海域及站位設置
本研究于2018年6月上旬,在渤海海域進行了大型水母種群特征調查,科考船舶為水科院黃海水產研究所“中漁科102”號。調查站位按0.5°×0.5°設定,計劃調查站位60個,因部分站位所在海域水較淺或存在漁業養殖區,因此實際調查站位為47個,且實際走航站位與理論設定的0.5°×0.5°有所不同,詳如圖1所示,拖網時間均為白天。為對比渤海不同海域水母生物學信息是否具有差異,將整個渤海海域劃分為渤海中部、遼東灣、渤海灣、萊州灣四部分海域,詳見圖1。
1.2 樣品采集
水母樣品采用漁業資源調查底拖網(網目:200mm×440目,囊網網目:2mm×287目,網口周長:46.6m,網長:51.7m)采集,各站位拖網時間控制在0.5—1h,部分站位受惡劣天氣、底泥等因素影響拖網時間略短,拖速控制在3kn左右。各站位起網后將所有出現的水母進行收集,在船載實驗室中測定記錄水母種類、個數,并分別測量各類水母傘徑(cm)。海區中大型水母的漁獲量用漁獲密度[CPUE,單位:ind./(net·h)]表示,即每小時每網大型水母的漁獲數量。停船時使用船載CTD測量所在站位表、底層溫度、鹽度。
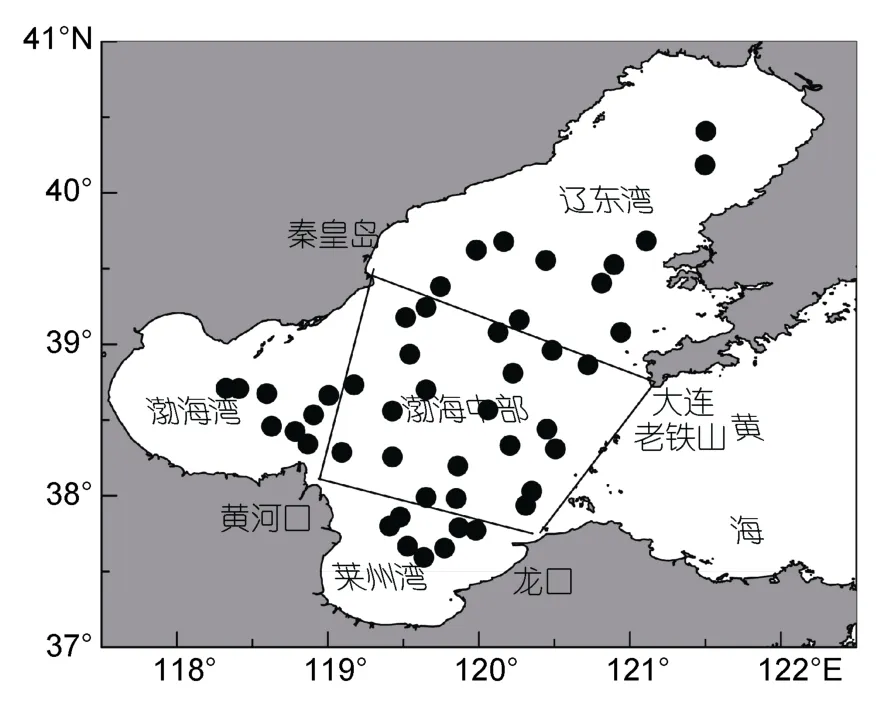
圖1 水母調查站位圖Fig.1 Sampling stations in the Bohai Sea
1.3 數據統計
本研究數據統計、處理所使用軟件為Surfer12、SPSS Statistics19。本文使用Surfer12進行調查站位圖、環境參數分布圖以及水母傘徑與密度分布圖的繪制;分別使用SPSS Statistics19中KolmogorovSmirnov檢驗和Levene檢驗進行渤海各海域(遼東灣、渤海中部、渤海灣、萊州灣)溫鹽數據組內正態分布檢驗和組間方差齊性檢驗,然后利用One-Way ANOVA分析各海域溫鹽的差異顯著性;使用Pearson相關性分析檢驗溫鹽與水母密度之間所存在的相關關系;使用非參數Kruskal-Wallis檢驗用來對比渤海各海域(遼東灣、渤海中部、渤海灣、萊州灣)水母傘徑大小是否具有顯著差異性。P<0.05表征具有顯著差異。
2 結果
2.1 調查海域溫、鹽分布特征
圖2展示了2018年6月渤海海域表、底層溫度、鹽度分布特征。結果顯示,渤海各海域表溫無明顯差異(F=2.780,P=0.052),均值為(19.5±1.4)°C,最高溫度 22.0°C(站位:39.16°N,120.27°E),最低溫度16.2°C(站位:39.08°N,120.94°E);渤海各海域底層溫度變化較大,10—20°C均有分布,萊州灣底層溫度要顯著高于其他海域(F=4.438,P=0.008)。渤海各海域表、底層鹽度變化較大,最大差值可達3.2;萊州灣表、底層鹽度均要顯著低于其他海域(表層:F=13.470,P<0.001;底層:F=48.842,P<0.001),黃河入海口附近海域形成鹽度低值區。
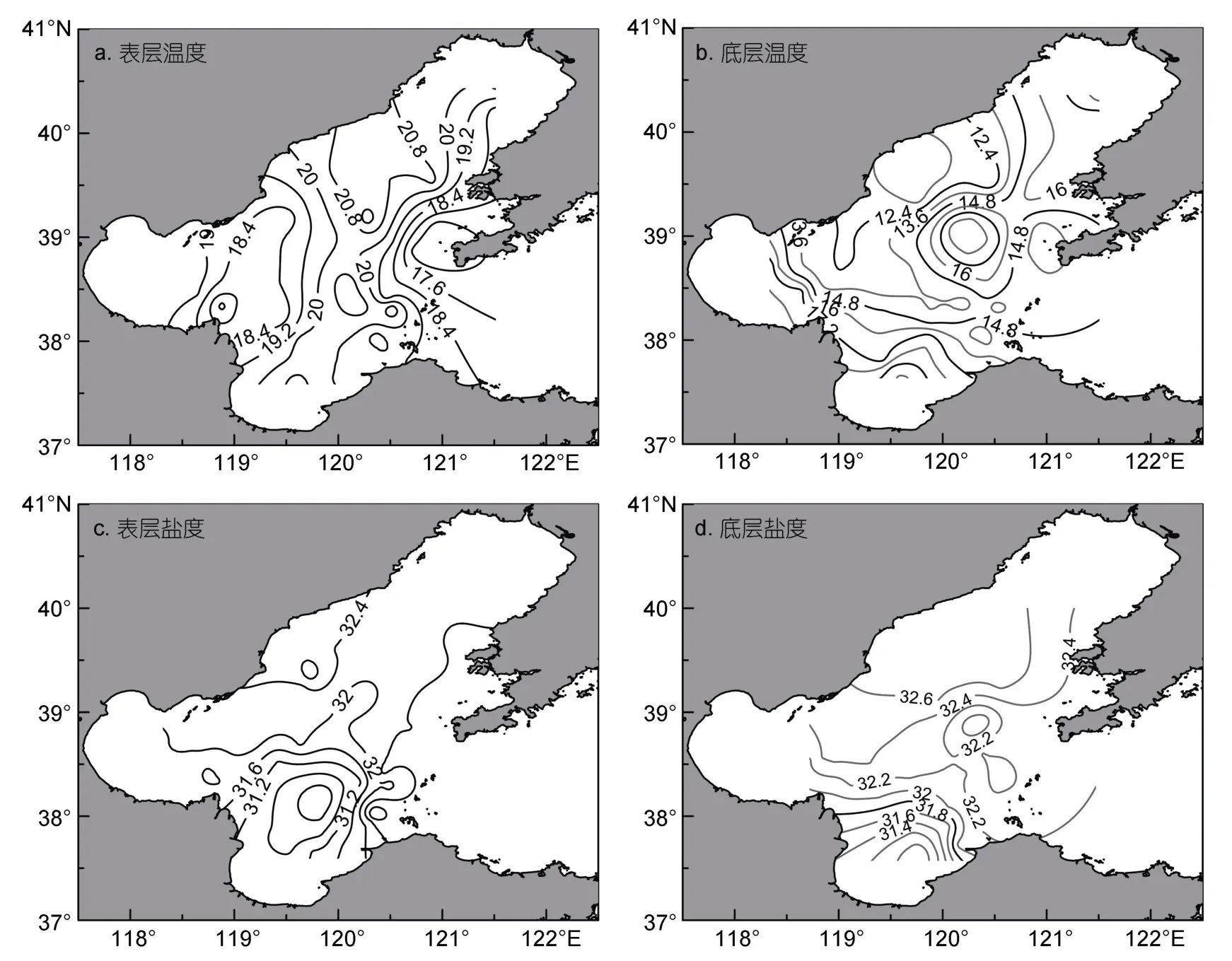
圖2 調查海域表、底層溫度與鹽度分布Fig.2 Distribution of temperatures(°C)and salinity at the sea surface and bottom
2.2 水母種類與密度
本航次共發現四種大型水母:海月水母(Aurelia coerulea);沙海蜇(Nemopilema nomurai)、海蜇(Rhopilema esculentum);多管水母(Aequoreasp.)。
本次調查共有9個站位出現海月水母,19個站位出現沙海蜇,分別占調查站位的19%、40%(圖3a,3b)。海月水母密度最大為221.21ind./(net·h),出現在站位(38.34°N,118.86°E),其次為 38ind./(net·h),出現在站位(38.30°N,119.06°E),其余海月水母出現站位密度在1—5ind./(net·h);渤海海域6月份海月水母密度地理分布特征為:高密度區域集中于渤海灣外部海域,萊州灣、遼東灣海域海月水母密度較小,遼東灣北部、渤海灣內部、渤海中東部海域未見海月水母出現(圖3a)。Pearson相關性分析結果表明,溫度、鹽度對海月水母密度影響不顯著(P>0.05)。
沙海蜇密度最大為75.6ind./(net·h),出現在站位(39.73°N,121.15°E);遼東灣海域沙海蜇出現站位密度均值為(35.32±21.64)ind./(net·h);渤海灣外側、渤海中部海域沙海蜇出現站位密度均值為(3.68±3.34)ind./(net·h);在渤海海域沙海蜇高密度分布區域為萊州灣、渤海灣外側海域以及遼東灣海域(圖3b)。Pearson相關性分析結果表明,除表層鹽度對沙海蜇密度具有顯著影響外(P<0.05),溫度與底層鹽度均對沙海蜇密度影響不顯著(P>0.05)。
海蜇在測區僅在一個站位出現(39.73°N,120.24°E),且密度較低[1—2ind./(net·h)];多管水母同樣只有一個站位出現,密度在[1—2ind./(net·h)],位于站位(39.51°N,120.95°E)(圖c,3d)。
2.3 傘徑大小分布
2018年6月渤海海域海月水母傘徑均值最大為8.3cm,出現在站位(38.34°N,118.86°E);均值最小為1.8cm,出現在站位(38.34°N,119.02°E)(圖a);遼東灣南部海月水母平均傘徑(7.00±1.00)cm,渤海中部均值為(6.03±2.99)cm,渤海灣均值為(4.95±2.57)cm,萊州灣(5.33±0.58)cm(圖5a),非參Kruskal-Wallis檢驗結果顯示,渤海各海區海月水母傘徑大小無顯著差異(H=2.453,df=3,P=0.484>0.05)。總體來講,6月份渤海海域海月水母傘徑較小,大部分個體在10cm以下,各海區傘徑大小無顯著差異。
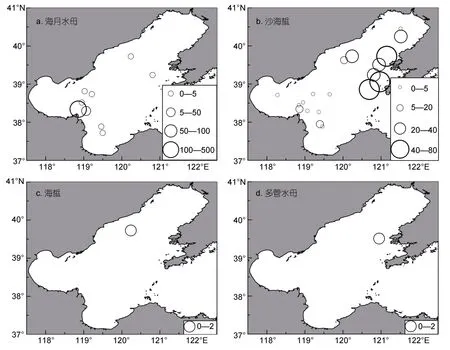
圖3 2018年6月渤海海域大型水母漁獲密度[ind./(net·h)]分布圖Fig.3 Distributions of densities[ind./(net·h)]of Aurelia coerulea(a),Nemopilema nomurai(b),Rhopilema es culentum(c)and Aequorea sp.(d)in the Bohai Sea in June 2018
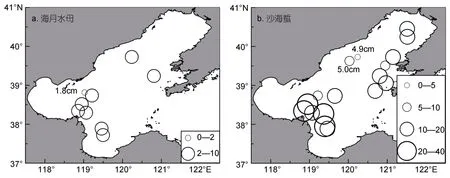
圖4 2018年6月渤海海域各站位海月水母(a)與沙海蜇(b)平均傘徑大小(cm)分布圖Fig.4 Distributions of mean diameters(cm)of Aurelia coerulea(a),Nemopilema nomurai(b)in the Bohai Sea in June 2018
2018年6月份渤海海域沙海蜇傘徑均值最大為39cm,出現在站位(38.34°N,118.86°E);均值最小為4.9—5cm,出現在遼東灣西側、秦皇島外側海域(圖4b);遼東灣沙海蜇平均傘徑(12.15±6.52)cm,渤海中部均值為(17.45±9.20)cm,渤海灣均值為(33.86±7.40)cm,萊州灣(27.67±16.50)cm(圖b),非參Kruskal-Wallis檢驗結果顯示渤海灣、萊州灣外側交界海域沙海蜇傘徑要顯著大于遼東灣海域(H=27.5,df=3,P<0.001)。總體來講,6月份渤海海域沙海蜇傘徑分布極不均勻,具體呈現為渤海灣、萊州灣外側交界海域沙海蜇傘徑要顯著大于遼東灣海域,且在秦皇島外側海域(39.63°—39.73°N,120.03°—120.23°E)發現沙海蜇個體傘徑在1—3cm之間。
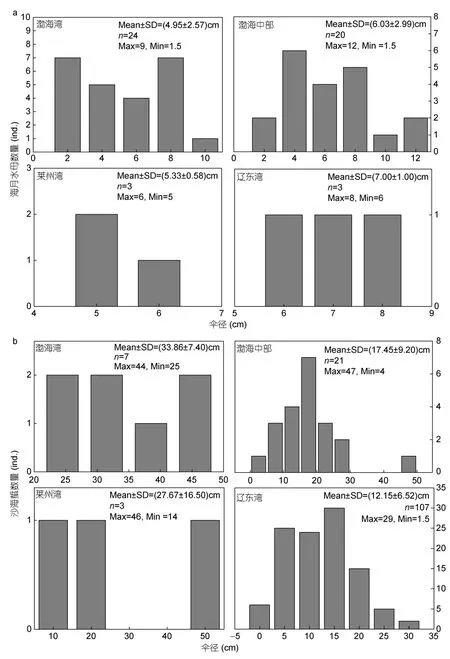
圖5 海月水母(a)與沙海蜇(b)在渤海各海域(渤海灣、萊州灣、遼東灣、渤海中部)傘徑頻度分布圖Fig.5 Frequencies of Aurelia coerulea(a),Nemopilema nomurai(b)in the central part of Bohai Sea,Laizhou Bay,Bohai Bay and Liaodong Bay,in June 2018,respectively
3 討論
3.1 渤海大型水母分布及源地探討
3.1.1 海月水母 6月份渤海海月水母在三灣海域均有分布,渤海灣東南部為海月水母密度高值區,可達38—221.21ind./(net·h),其他站位密度較低,多在 5ind./(net·h)以下(圖a)。雖然 Pearson相關性分析顯示溫度均對海月水母密度不具有顯著影響,但渤海灣東南部海域底層溫度要顯著高于其他海域(圖2b),Wang等(2015b)指出海月水母水螅體在8—17°C均能產生橫裂生殖,但13°C是其最適橫裂溫度;董婧等(2013)指出海月水母水螅體在15—20°C橫裂生殖產生的碟狀體比例最高。調查時發現,遼東灣海域多數站位底層溫度在13°C以下,而萊州灣、渤海灣交界海域底層溫度均在15°C以上,這可能導致此處海月水母水螅體橫裂發生時間要早于其他源地,因此呈現出密度高值區。此外,此部分海域鹽度要顯著低于其他海域,最大差值可達3以上(圖2c,2d)。但眾多研究表明,海月水母是一種廣鹽性生物,具有很強鹽度適應性(董婧等,2013),因此,在本研究中Pearson相關性分析顯示鹽度對海月水母分布不具有顯著影響。
各海域海月水母傘徑均值較小,多為10cm以下的水母幼體(圖4a),但海月水母在野外海域傘徑最大可達30cm以上,這表明6月上旬海月水母在渤海處于發生初期階段。渤海灣東南部海域海月水母傘徑多集中于 4.95—6.03cm,最大可達12cm,最小值為1.5cm(圖4a,5a),據此推測,此處出現的海月水母應為土著種,萊州灣與渤海灣交界海域可能是渤海海月水母種群發源地之一。渤海灣內部測區未見海月水母,東側河北沿岸海域有兩個站位出現,密度為2—4.8ind./(net·h)(圖3a),值得注意的是所有出現的海月水母傘徑僅有1.5—3cm(圖4a,5a),作者推測,附近海域同樣可能為渤海海月水母種群源頭之一。
本次調查發現,遼東灣海月水母僅出現于灣南,在秦皇島外海與大連外海兩個站位有所顯現,密度在1.5—2ind./(net·h),且均為海月水母幼體(傘徑均值為7cm±1cm;圖4a,5a),在灣北測區未見海月水母(圖3a),這可能由于本研究未在灣北近海設置調查站位所致。事實上,這與遼寧水科院長期對遼東灣海月水母觀測結果類似,2004—2011年調查中,同樣發現海月水母主要分布于遼東灣灣南,灣北海域較少出現(王彬等,2012;董婧等,2013),例如:2010年海月水母在灣南密度可達533ind./(net·h),而灣北僅有1—15ind./(net·h)(王 彬 等,2012)。王 彬 等(2012)于2009—2011年6月下旬調查時發現灣北海月水母有增加的趨勢,這可能由于灣北溫度要低于灣南,因此海月水母在灣北發生時間要晚于灣南,推遲到6月下旬。作者認為遼東灣南部海月水母于6月上旬開始出現,這與前人調查結果一致;由于此次調查對遼東灣站位調查時間集中于6月中上旬,此時海月水母在遼東灣尚未大規模出現,因此所測密度要顯著低于前人調查結果。
萊州灣內只有兩個站位出現海月水母,密度在1—4ind./(net·h)(圖a),傘 徑 均 值 為(5.33±0.58)cm(圖5a),但據左濤等(2016b)的調查結果,6月份萊州灣未發現有海月水母出現,直到7月份才有一個站位出現海月水母,而到9、10月份海月水母會在個別站位集中出現,生物量最高可達45454kg/km2,這表明不同年份該海域海月水母的發生時間、旺發時間會有所不同。
一方面海月水母水螅體具有很強的環境適應性,其最適鹽度范圍可在17.5—37(董婧等,2013),即使在本研究中發現各海域鹽度有所差異(30—34),但這并不影響水螅體生存與生殖;另一方面,各地人工設施的建造,如防波堤、海港、碼頭等,為水螅體的附著提供了適宜的棲息地(Uyeet al,2004;Holstet al,2007),因此,海月水母在渤海沿岸可能存在多個源頭,諸如:萊州灣與渤海灣交界近岸海域、河北近岸、遼東灣大連近岸以及北部近岸,此處所提出的海月水母源頭是基于本研究所給出的推測,當然在渤海其他近岸海域可能同樣存在海月水母水螅體棲息地,這需要更加深入的調查才能清楚的了解渤海海月水母的發生地。在6月上旬溫度適宜時,渤海海月水母水螅體逐漸橫裂生殖釋放碟狀體。
3.1.2 沙海蜇 6月份沙海蜇在渤海分布范圍較廣,集中出現于兩部分海域:遼東灣、萊州灣和渤海灣交界外側海域,且遼東灣沙海蜇密度要顯著高于萊州灣與渤海灣外側(圖3b),但前者沙海蜇傘徑要顯著大于后者(圖5b)。前人研究指出沙海蜇水螅體更適合在低鹽海水中生長(董婧等,2013),Feng等(2016)同樣指出河口附近低鹽海區可能成為沙海蜇發生源頭,萊州灣和渤海灣交界外側海域由于黃河淡水注入,形成鹽度低值區;但本次調查并未在此近海發現沙海蜇幼體的存在,因此作者推測,萊州灣和渤海灣交界海域所出現的沙海蜇并非土著種。Pearson相關性分析表明,雖然表層鹽度對沙海蜇密度具有顯著影響,但溫度與底層鹽度均對其無顯著影響,作者認為溫鹽條件并不能對沙海蜇的分布特征作出很好的解釋。但水母自主游動能力很弱,其分布很大程度受到風流、海流的影響。6月調查時發現,渤海灣傘徑均值為(33.86±7.40)cm,傘徑最小也有25cm,萊州灣采集到的沙海蜇傘徑均值為(27.67±16.50)cm,渤海中部出現的沙海蜇傘徑均值也有(17.45±9.2)cm(圖5b),沙海蜇傘徑要顯著大于遼東灣,渤海灣、萊州灣出現的沙海蜇傘徑最小分別為25、14cm,最大可達46cm(圖5b)。春、夏季渤海中部存在較為明顯的順時針大流環和逆時針小流環,遼東灣內部存在明顯的逆時針流環(Weiet al,2001);渤海灣灣口北部存在一支形態清晰的沿岸南下入流,其在灣內僅到達南堡附近便轉向東南,從灣口中部流出,夏季在老黃河口外側流向發生轉變,向北匯入渤海中部順時針大流環(Zhouet al,2017)。作者推測在此環流作用下,遼東灣或者其他海域的沙海蜇進入到渤海灣、萊州灣灣口附近海域,由于此部分沙海蜇出現較早,經過長時間漂移傘徑自然要大于遼東灣,但整體密度并不高。
本次調查發現,沙海蜇在大連外海形成密度高值區,密度在39—80ind./(net·h)(圖3b),其傘徑分布極不均勻,從1.5cm到29cm均有出現;在遼東灣北部有兩個站位出現沙海蜇,密度范圍在3—37ind./(net·h),傘徑均值為10—20cm,最小個體為8cm(圖4b)。王彬等(2012)于2009—2011年對遼東灣北部海域調查時發現,5月底未見沙海蜇,海區中能采集到沙海蜇幼體最早的時間為6月上旬,且在灣北近海雙臺子河口有大量沙海蜇幼體聚集,形成數量高峰(王彬等,2013),但本次調查發現,6月上旬灣南沙海蜇個體最大可達29cm,這表明灣南沙海蜇發生時間要明顯早于灣北,這可能是由于溫度差異導致水螅體橫裂時間的不同。可以看出,雖然沙海蜇在遼東灣分布范圍較廣,但同時具有分布不均勻的特點。另外,我們在秦皇島東側海域同樣采集到沙海蜇,其密度為7.5—26ind./(net·h)(圖b),該海域沙海蜇傘徑均值不足5cm(圖4b),最小個體只有1.5cm。作者推測,秦皇島東側海域出現的沙海蜇可能與遼東灣北部為不同的兩個群體,同樣在6月上旬逐漸開始出現。
鄭向榮等(2014a,b)2012年8—9月對河北沿岸調查時發現,該海區沙海蜇生物量要顯著高于其他水母,且分布較廣,從秦皇島浴場到渤海灣大部均有分布,灤河口附近生物量更是高達15—75t/km2,而本次調查在渤海灣內只有一個站位采集到沙海蜇,且密度只有0.67ind./(net·h),一方面可能由于本次調查時間處于6月上旬,沙海蜇處于發生初期,因此尚未形成如此高密度,另一方面沙海蜇分布可能具有較大的年際差異性。沙海蜇在萊州灣內部并未采集到,只有灣外站位出現,這與左濤等(2016b)在2011年調查結果類似。
通過本研究,作者認為沙海蜇在渤海海域發源地主要集中于遼東灣,在灣南大連近岸海、秦皇島近岸,以及灣北都有分布,6月初逐漸顯現。渤海中部、萊州灣、渤海灣所出現的沙海蜇并非本地種,很可能是在海流作用下由遼東灣或其他源頭漂流至此。
3.1.3 海蜇與多管水母 海蜇屬暖水性大型水母,幼體多棲息于河口咸淡水交匯海域(李培軍等,1988;Donget al,2009)。本次調查中,所采集到的海蜇出現在站位(39.73°N,120.24°E)(圖c),位于秦皇島外側海域,傘徑為5.5cm。海蜇作為我國具有很高經濟價值的漁業資源,總體來講,在渤海的自然資源量下降明顯,渤海灣、萊州灣海蜇資源難以形成汛期(紀靈等,1994),只是零星出現,長期對海蜇資源進行增殖放流,使得遼東灣成為尚能形成海蜇集中漁汛的代表性漁場(姜連新等,2007)。多管水母在本次調查中只采集到一只,出現在大連瓦房店外側海域(圖3d),傘徑為7cm,渤海其他海域未見。
3.2 渤海與黃東海海月水母、沙海蜇對比
海月水母與沙海蜇在我國近海分布較廣,在統計歷史調查數據的基礎上,對兩種水母在渤海與黃、東海的生長進行了對比。黃海海月水母的報道多見于膠州灣,據萬艾勇等(2012)于2009年調查時發現海月水母碟狀體在膠州灣4月份開始出現,到6月份達到高峰,6月上旬傘徑為(8.74±1.72)cm,下旬可達(15.21±4.29)cm;2010年6月中旬海月水母傘徑可達(11.5±2.5)cm(表1);Wang 等(2015a)在 2011年對膠州灣調查時報道,海月水母幼體在5月下旬傘徑為(0.97± 0.2)cm,7月初可達(16.2±1.8)cm。而王彬等(2012)在2009—2011年對遼東灣北部調查時發現,5月下旬未見海月水母幼體,到6月下旬傘徑只有5—10cm;同樣,本次調查發現6月份渤海所出現的海月水母傘徑多數在10cm以下,最大個體也不過12cm,渤海灣東南部海月水母傘徑只有1.5cm。因此,膠州灣海水母發生時間要早于渤海,且在同一月份傘徑要高于渤海。
6月份黃海與東海北部沙海蜇發生時間同樣要早于渤海,且同期傘徑明顯要大(表1)。據程家驊等(2004)報道,2003年6月中旬東海北部沙海蜇傘徑可達 40—50cm,2004年可達50—70cm;據Zhang等(2012)在2007年調查時報道,5月份在南黃海、東海西北部均有沙海蜇分布,到6月份其傘徑可達30cm以上;李建生等(2009)報道2008年6月中旬在東海北部沙海蜇傘徑范圍在10—50cm。而在遼東灣最早的沙蜇發現記錄是在6月上旬,此時沙海蜇傘徑在2—10cm,而到6月中旬傘徑也只有6—12cm(王彬等,2010)。本次調查發現,在渤海海域沙海蜇傘徑大小分布具有不均勻性,遼東灣海域沙海蜇平均傘徑只有(12.15±6.25)cm,在秦皇島外海位采集到的沙海蜇傘徑均值甚至不足5cm,而在萊州灣與渤海灣交界海域出現的沙海蜇其傘徑均值在17—34cm,最大可達47cm(圖5b),但從總體來看,黃海、東海北部沙海蜇種群出現時間要先于渤海沙海蜇種群。
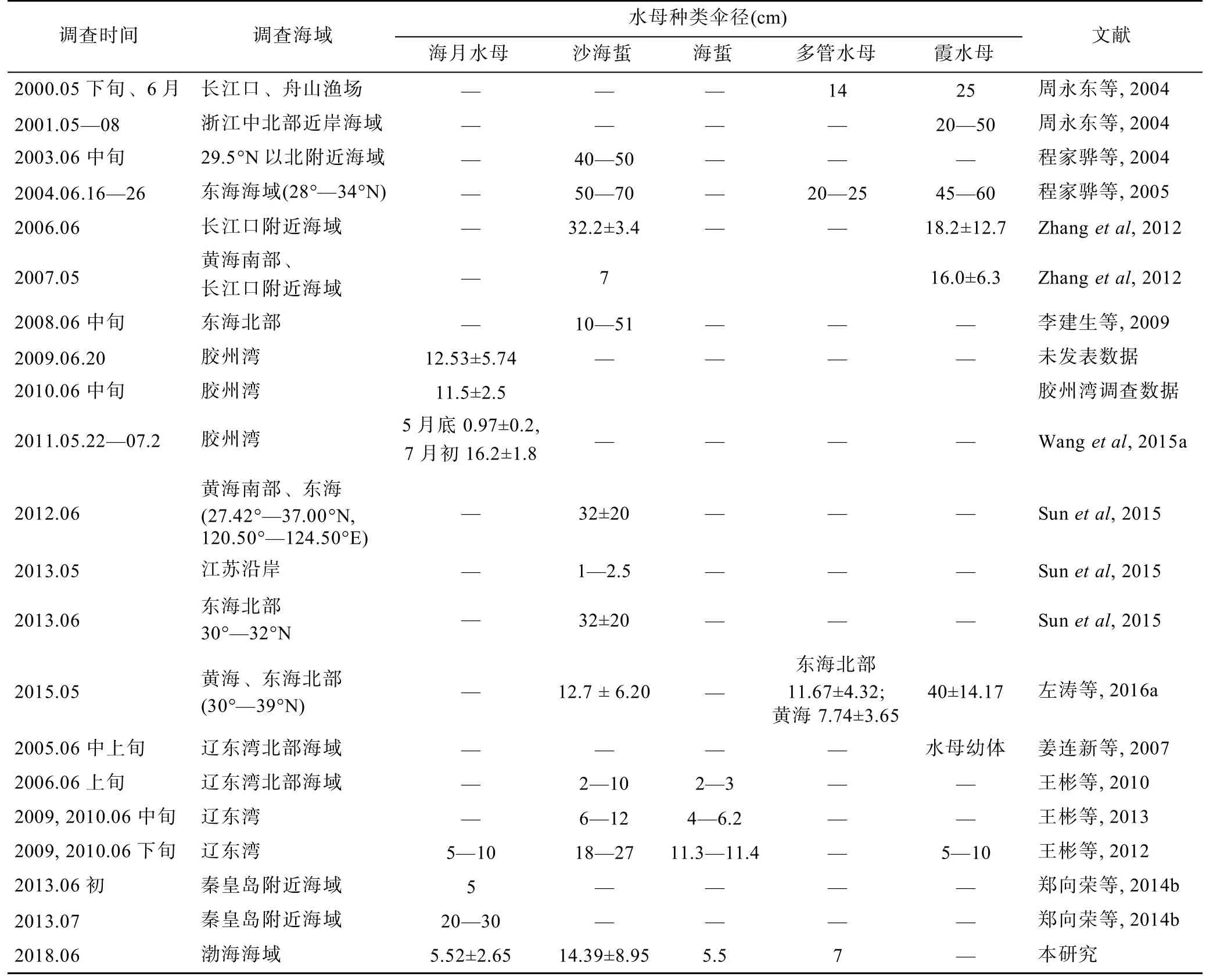
表1 6月份前后東海北部、黃海以及渤海各海域海月水母、沙海蜇、海蜇、多管水母、霞水母傘徑大小歷史記錄Tab.1 previous studies of diameters of Aurelia sp.,Nemopilema nomurai,Rhopilemaes culentum,Aequorea spp.,and Cyanea spp.,in the Yellow Sea,northern East China Sea,and Bohai Sea in June,2018
作者認為海月水母、沙海蜇發生時間與傘徑最大值的出現期,隨著緯度增加均有推遲的現象。溫度對水螅體橫裂、水母體生長起到至關重要的作用,隨緯度升高溫度降低,這可能是導致水母發生時間推遲的主要原因。另外,浮游動物的組成在黃、東海與渤海有很大差異(代魯平等,2016),這為水母提供了不同的餌料環境,餌料差異不僅能夠影響水螅體階段的分裂生殖(Wanget al,2015b),對于水母體的生長發育更會產生很大影響,這也可能是導致黃、東海與渤海水母種群差異的原因之一。
4 結論
2018年6月海月水母、沙海蜇、海蜇、多管水母在渤海均有分布,但以海月水母、沙海蜇為主。總體來講,6月份渤海海月水母分布主要集中于渤海灣東南側海域,遼東灣海月水母主要出現于灣南,各海域海月水母傘徑均較小,多為水母幼體;沙海蜇分布廣泛,但其密度分布與傘徑分布具有不均勻的特點,遼東灣為沙海蜇密度高值區,但其傘徑較小,渤海灣與萊州灣外側海域沙海蜇密度雖小,但傘徑要顯著大于遼東灣;海月水母、沙海蜇在渤海發生時間要晚于黃、東海。
致謝感謝中國水產科學研究院黃海水產研究所承擔的青島海洋科學與技術試點國家實驗室共享搭載航次和環境數據,及相關調查人員和“中漁科102”號調查船全體船員的辛勤外業工作。
