坡壘生境調查及瀕危原因分析
2020-03-05 09:39:32方發之陳素靈桂慧穎
熱帶林業 2020年4期
關鍵詞:生長
方發之,陳素靈,桂慧穎
海南省林業科學研究院(海南省紅樹林研究院),海南海口 571100
坡壘(Hopea hainanensis)為龍腦香科常綠大喬木,是海南熱帶溝谷雨林代表種和特有種[1]。坡壘樹形優美,木質堅硬,結構致密,紋理交錯,不易變形,切面油潤,光澤性強,材色美觀,特別耐腐性強,不受蟲蛀。可與世界名材坤甸媲美,是極珍貴的工業用材和上等家具用材[2],為海南特類材。從20 世紀60 年代以來,坡壘列為禁伐樹種,在1999 年國家林業局和農業部發布的《中國國家重點保護野生植物名錄(第一批)》中,被列為國家Ⅰ級保護植物。但歷經近60 年,坡壘種群資源狀況一直得不到有效的改善,被《中國植物紅皮書》定為瀕危種,被IUCN 評估定為“瀕危滅絕”(EN)等級[3]。導致植物瀕危的原因包括外部因素(氣候、生境、過度利用和采收)和內部因素(包括遺傳力、生殖力、生活力、適應力的衰竭等)兩大類,該研究通過實地調查坡壘野外生境、種群特征及種苗生長發育情況等,分析總結導致坡壘瀕危的主要因素,為保護和人工栽培坡壘提供科學依據。
1 研究地概況
據資料記載,坡壘的野生分布區主要集中在海南中部的霸王嶺、尖峰嶺、黎母山等林區,地理坐標18°14′N~19°25′N,109°31′E~110°09′E。屬熱帶季風性海洋氣候,平均氣溫22℃,年均降水量為1900mm。但年內雨量分布不均勻,80%雨量集中在5 月~10 月,形成明顯的干濕季。年平均相對濕度為80%~85%,年平均蒸發量為1824.1mm,土壤主要為花崗巖磚紅壤[4-5]。
2 研究方法
采用文獻檢索、問詢與野外實地調查相結合的方法,對主要分布區坡壘野外種群資源進行調查。記錄其所處的地理坐標、海拔、坡度、坡位、坡向等。并以坡壘為中心,布設25m×40m 的調查樣地,對樣地內的植物進行每木調查,記錄植物種類、高度、胸徑(基徑)、冠幅、林冠、郁閉度等群落特征,并調查樣方內坡壘單株結實量、種子傳播距離及幼苗數量等。
采集成熟種子,測量種子長度、直徑、千粒重等特征,開展野外種子萌發試驗,在野外人工坡壘林下及林窗內分別設置1m×1m 的試驗小樣方,樣方內整地,播撒經催芽后的種子50 粒,設置覆蓋枯枝落葉2cm、細土2cm、細土1cm 及對照4 個不同覆蓋措施,每種處理3 個重復,35d 后調查統計種子萌發率。此外還對坡壘人工造林林分進行生長情況調查。
3 結果與分析
3.1 自然分布與伴生樹種
坡壘是龍腦香科植物,地處熱帶北緣的中國海南和越南義安有分布。在海南島主要分布于霸王嶺、尖峰嶺、黎母山、鸚哥嶺、五指山、吊羅山等熱帶雨林區,分布面積約8820hm2[6]。野外調查發現,坡壘天然分布區主要集中于海南中部山區,其中霸王嶺(266株)、尖峰嶺(31 株)、黎母山(22 株)、鸚哥嶺(5 株)。在各個調查樣地中,均發現有橄欖(Canarium album)、黃杞(Engelhardia hainanensis)、青梅(Vatica mangachapoi)、 托盤青岡 (Cyclobalanopsis patelliformis)、烏柿(Diospyros cathapensis)、長序厚殼桂(Cryptocarya metcalfiana)等的樹種伴生。此外在尖峰嶺熱帶雨林中還發現坡壘常與中華厚殼桂(Cryptocarya chinensis)、紅錐(Castanopsis hystrix)、卵葉桂 (Cinnamomum rigidissimum)、 雞毛松(Podocarpus imbricatus)、 吊皮錐(Castanopsis kawakamii)、大葉蒲葵(Livistona saribus)等混生。
3.2 生境特征
3.2.1 氣候
氣溫、水、日照等氣象因子是決定植物分布的重要因子。根據收集的氣象資料統計,坡壘分布區為典型的熱帶海洋性季風氣候,分布區年均氣溫為21.5℃,最熱月均溫22.8℃(5 月),極端高溫37.5℃,在冬季1 月均溫12.2℃,極端低溫1.1℃;年均降雨量1650mm~1900mm,平均降雨日數100d。干濕季分明,5 月~10 月為雨季,11 月~次年4 月為旱季。季風對坡壘分布區的降水量分配影響很大,使月、季、時段降水量出現明顯的差異。坡壘在1.1℃能正常生長,具有一定抗寒能力,但在氣溫驟降到-1℃時死亡[7]。
3.2.2 海拔
坡壘分布的海拔范圍大致在300m~1100m 之間,其在600m~700m 海拔范圍內分布最多,表明坡壘的分布范圍較廣、生態幅較寬[8]。在人工種植中,坡壘在海南的樂東、定安、澄邁、儋州等海拔低于100m 的臺地均能正常生長。
3.2.3 地形因子
對霸王嶺(266 株)坡壘的實地調查發現,長于山脊的有48 株,占18.0%;山谷的有96 株,占36.1%;溪澗兩旁邊的有122 株,占45.9%。其中山谷與溪澗共218 株,占82%,說明坡壘喜生于溫暖靜風,空氣濕度較大的山谷與溪澗邊。
3.3 種群特征
調查發現,野生坡壘成材樹(胸徑≥20cm)均單株零星地分布于天然林中,未發現片林形成的群落。324 株成材坡壘均處于所在群落的最頂層,說明坡壘在其所在的群落種間竟爭較強。此外調查還發現,散落的坡壘種子具有較高萌發率,坡壘母樹周邊約50m~100m 范圍內,株高3cm~5cm 的幼苗總數約有280 株~510 株,幼苗密度最高達36 株/m2。樹高0.5m~1.5m 的幼樹僅有2 到3 株,均生長于石頭縫隙或石頭下方。說明在熱帶雨林中,由于林木和雜草灌木茂密,種間竟爭激烈,坡壘幼苗競爭和適應能力弱,易被淘汰,而石頭周邊林木和雜草灌木稀疏,能提供坡壘生長空間,并且坡壘在石縫有較強適應能力,這與上述有68.7%的成材樹分布石頭邊或石縫基本相吻合。調查中則未發現高1.5m 以上的坡壘未成材植株,可推測坡壘野外自然更新能力較弱。
3.4 繁育特征
人工種植的坡壘7a 可開花結果,花期為5 月至10 月上旬,種子成熟期為次年3 月上旬到5 月上旬。坡壘種子為堅果類,帶翅。種子產量存在大小年之分,大年時單株產果量達10kg~30kg。種子近似心形,長約1.1cm~1.6cm,直徑0.7cm~1.2cm,帶萼片千粒重1025g~1109g,除去萼片千粒重650g~849g。種子成熟后易脫落,隨風飄落。飄落至陰濕環境中的種子在5d 后可發芽。至開闊干燥之處,種子則失去水分難以萌發。60%~70%種子飄落在距母株60m 內,表明坡壘種子雖帶翅,但其種子質量相對重,傳播距離有限。
不同郁閉度不同覆蓋措施對坡壘種子發芽率的影響結果見表1。
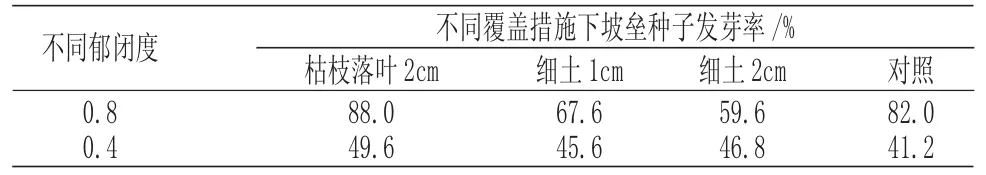
表1 不同郁閉度及覆蓋措施下坡壘種子發芽率Tab.1 The seed germination rate of Hopea hainanensis under different canopy density and mulching measures
由表1 可知,所有處理中,坡壘林下被枯枝落葉覆蓋2cm 的坡壘種子發芽率最高,郁閉度0.4 下不覆蓋的坡壘種子發芽率最低。郁閉度對坡壘種子萌發有顯著影響,郁閉度0.8 的各項措施發芽率均高于郁閉度0.4 下發芽率。這可能與萌發溫度有關,郁閉度小的區域光照更強,蒸發量更大,坡壘種子經照曬脫水后失去生活力,導致種子未能萌發。此外對比不同郁閉度下的各項覆蓋措施發芽率均可發現,枯枝落葉覆蓋2cm 處理的發芽率均高于對照處理,而細土覆蓋則表現為在坡壘林下發芽率低于對照,在林窗下發芽率高于對照。這可能與枯枝落葉層質量較輕、疏松多孔、空氣流通良好有關,且適當的覆蓋使得坡壘種子避免陽光直射而過分失水。
3.5 生長特征
從表2 可看出,坡壘幼苗在郁閉度0.4 的以上林下生長緩慢,并隨著郁閉度的增加而降低,而保存率則相反,說明幼苗生長需要陽光,但光照強的地方種間競爭激烈,坡壘幼苗容易被淘汰。在東方市江邊造林中采用不套種不撫育措施的保存率逐年降低,同樣也說明這個問題。從東方市江邊(套種農作物)的調查結果來看,人工種植的坡壘3a 后,其年生長旺盛,第4 年至第7 年,樹高年均生長量為0.9m~1.3m。楊德軍等[9]對42 年生坡壘生長觀測也發現,坡壘樹高、胸徑生長速度較為均勻,其生長速度沒有出現較大波動,最大高年生長量0.95m,最大胸徑年生長量1.4cm。因此,坡壘能成片林成長,并有長時期旺盛的生長力。
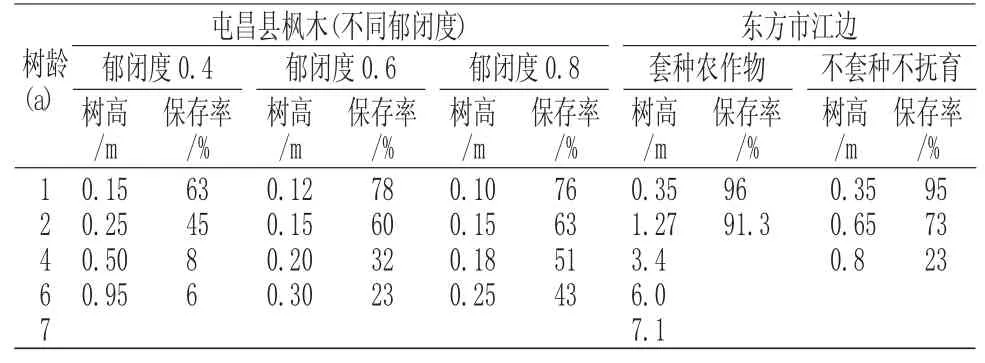
表2 不同造林區域及條件下的坡壘生長情況Tab.2 The growth of Hopea hainanensis in different afforestation areas and conditions
3.6 人類活動的影響
(1)毀林開墾種植農作物,導致坡壘生境萎縮。解放初期海南原始熱帶雨林面積為120 萬hm2,森林覆蓋率35.5%。從20 世紀50 年代起,由于過量采伐、毀林墾植、毀林開荒、亂砍濫伐和森林火災等人為破壞,至1978 年森林資源清查時天然林僅存40萬hm2,森林覆蓋率12.0%。1987 年下降到24.4 萬hm2,森林覆蓋率7.2%。到1999 年海南原始熱帶雨林面積僅13.3 萬hm2,森林覆蓋率4%。58%的天然林郁閉度從0.8 下降至0.4~0.5。大面積原始熱帶雨林被開墾和破壞,使坡壘的生境逐步退化喪失和碎片化,導致繁殖能力下降,種群數量不斷減少。
(2)坡壘被偷砍盜伐,成材樹數量銳減。坡壘木材材質堅硬,耐腐力極強,是極珍貴的上等家具用材。2012 年至2013 年間,黑市上交易價格達100 元/kg 至200 元/kg,在經濟利益的驅使下不法分子偷砍濫伐,導致野生坡壘母株數量銳減。近些年來,執法部門對偷砍濫伐行為進行嚴厲打擊,偷砍盜伐的行為受到厄制,坡壘資源得到有效保護。
4 瀕危原因
造成物種瀕危的外界因素主要為生境退化喪失、人類對生物資源的過度開采利用、外來種入侵、全球氣候變化等,以及物種繁殖力低、種群小與種群密度低、地理分布狹窄、棲息地特化等與植物自身相關的易導致其瀕危的滅絕特征[10-11]。坡壘在海南的分布范圍較廣、生態幅較寬,同時有較大結實量,種子發芽率較高,因此不存在自身繁殖能力上的問題。根據調查研究結果,導致坡壘瀕危的原因歸結為下列4 點:
(1)人類破壞是導致坡壘瀕危的主要原因,從1952 年至1999 年間,由于不合理的人類活動,海南原始熱帶雨林面積從120 萬hm2銳減至13.3 萬hm2。近90%的熱帶雨林遭受破壞,坡壘群落作為熱帶雨林頂級群落代表之一,其生境的劇烈收縮,加之不法分子的偷砍濫伐,使坡壘母株數量銳減。
(2)坡壘生長特性也是導致瀕危的原因之一,在熱帶雨林中各物種種間競爭較為激烈,而坡壘幼苗期生長極其緩慢,雖有大量幼苗分布,但坡壘幼苗競爭力弱易被淘汰,導致僅存的成年坡壘植株較少,種群野外自然更新能力弱。
(3)種群密度低,致使種群自然恢復與發展緩慢。野生坡壘成材樹均單株零星地分布于天然林中,未發現片林形成的群落,在8820hm2的范圍內,調查僅發現324 株成材樹,種群密度極低,導致種群繁殖能力與抗逆性弱,種群自然恢復與發展緩慢。
(4)坡壘種子及其萌發特性制約種群擴張。坡壘種子為帶翅堅果,但種子質量相對較重傳播距離有限,其種子集中脫落分布在距母株100m 的范圍內,限制其種群生長范圍的擴張。同時種子含水率高,在陰濕環境下適宜發芽,開闊干燥環境下則易被曬干而失去萌芽力,坡壘種子在自然環境下對萌芽條件有較高要求,制約種群的發展。
5 保護及發展建議
(1)加強坡壘的保護與宣傳,恢復其生境與種群。一是加強啟動坡壘專項保護,以林業主管部門為主體,開展野外極小種群保護行動,優先關注和保護坡壘等瀕危物種。建立自然保護區和保護小區是有效保護瀕危物種的方法之一[12],在集中分布地建立重點保護小區,定期巡視以保護種群生境,維護種群穩定,降低坡壘因種群過小而滅絕的風險。二是打擊偷砍盜伐行為,保護好坡壘成材母樹。三是積極開展珍稀瀕危植物的宣傳教育工作,增強公眾保護意識和各渠道宣傳是保護珍稀瀕危植物的有效手段之一[13],相關職能部門應充分利用各種媒體,采用多種形式,廣泛開展宣傳教育活動,普及保護珍稀瀕危植物的重要性及方式方法,在民眾中樹立保護共識,提高參與保護行動積極性,使坡壘等珍稀瀕危植物保護工作成為民眾的自覺行為。
(2)加大人工造林力度,擴大種群資源。坡壘是特類用材樹種,其材質好,用途廣,經濟價值極高,是優良的國家儲備林樹種,且人工造林的純林林分生長快,是今后海南發展珍貴鄉土樹種的主要造林樹種。建議采取以下措施擴大坡壘種群資源:一是大力發展以坡壘等珍貴鄉土樹種代替生態、經濟效益低下的多代桉樹林分的更新改造模式。二是在重要生態區位的桉樹等人工公益林內,應用坡壘等珍貴鄉土樹種進行補植補造。三是將坡壘作為四旁綠化樹種進行推廣種植。坡壘是常綠喬樹種,其冠形優美,應用于園林綠化,既可提升綠化成效,又可增加優質木材儲備,擴大坡壘種群數量,保存坡壘種質資源。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
