三亞蜈支洲島珊瑚礁-沙質底復合區棘皮動物群落結構*
2020-02-06 06:00:42黃端杰李秀保薛英樓吳沛霖
海洋與湖沼 2020年1期
黃端杰 許 強 李秀保 薛英樓 吳沛霖 高 菲
(南海海洋資源利用國家重點實驗室 海南大學海洋學院 海口 570228)
我國南海的珊瑚礁生態系統是生物資源寶庫,同時也蘊藏著豐富的底棲棘皮動物資源。大型底棲動物與珊瑚礁之間存在密切的相互依存關系,如以棘皮動物為代表的底棲動物群落能夠反映珊瑚礁生態系統的健康狀況,其生物多樣性對珊瑚礁的穩定和功能具有重要的意義(朱文濤等,2018)。研究表明,珊瑚礁生態系統的健康程度與棲息在礁內生物的多樣性呈正相關關系。李元超等(2015)通過調查三亞后海海域珊瑚礁健康狀況后分析得出,珊瑚礁區高密度的植食性動物是該區域珊瑚礁生態系統保持健康的主要原因之一,珊瑚礁區海膽通過攝食珊瑚礁上附著的藻類,確保珊瑚能夠正常生長。因此,礁區生物多樣性是反映珊瑚礁生態系統變化的重要指征之一(Vázquez-Domínguez,2003;Langmeadet al,2004)。
棘皮動物作為大型底棲動物的重要類群,在維護和調節珊瑚礁底棲生態系統中起著不可或缺的作用(廖玉麟等,2011)。以珊瑚礁生態系統里的海參綱和海膽綱為例,海參綱多以海底沉積物為食,凈化海底同時促進營養鹽循環,珊瑚礁內海參(玉足海參和花刺參)通過腸道消化含有碳酸鹽的沙子和碎石,在攝入食物同時溶解碳酸鈣增加珊瑚礁堿度,防止海水酸化,有利于珊瑚礁的生長(Schneideret al,2011)。海膽在珊瑚礁生態系統中扮演著極其關鍵的角色,海膽以藻類為食,可以抑制大型藻類的過度生長,避免其爭奪珊瑚正常的生長空間(Tuyaet al,2005)。因此,珊瑚礁內棘皮動物群落生物多樣性調查和生態學分析可以作為指示礁區健康監控手段之一。
海南三亞海域擁有中國大陸發育最好的岸礁型珊瑚礁區域,與此同時,也頻繁受到人為活動影響,因此是研究人類活動-珊瑚礁生態系統健康相關關系的絕佳區域。位于海棠灣蜈支洲島的珊瑚礁區在三亞蜈支洲島旅游區(5A級)的綜合管理下,通過采取嚴格的資源環境管護措施,使得島礁周邊的珊瑚生態系統得以有效保護,因此成為海南省珊瑚礁分布的代表區域之一。在2011年對海南島東、南部珊瑚礁生態健康初步調查中,發現蜈支洲島活珊瑚種類數量僅次于鹿回頭,且蓋度分布最高(吳瑞等,2013)。近幾年,國內對三亞海域珊瑚群落現狀的研究較多,但反映珊瑚礁健康情況的底棲生物群落研究較為匱乏。本文擬選擇蜈支洲島珊瑚礁代表區域,調查研究棘皮動物群落多樣性現狀及其季節變化,評估其群落健康狀況,進一步為該區域珊瑚礁生態系統健康狀況評價提供參考數據。
1 材料與方法
1.1 研究區域概況
蜈支洲島位于三亞東南側海棠灣內,是典型的近岸熱帶島礁(圖1)。島東西兩端相距1500m,南北兩端相距1100m,全島總面積為1.48km2。海岸至水深4—5m處的底質類型主要由珊瑚生長發育形成的巖礁、5—10m主要為活珊瑚生長地帶,海底沉積物類型有基巖、珊瑚礁→粗砂→細砂→中砂混合→中細砂→細砂→粉砂,逐漸變細。島北岸西部沙灘由主要泥沙落淤形成,西北側還形成了沙咀(顏慧慧,2017)。

圖1 蜈支洲島海域棘皮動物調查范圍圖Fig.1 The investigation area of echinoderms in Wuzhizhou Island
在前期的底棲生境調查中發現,蜈支洲島東南側近岸海域為活珊瑚礁-巖礁底質復合區,海底地形陡峭,10m以淺水域狹窄;島西南側以沙質底為主,珊瑚覆蓋度較低;而島北側近岸海域為活珊瑚礁塊-沙質底復合分布區,海底平緩,在該處分布的棘皮動物棲息密度(特別是海參類)遠較其它區域高。同時,島北側靠近游艇碼頭,為旅游潛水地點,人為活動頻繁,因此本文選擇島北側活珊瑚礁塊-沙質底復合分布區作為重點研究區域。
1.2 調查方法與內容
1.2.1 調查時間 2018—2019年共進行了四個季度月調查,分別為2018年8月、2018年11月、2019年1月和2019年4月。
1.2.2 水質因子調查 利用多參數水質測試儀(美國,YSI-650)對調查區域表層水體(水下1m)進行基本水質理化因子監測,指標包括溫度、鹽度、pH和溶解氧。
1.2.3 棘皮動物群落調查方法 棘皮動物群落調查采用隨機樣帶結合視頻定量調查法。具體操作如下:
(1)隨機樣帶設置
在研究區域(18°19.000′—18°19.067′N,109°45.890′—109°46.009′E,圖)內利用手持式測深儀測量水深,分別在淺水區(3—5m)和深水區(8—10m)用繩子隨機布置3條100m樣帶,樣帶兩側各1m為視頻拍攝定量范圍,最終構成100m×2m(200m2)的樣方,作為資源密度的統計基準。
(2)視頻拍攝方法
潛水員手持水下高清攝像機(GoPro 5,美國)沿樣帶拍攝,分辨率為2.7kp,幀速為60幀/秒,拍攝時潛水員的平均游泳速度為0.17m/s。拍攝視頻的截圖清晰,能夠滿足水下棘皮動物種類鑒定的要求。
(3)種類鑒定
由于采用視頻拍攝數據和單一種類水下照片進行種類鑒定,本研究中僅將形態特征顯著的常見棘皮動物種類納入統計范圍。代表性種類特征圖片和特征描述參考《中國動物圖譜:棘皮動物》(張鳳瀛,1964)和《中國動物志:棘皮動物門 海參綱》(廖玉麟,1997),分類采用Pawson et Fell(1965)的分類系統。
1.3 數據處理
利用Excel將各季節和不同水深的棘皮動物數量換算為單位面積的棲息密度(ind./10m2)。
生物多樣性指數(物種豐富度指數、多樣性指數、均勻度指數)和優勢度的計算方法分別采用牛翠娟等(2015)和董棟等(2015)。
a.優勢度(Y)

式中,fi為第i個種在各樣方中出現頻率,ni為群落中第i個種在空間中的個體數量,N為群落中所有種的個體數總和。
b.棲息密度ρ

式中,ρ為棘皮動物棲息密度,N為所有樣方中棘皮動物的數量,S為所有樣方面積,單位為ind./10m2。
1.4 統計分析
采用數據分析軟件SPSS(PASW Statistics 18,美國)進行數據分析,分別對4個季節的水質因子,淺水區和深水區棘皮動物棲息密度進行單因素方差分析(ONE-WAYANOVA),檢驗不同季節水環境指標,淺水區和深水區棲息密度的差異,顯著性水平設置為0.05。
2 結果
2.1 蜈支洲島北海域水質特征
在2018—2019年4次監測中,冬季水溫最低,為23.72°C,夏季水溫最高,為27.39°C。全年鹽度相差不大,變化范圍為31.54—33.77。溶解氧為5.16—5.49mg/L,春季最高,冬季最低。pH為7.61—8.40,最低為春季,最高為冬季。

表1 調查期間基本水文特征Tab.1 Basic hydrological characteristics during the investigation period
2.2 棘皮動物種類組成
在為期一年的調查中,共發現棘皮動物15種,隸屬于4綱10科:海參綱有6種,占總數的40%;海星綱有5種,占34%;海膽綱2種,占13%;海百合綱2種,占13%。在不同季節調查中,春季發現棘皮動物種類最多,有13種。周年調查中均以海參綱占優勢(表2)。
不同深度調查中,夏季淺水區分布的棘皮動物種類多于深水區,而秋季和冬季深水區高于淺水區。兩個深度分布的種類大致相同,海參綱均占優勢。周年調查中,兩個水深共有的種類是紅腹海參、黑海參、呂宋棘海星、刺冠海膽和許氏大羽花。對比之下,綠刺參未在冬季淺水區發現,本氏海齒花未在秋季淺水區發現,這2種在其他季節和深度均有分布。玉足海參未在秋冬季節淺水區發現,黃疣海參未在春秋兩季深水區發現,這2種在其他季節和深度均有分布。面包海星僅在春冬兩季深水區發現,多篩指海星僅發現于夏季淺水區和秋季深水區。斑錨參僅發現在春季淺水區,長棘海星僅發現于夏季淺水區(表3)。
2.3 不同綱棘皮動物分布密度的季節與深度變化
棘皮動物棲息密度季節變動情況整體呈現出春夏高,秋冬低的特點。不同綱的棘皮動物棲息密度在春夏兩個季節呈現出升高趨勢,到了秋冬季節呈現下降趨勢。全年棘皮動物平均棲息密度變化較大,以冬季最低,僅為3.9ind./10m2;夏季最高,為10ind./10m2;春季稍低于夏季,為9.2ind./10m2;秋季為7.7ind./10m2(表4)。
總體上,深水區分布的棘皮動物棲息密度高于淺水區,但差異并不顯著(F=5.08,P>0.05,圖2)。海參綱、海星綱和海膽綱的優勢種在深水區棲息密度遠高于淺水區,例如海參綱紅腹海參和黑海參數量遠高于其他海參,2種數量之和占全部海參總數的91.25%;海星綱以呂宋棘海星為主,數量占到全部海星的 61.29%(表4)。
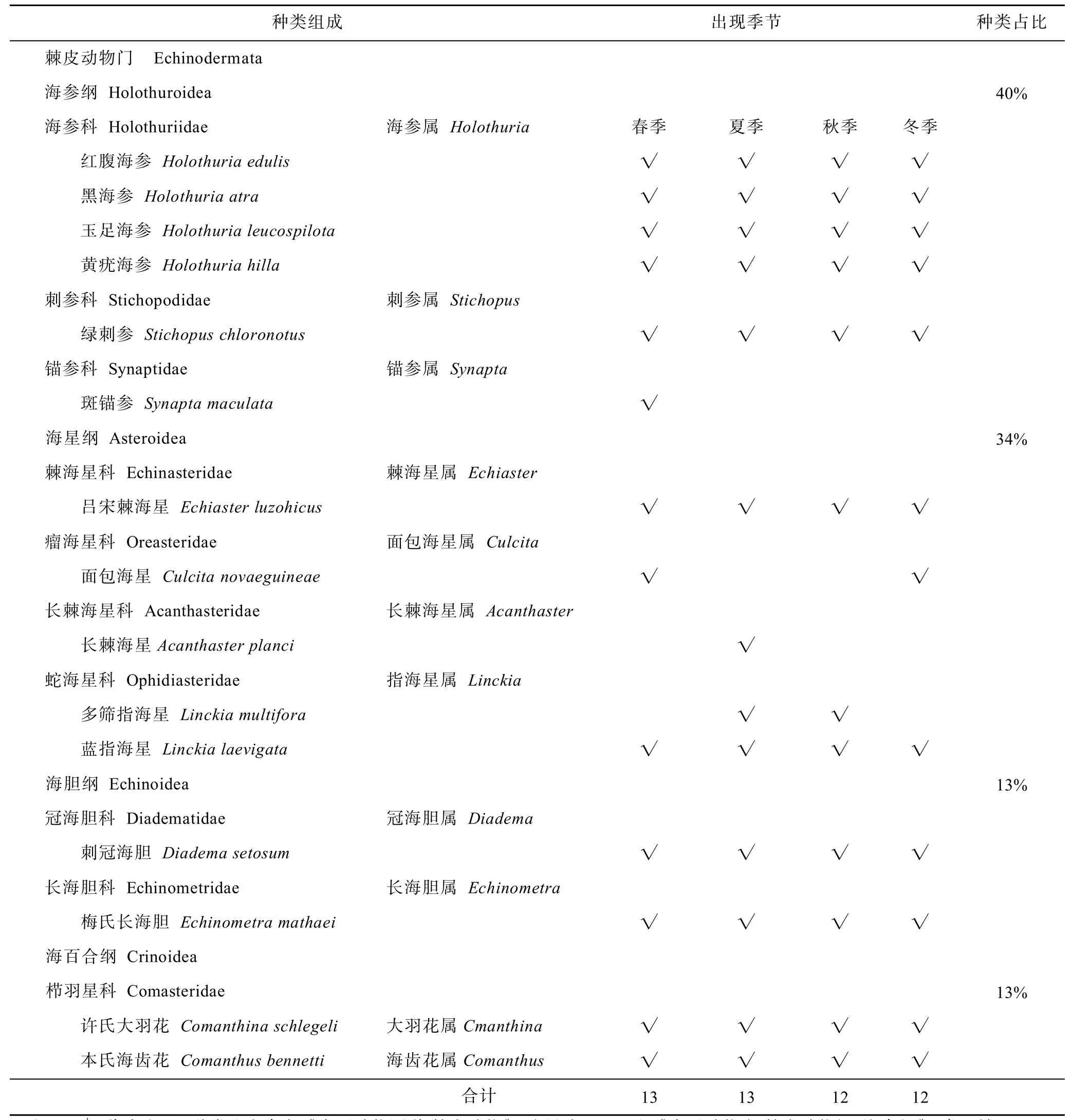
表2 不同季節棘皮動物種類組成情況Tab.2 Species composition of echinoderms in different seasons
2.4 棘皮動物優勢種的季節變化
按優勢度≥0.02來劃分優勢種類,共發現8種優勢種類,分別是黑海參、紅腹海參、黃疣海參、綠刺參、呂宋棘海星、許氏大羽花、本氏海齒花、刺冠海膽,優勢類群以海參綱占優勢。第一優勢種為紅腹海參,優勢度在夏季達到0.52。黑海參、刺冠海膽和許氏大羽花優勢度較紅腹海參小,但也達到了較高水平,其他種類優勢度一般。
各季節優勢種數量為7種,其中屬于全年優勢種的有黑海參、紅腹海參、呂宋棘海星、許氏大羽花和刺冠海膽。屬于三季優勢種為本氏海齒花和綠刺參。黃疣海參僅在夏季為優勢種。
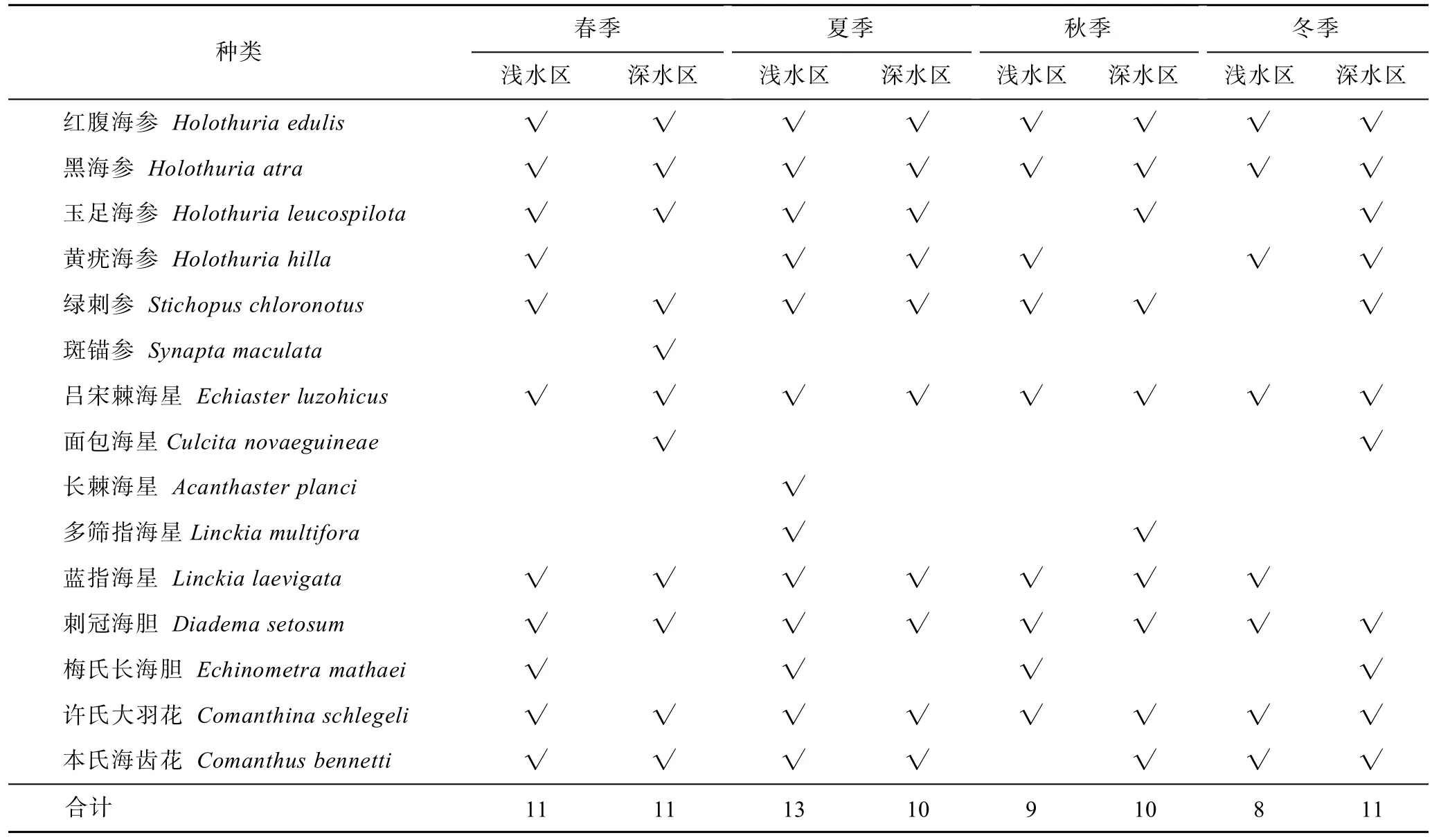
表3 不同季節淺水區和深水區棘皮動物種類組成Tab.3 Species composition of echinoderms in shallow water and deep water in different seasons

表4 不同季節淺水區和深水區棘皮動物棲息密度(單位:ind./10m2)Tab.4 Density of echinoderms in shallow water and deep water in different seasons

圖2 不同季節淺水區和深水區棘皮動物棲息密度Fig.2 Density of echinoderms in shallow water and deep water in different seasons

表5 不同種類棘皮動物優勢度季節變化Tab.5 Dominant species of echinoderms in different seasons
2.5 棘皮動物生物多樣性指數的季節變化
均勻度指數全年變動較大,呈現交替變化。多樣性指數全年變化較大,春季和秋季最低,為2.4。物種豐富度指數在夏季有一定下降,其他季節變化不大。
對于各個綱,海參綱均勻度指數全年變動較大,呈現交替變化。多樣性指數全年變化較大,秋季最低,為1.0。物種豐富度指數在夏季有一定下降,其他季節變化不大。海星綱種豐富度指數全年變化很小,均勻度指數在夏季有一定下降,多樣性指數全年變化最大,夏季最低,為0.56。由于海百合綱和海膽綱只發現2種,得出的生物多樣性指數無討論意義,故不計算(表6)。
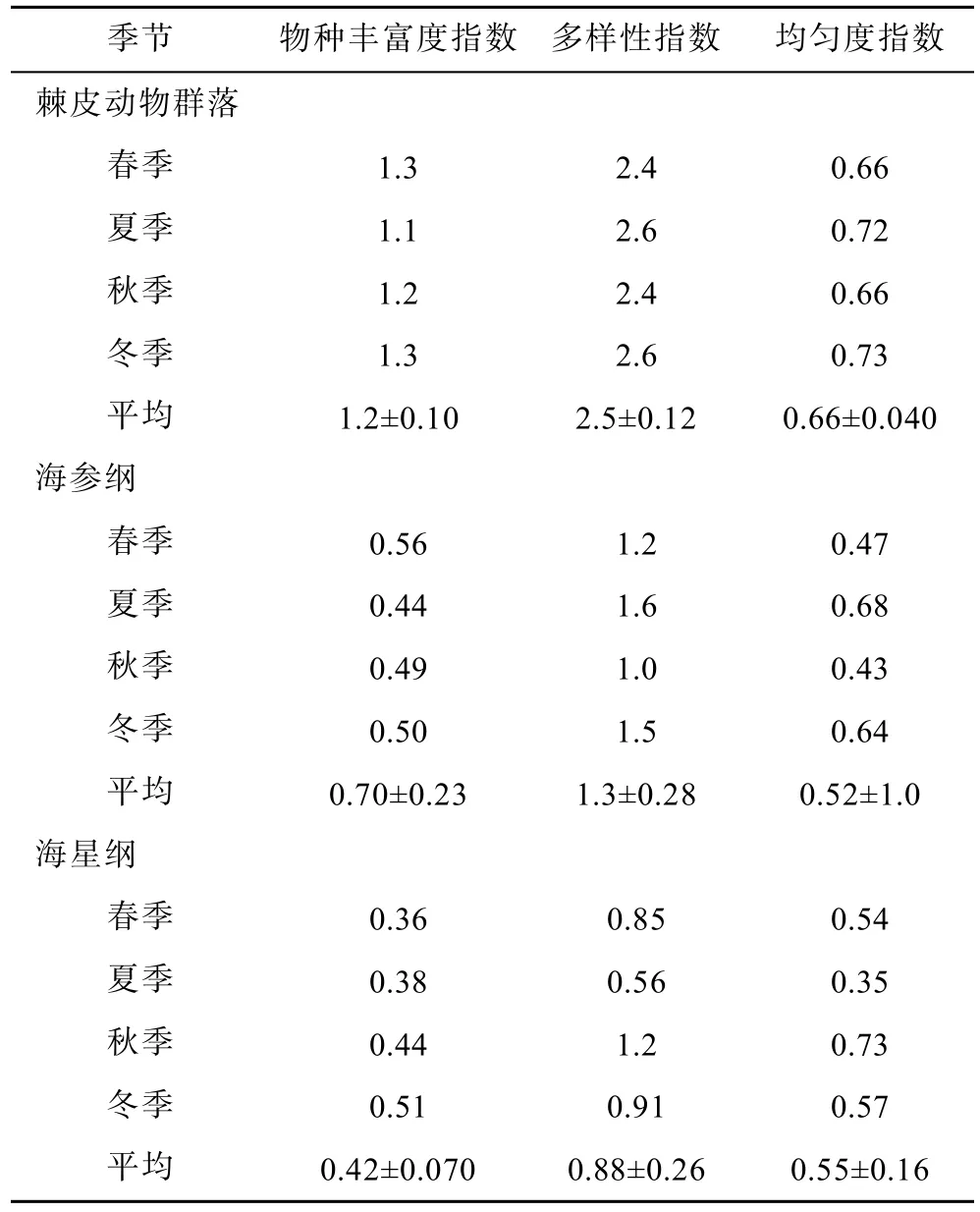
表6 棘皮動物群落生物多樣性指數季節變化Tab.6 Seasonal variation of echinoderms diversity index
3 討論
對比本研究與歷史文獻結果,三亞蜈支洲島目前棘皮動物群落種類組成更為多樣,種類多于三亞其他海域,且相較于同海域十年前顯著增加。2004到2008年,吳鐘解等(2011)等對蜈支洲島珊瑚礁區底棲動物進行了監測,發現了19種珊瑚礁動物,其中8種為棘皮動物,以海參和海星為主。本次調查結果,棘皮動物種類增加到15種,種類明顯增多,群落結構更趨多樣化。2011年西島、鹿回頭和亞龍灣3個珊瑚礁自然保護區內大型底棲動物共有166種,其中棘皮動物有11種,全部為蛇尾類(董棟等,2015)。對比南海其它島礁海域,李新正等(2007)采用輕潛采集方式調查南沙渚碧礁大型底棲動物群落特征,采集到314種珊瑚礁動物,其中棘皮動物有23種,主要種為黑櫛蛇尾Ophiocoma erinaceus和畫櫛蛇尾Ophiocoma pica。綜合上述對比可見,目前蜈支洲島棘皮動物群落種類組成較多,資源保護較好,這很可能得益于蜈支洲島旅游區嚴格的資源管護措施。
3.1 群落組成與底質分布特點
蜈支洲島海域棘皮動物種類組成中,海參綱占絕對優勢,夏季棲息密度最高,為6.5ind./10m2,特別是屬于全年優勢種的紅腹海參和黑海參,2種數量之和占全部海參總數的91.25%,分布范圍廣泛,這與該島海域底質類型更適合海參的棲息習性有著密不可分的關系。Dissanayake等(2010)研究發現,黑海參更傾向分布在長有海草的棲息地,在具有藻類和海草的巖石區域發現了高密度的紅腹海參,對特定棲息地的偏好似乎與兩種海參攝食和保護有關。現場潛水調查發現,蜈支洲島北部為泥沙、基巖以及珊瑚礁構成的錯綜復雜的生境,非常適合活動能力較弱,喜隱蔽棲息的沉積食性海參生存。
3.2 群落分布密度季節變動原因
棘皮動物棲息密度呈現出春夏高、秋冬低的特點,這可能與不同季節水溫和餌料可獲得性的波動相關。李新正等(2004)對膠州灣底棲棘皮動物的數量變化及與環境因子的關系進行了初步研究,認為棘皮動物主要分布在底質沉積類型為粉砂-黏土、有機質含量較高的區域,同時其數量變化與溫度、初級生產力關系較明顯,二者升高,其生物量、密度也較明顯升高。對蜈支洲島底棲生境的同步調查發現,筆者推測棘皮動物平均棲息密度受餌料可獲得性影響較大。如海底大型藻類(如綠藻類)僅在春季和夏初豐度較高,其釋放的有機質對底棲餌料貢獻較大,棘皮動物食物來源豐富,故棘皮動物平均棲息密度較高,但該推論尚需進一步實驗驗證。
3.3 深度對棘皮動物棲息密度的影響
不同深度棘皮動物分布結果表明,總體上深水區棘皮動物的棲息密度高于淺水區,但是差異不顯著。造成該分布差異的因素較為復雜,諸如水溫差異、底質類型、餌料適宜性以及隱蔽場所可獲得性等方面均會造成影響。一般來說,淺水區光照條件優良,底棲初級生產力水平更高,餌料獲得性高,適宜于小個體棲息。但同時受風浪影響可能更大,而深水區環境更為穩定,大個體較多棲息于此。以海參綱為例,海參在自然界常呈不均勻或者斑塊分布(Purcellet al,2009),這主要歸結于底質類型、底部沉積物特征、食物來源、深度、躲避捕食者和隱蔽場所這些因素。同時,海參對于生境存在一定的選擇偏好(Uthickeet al,1999;Mercieret al,2000;Uthickeet al,2004;Shiellet al,2010;Slateret al,2010;張宏曄等,2015)。據研究,底泥沉積物特征與深度存在著一定聯系,隨著深度增加,粉砂含量增加而礫石含量減少(Martinet al,2009),細小顆粒的沉積物更是海洋生物的營養豐富食物來源(Mayeret al,1985;Yamamotoet al,1985)。筆者推測,深水區受風浪影響較小,水流平穩,基巖和珊瑚礁等自然礁體可以更加有效地沉降自然水體中細顆粒的沉積物,同時其表面可以附著底棲微藻和其他微型生物(王吉橋等,2008;秦傳新等,2009),這有利于海參獲得營養豐富的食物。另外,Conand(2008)和Purcell等(2009)發現,紅腹海參和黑海參在珊瑚礁和巖石上會存在較大規模的集群分布,并且認為這與海參躲避波浪和急流有關。相較于淺水區,深水區水流更加平穩,受波浪影響小,且基巖和珊瑚礁組成的自然礁體更有利于海參找到隱蔽場所來躲避惡劣海況(張宏曄等,2015)。綜上所述,由于深水區具有水流平穩,受波浪影響小的特點,棲息在深水區的棘皮動物不僅可以更加有效獲得營養豐富的餌料,同時,基巖和珊瑚礁能夠為行動遲緩的棘皮動物提供重要的棲息地來躲避惡劣的海況,因此深水區分布有較多的棘皮動物。
3.4 棘皮動物生物多樣性評價
優勢種通常控制群落中的能量流和物質循環(Roy,1998),并且通過數量上的優勢以及種間關系(競爭、捕食和他感作用)以擴大對生境資源的利用,主導大型底棲動物的分布格局。穩定的優勢種種群是生態系統穩定性的重要指征。Conand等(2002)對La Re′union島調查發現17種海參,其中,黑海參為最常見的種類;Eriksson等(2012)對西印度洋(Mayotte)珊瑚礁內海參群落棲息密度和多樣性進行了研究,發現22種海參,其中黑白尼參Bohadschia atra和黑乳海參Holothuria nobilis數量最為豐富,上述海域均對應著健康穩定的珊瑚礁生態系統。對蜈支洲島全年調查中,各季節的優勢種數量穩定,為7—8種。全年優勢種的有黑海參、紅腹海參、綠刺參、呂宋棘海星、許氏大羽花和刺冠海膽,其中,屬于第一優勢種的紅腹海參在夏季優勢度達到0.52,表明蜈支洲島海域珊瑚礁生態系統保持較好的穩定性。
按Shannon-Wiener指數分級評價標準,蜈支洲島海域棘皮動物平均多樣性指數為2.5,大于2小于3,屬于輕度的人為干擾。2016年,蜈支洲島旅游區年接待游客達到254.79萬人次(周揚等,2018),在如此高的游客接待量下能保持該人為干擾程度,應得益于經營企業對珊瑚礁生態系統的有效保護與嚴格的游客管理措施。Woo等(2013)研究南海、蘇祿海和蘇拉威西海3個珊瑚礁海參群落的分布及其多樣性,采集到12種海參,對比該區域歷史研究結果,發現這3個珊瑚礁生態系統海參群落及其多樣性目前正受到威脅。調查結果發現,除了南海受到嚴格的監管和安全監測外,其他兩個海域珊瑚礁缺少執法,海參群落長期面臨過度捕撈而日益減少,加上傳統的捕魚方式,如炸魚等嚴重破壞珊瑚礁行為導致海參棲息地嚴重減少。綜合上述對比可知,良好的珊瑚礁生態系統離不開嚴格的監管和保護,蜈支洲島旅游區在對于珊瑚礁保護的管護措施非常到位,因此棘皮動物群落結構趨于多樣化。
4 結論
本研究發現,三亞蜈支洲島海域棘皮動物種類較以往研究顯著增加,群落結構趨于多樣化。沉積食性海參紅腹海參、黑海參以及牧食性刺冠海膽是主要的全年優勢種,這3個種在維護珊瑚礁生態系統物質能量循環過程將發揮重要生態作用。珊瑚礁區沉積食性的海參將覆蓋在珊瑚礁上的泥沙清除,有利于珊瑚正常生長;植食性的海膽負責攝食生長在珊瑚礁上及其周圍藻類,為珊瑚提供更多的生存空間。根據Shannon-Wiener指數分級評價標準,該海域屬于輕度干擾,人為擾動小,群落生物多樣性高。綜上所述,蜈支洲島海域珊瑚礁生態系統保護效果良好,應繼續保持當前保護力度。



附圖 蜈支洲島海域常見棘皮動物生態照片(共計15種)Photos of common echinoderm species in the waters of Wuzhizhou Island(15 species in total)
