中國(guó)主栽櫻花的分子系統(tǒng)發(fā)育分析
2020-03-05 04:05:40王志龍林樂(lè)靜袁冬明
植物研究 2020年6期
付 濤 王志龍* 林樂(lè)靜 林 立 李 文 袁冬明
(1. 寧波城市職業(yè)技術(shù)學(xué)院/浙江園林綠化技術(shù)協(xié)同創(chuàng)新中心,寧波 315100;2. 寧波市鄞州區(qū)林業(yè)技術(shù)管理服務(wù)站,寧波 315100)
櫻花為櫻屬(Cerasus)植物栽培品種的統(tǒng)稱(chēng),經(jīng)過(guò)幾百年的培育,已有幾百個(gè)品種,目前國(guó)內(nèi)主栽的櫻花大多數(shù)是日本品種,很多景區(qū)、公園、學(xué)校、街道甚至是家庭都有日本櫻花的種植,最常見(jiàn)的有染井吉野櫻(俗稱(chēng)早櫻)和關(guān)山櫻(俗稱(chēng)晚櫻),然而,有許多品種親本來(lái)源不明、種系定位不一、命名不規(guī)范等諸多問(wèn)題,不利于櫻花產(chǎn)業(yè)的健康發(fā)展[1~3]。我國(guó)擁有豐富的野生資源,遠(yuǎn)超日本和朝鮮等國(guó)家[4],然而我國(guó)重視食用櫻桃的栽培,對(duì)具有觀賞性的櫻屬植物關(guān)注甚少,育成品種更少,但近幾年國(guó)內(nèi)“櫻花熱”持續(xù)升溫,對(duì)自主櫻花品種的培育也在如火如荼地進(jìn)行中,依托我國(guó)豐富的野生資源(我國(guó)約50 種櫻屬植物),不久的將來(lái),將會(huì)擁有一大批具有我國(guó)自主知識(shí)產(chǎn)權(quán)的櫻花新品種,逐漸將日本品種替換成我國(guó)自主栽培櫻花品種[5]。
日本人Kawasaki 將日本櫻花分為7 個(gè)群,得到了國(guó)際社會(huì)的一致認(rèn)可,但對(duì)于雜交或來(lái)源不明的品種分類(lèi)和鑒定有一定的局限性[6~7]。國(guó)內(nèi)南京林業(yè)大學(xué)王賢榮團(tuán)隊(duì)提出了櫻花品種分類(lèi)的五級(jí)標(biāo)準(zhǔn),但該標(biāo)準(zhǔn)易受生長(zhǎng)環(huán)境、栽培管理、砧木選擇等因素的影響,其應(yīng)用也受到一定的影響[8]。因此,單純依靠形態(tài)分類(lèi)很難將所有品種正確分類(lèi),需要借助分子手段加以輔助鑒定。目前,DNA 條形碼廣泛應(yīng)用于動(dòng)植物的分類(lèi)鑒定研究[9~17],并取得了很好的結(jié)果,其中核DNA 序列ITS 和葉綠體DNA 序列rbcL、matK 以及psbA-trnH應(yīng)用比較多,但rbcL 和matK 為葉綠體功能基因,序列變異較小,因此主要應(yīng)用于屬及以上水平的分類(lèi)鑒定研究,而ITS 和psbA-trnH 主要由非功能基因構(gòu)成,序列變異相對(duì)較大,因此可應(yīng)用于種及種以水平的分類(lèi)鑒定研究[18-19]。本研究利用ITS和psbA-trnH 對(duì)國(guó)內(nèi)引進(jìn)的櫻花品種進(jìn)行測(cè)序分析,探究彼此間的分子差異,構(gòu)建系統(tǒng)發(fā)育樹(shù),旨在為日本櫻花的分類(lèi)和種系定位以及糾正形態(tài)學(xué)分類(lèi)的不足甚至錯(cuò)誤提供分子依據(jù)。
1 材料與方法
1.1 材料
2016~2017年從寧波、武漢和上海等地采集了88個(gè)櫻花品種樣品,采集不同品種櫻花的健康、無(wú)病害的幼葉,硅膠處理,4℃保存,詳細(xì)樣品信息見(jiàn)表1。

表1 樣品信息Table 1 Samples information and source of the 88 cherry blossoms

續(xù)表1Continued table 1

續(xù)表1Continued table 1
1.2 方法
利用試劑盒(上海萊楓生物科技有限公司)進(jìn)行88個(gè)樣品的總DNA提取,提取的DNA稀釋后放入冰箱-20℃保存、備用。
選用核基因ITS和葉綠體基因trnH-psbA 進(jìn)行DNA條形碼分析,2個(gè)DNA片段引物序列詳見(jiàn)表2。PCR 反應(yīng)體系為50μL,其中含預(yù)混合液25μL,模板DNA 1μL,引物對(duì)4μL,最后補(bǔ)加ddH2O 20μL。PCR擴(kuò)增程序:95℃預(yù)變性5 min;95℃變性30 s,55或58℃退火30 s(ITS 為58℃;trnH-psbA 為55℃),72℃延伸1 min,45個(gè)循環(huán);最后72℃延伸7 min。擴(kuò)增完成后直接取5μL PCR產(chǎn)物進(jìn)行點(diǎn)樣,進(jìn)行瓊脂糖凝膠電泳(1.5%),驗(yàn)證擴(kuò)增結(jié)果,之后將有目標(biāo)條帶的PCR擴(kuò)增產(chǎn)物送到測(cè)序公司進(jìn)行測(cè)序。
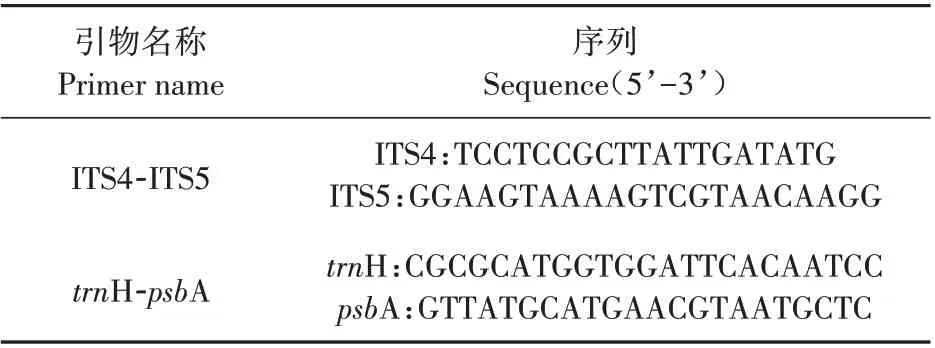
表2 DNA條形碼引物名稱(chēng)及其序列Table 2 Primer names and sequences of DNA barcoding
1.3 數(shù)據(jù)分析
在NCBI 中搜索有關(guān)櫻屬植物的ITS 和trnHpsbA 的全長(zhǎng),以此作為標(biāo)準(zhǔn)序列,再將測(cè)序得到的序列運(yùn)用DNAman 軟件與標(biāo)準(zhǔn)序列進(jìn)行比對(duì),以確認(rèn)測(cè)序的準(zhǔn)確性,然后對(duì)合格的序列進(jìn)行修剪,獲得完整的片段序列。利用MEGA 6.0 軟件計(jì)算出88 份材料的ITS 和trnH-psbA 各基因序列的組成、G+C 含量(%)等,此外還計(jì)算了各序列中的堿基的變異位點(diǎn)率和信息位點(diǎn)率以及轉(zhuǎn)換/顛換比率等,并通過(guò)2 000 次重復(fù)的bootstrap 值來(lái)確定每個(gè)分支的支持率,構(gòu)建鄰接樹(shù)(NJ),選擇K2-P 模型遺傳距離。
2 結(jié)果與分析
2.1 88 個(gè)櫻花品種ITS 和trnH-psbA 序列特點(diǎn)分析
由表3可知,ITS序列長(zhǎng)度在605~606 bp,變異很小,其中僅有寒緋櫻、才力櫻、大漁櫻、橫濱緋櫻和陽(yáng)光櫻的ITS 序列長(zhǎng)度為605 bp,其余品種ITS序列長(zhǎng)度皆為606 bp。trnH-psbA序列長(zhǎng)度在288~306 bp,其中內(nèi)里櫻trnH-psbA 序列長(zhǎng)度為289 bp,江戶(hù)彼岸櫻、染井吉野櫻、小松乙女櫻、神代曙櫻、澳博拉大葉早櫻、咲耶姬櫻、御帝吉野櫻、陽(yáng)春櫻、白花真櫻、赤實(shí)大島櫻、寒緋櫻和鵯櫻trnH-psbA序列長(zhǎng)度為296 bp,陽(yáng)光櫻trnH-psbA 序列長(zhǎng)度為306 bp,其余品種trnH-psbA序列長(zhǎng)度均為288 bp。
由表3 還可知,ITS 的G+C 平均百分含量較高(59.6.%),而trnH-psbA 的G+C 平均百分含量卻較低(21.8%);ITS 和trnH-psbA 的保守位點(diǎn)百分含量相差不大(分別為94.6%和94.5%),而變異位點(diǎn)百分含量相差較大,其中ITS 為5.3%(簡(jiǎn)約信息位點(diǎn)為3.3%,單突變位點(diǎn)為2.0%),而trnH-psbA 僅為2.0%(簡(jiǎn)約信息位點(diǎn)為0.7%,單突變位點(diǎn)為1.3%),ITS 的簡(jiǎn)約信息位點(diǎn)百分含量顯著高于trnH-psbA;ITS插入/缺失序列最少,僅為1個(gè)堿基,而trnH-psbA 高達(dá)19 個(gè)堿基;此外,ITS 轉(zhuǎn)換/顛換比率為0.7,而trnH-psbA卻為0。
2.2 88個(gè)櫻花品種變異位點(diǎn)分析
88 個(gè)櫻花品種通過(guò)ITS 序列比對(duì)得到32 個(gè)變異位點(diǎn),其中簡(jiǎn)約信息位點(diǎn)有20個(gè),單突變位點(diǎn)有12 個(gè)。通過(guò)trnH-psbA 序列比對(duì)得到6 個(gè)變異位點(diǎn),其中簡(jiǎn)約信息位點(diǎn)有2 個(gè),單突變位點(diǎn)有4個(gè)。由表4 可知,ITS 變異位點(diǎn)中有18 個(gè)是顛換,有14個(gè)是轉(zhuǎn)換,顛換要高于轉(zhuǎn)換,其中顛換中A 突變成C、C 突變?yōu)锳 和G 突變成T 出現(xiàn)頻率相對(duì)較高,轉(zhuǎn)換中G 突變成T 出現(xiàn)頻率較高。trnH-psbA 的6 個(gè)變異位點(diǎn)都是顛換,無(wú)轉(zhuǎn)換。由此說(shuō)明88 個(gè)櫻花品種的ITS 的變異程度明顯高于trnH-psbA,ITS 的分辨率要明顯高于trnH-psbA,ITS可作為櫻花品種系統(tǒng)發(fā)育研究、品種鑒定等的核心條形碼,而trnH-psbA可作為輔助序列予以應(yīng)用。
2.3 88 個(gè)櫻花品種ITS 和trnH-psbA 插入/缺失序列分析
本研究共得到ITS 的排列位點(diǎn)607 個(gè),在排列位點(diǎn)437 處,寒緋櫻含有1 個(gè)單堿基(G)的插入。trnH-psbA 有307 個(gè)排列位點(diǎn),在41~58 排列位點(diǎn)處,陽(yáng)光含有18 個(gè)堿基(CTTGTAAGTTTATCATTA)的插入;在51~58 排列位點(diǎn)處,澳博拉大葉早櫻、白花真櫻、赤實(shí)大島櫻、寒緋櫻、江戶(hù)彼岸櫻、染井吉野櫻、神代曙櫻、小松乙女櫻、咲耶姬櫻、鵯櫻、陽(yáng)春櫻和御帝吉野櫻含有8 個(gè)堿基(TATCATTA)的插入,此外,在202 排列位點(diǎn)處,內(nèi)里櫻含有1個(gè)單堿基(A)的插入。

表3 序列堿基組成及變異率Table 3 Sequence base composition and mutation rate

表4 變異位點(diǎn)分析Table 4 Analysis of variable
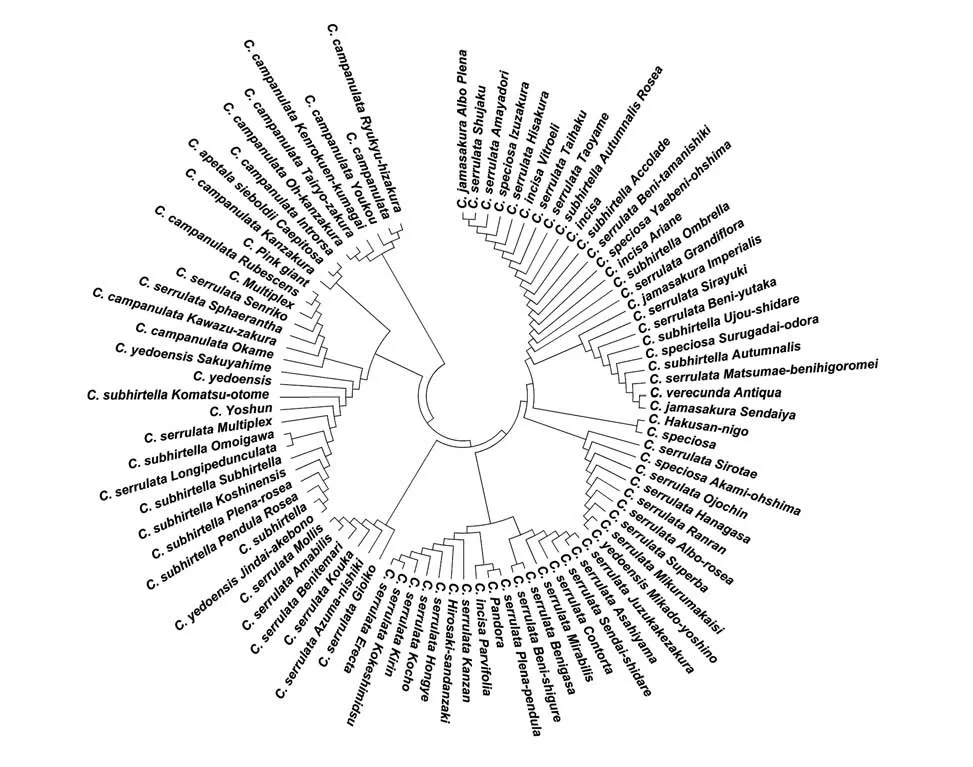
圖1 基于ITS+trnH-psbA組合序列構(gòu)建88個(gè)櫻花品種的NJ樹(shù)Fig. 1 The NJ tree of the 88 cherry blossoms with the concatenated ITS+trnH-psbA sequencesT
2.4 88個(gè)櫻花品種的系統(tǒng)發(fā)育分析
利用分析軟件MEGA 6.0 構(gòu)建了88 個(gè)櫻花品種的系統(tǒng)發(fā)育樹(shù)(見(jiàn)圖1),由圖1 可知,88 個(gè)櫻花品種可分為4 大類(lèi)(Ⅰ、Ⅱ、Ⅲ和Ⅳ),其中第Ⅰ類(lèi)有Ⅰa、Ⅰb 和Ⅰc 三小類(lèi),Ⅰa 包含市原虎尾櫻、朱雀櫻、雨宿櫻、伊豆櫻、一葉櫻、維多利富士櫻、太白櫻、手弱女櫻、十月紅櫻、三島富士櫻、嘉獎(jiǎng)櫻、紅玉錦櫻、八重紅大島櫻、阿麗亞娜富士櫻、澳博拉大葉早櫻、郁金櫻、內(nèi)里櫻、白雪櫻、紅豐櫻、雨晴枝垂櫻、駿河臺(tái)香櫻、十月櫻、松前紅緋衣櫻、奈良八重櫻和仙臺(tái)屋櫻,Ⅰb 包含白山2 號(hào)櫻和大島櫻,Ⅰc 包含白妙櫻、赤實(shí)大島櫻、大提燈櫻、花笠櫻、蘭蘭櫻、普賢象櫻、松月櫻、御車(chē)返櫻和御帝吉野櫻;第Ⅱ類(lèi)有Ⅱa 和Ⅱb 兩小類(lèi),Ⅱa 含有梅護(hù)寺數(shù)珠掛櫻、旭山、朝日山櫻、山櫻枝垂櫻、福祿壽櫻、大村櫻、紅笠櫻和紅時(shí)雨櫻,Ⅱb 含有菊垂櫻、潘多拉櫻、冬櫻、關(guān)山櫻、弘前三段咲櫻、紅葉櫻、蝴蝶櫻、麒麟櫻、苔清水櫻和天川櫻;第Ⅲ類(lèi)有御衣黃櫻、東錦櫻、紅華櫻、紅手毬櫻、日暮櫻和楊貴妃櫻;第Ⅳ類(lèi)有Ⅳa、Ⅳb、Ⅳc 和Ⅳd 4 小類(lèi),Ⅳa 含有江戶(hù)彼岸櫻、神代曙櫻、紅枝垂櫻、八重紅枝垂櫻、越之彼岸櫻、小彼岸櫻、鵯櫻、思川櫻、白花真櫻、陽(yáng)春櫻、小松乙女櫻、染井吉野櫻和咲耶姬櫻,Ⅳb 含有才力櫻、河津櫻、兼六園菊櫻、千里香櫻、青肌櫻、修繕?biāo)潞畽押头劬奕藱眩鬰含有寒櫻、高妙櫻和初美人櫻,Ⅳd含有大寒櫻、大漁櫻、橫濱緋櫻、陽(yáng)光櫻、寒緋櫻和琉球寒緋櫻。
3 討論
對(duì)于確定的植物品種,一般均采用扦插[20~21]、嫁接[22~23]或組培[24~25]等無(wú)性繁殖進(jìn)行擴(kuò)大再生產(chǎn),才能保持特定品種的穩(wěn)定性狀,其遺傳物質(zhì)完全不會(huì)改變(排除基因突變),因此,可以通過(guò)比較不同櫻花品種的基因差異用于品種鑒定。本研究中僅有寒緋櫻群里的寒緋櫻、才力櫻、大漁櫻、橫濱緋櫻和陽(yáng)光櫻等5個(gè)品種的ITS序列長(zhǎng)度為605 bp,其余83 個(gè)品種的ITS 序列長(zhǎng)度均為606 bp,此外,trnH-psbA 序列為基因間隔區(qū)域,進(jìn)化選擇壓力較小,其長(zhǎng)度變異較大,往往有大量插入/缺失片段,根據(jù)結(jié)果可知,陽(yáng)光櫻的trnH-psbA 序列長(zhǎng)度為306 bp,內(nèi)里櫻trnH-psbA 序列長(zhǎng)度為289 bp,江戶(hù)彼岸群的江戶(hù)彼岸櫻、染井吉野櫻、小松乙女櫻、神代曙櫻、澳博拉大葉早櫻、咲耶姬櫻、御帝吉野櫻、陽(yáng)春櫻和山櫻群的白花真櫻、赤實(shí)大島櫻、鵯櫻以及寒緋櫻群的寒緋櫻的trnH-psbA 序列長(zhǎng)度為289 bp。因此,可依據(jù)DNA 片段長(zhǎng)度多態(tài)性對(duì)部分品種進(jìn)行分子鑒定。
核基因ITS 包含ITS1、5.8S 和ITS2,其中ITS1和ITS2 為非編碼區(qū),而基因非編碼區(qū)所承受的選擇壓力較小,因此,ITS 突變位點(diǎn)(簡(jiǎn)約信息位點(diǎn)和單突變位點(diǎn))多位于ITS1 和ITS2[26]。葉綠體DNA呈單親遺傳(母系遺傳),不存在像核基因那樣的基因重組,編碼基因往往非常保守,但像trnH-psbA等基因間隔序列變異較大,但變異率往往要遠(yuǎn)低于核基因[27]。本研究ITS 變異位點(diǎn)百分率(5.3%)遠(yuǎn)高于trnH-psbA(2.0%),因此ITS 在櫻屬植物種或種下水平的鑒定率顯著高于trnH-psbA,ITS可作為核心條形碼序列用于系統(tǒng)發(fā)育研究,trnH-psbA可作為輔助序列予以應(yīng)用。此外,插入/缺失現(xiàn)象在葉綠體基因非編碼區(qū)較常出現(xiàn),如陽(yáng)光櫻有18個(gè)堿基(CTTGTAAGTTTATCATTA)的插入,江戶(hù)彼岸群的大多數(shù)品種都有8 個(gè)堿基(TATCATTA)的插入,根據(jù)這些特定序列長(zhǎng)度也可用于品種的分子鑒定。
本研究88 個(gè)品種均引自于日本,而日本的櫻花品種其原始親本均來(lái)自9 個(gè)野生種[山櫻(Cerasus serrulata)、大山櫻(Cerasus sargentii)、霞櫻(Cerasus verecunda)、江戶(hù)彼岸(Cerasus subhirtella)、豆櫻(Cerasus incisa)、大島櫻(Cerasus speciosa)、高嶺櫻(Cerasus nipponica)、深山櫻(Cerasus maximowiczii)和丁字櫻(Cerasus apetala)]以及2 個(gè)栽培種[寒緋櫻(Cerasus campanulata)和櫻桃(Cerasus pseudocerasus)],尤其是日本晚櫻大多是反復(fù)雜交所得,其遺傳背景比較復(fù)雜,很多日本晚櫻品種親本不詳[7]。日本人Kawasaki 在本田正次和林彌榮[28]分類(lèi)標(biāo)準(zhǔn)基礎(chǔ)之上,按種系將日本櫻花品種分為7 個(gè)群,即山櫻群、江戶(hù)彼岸群、寒緋櫻群、豆櫻群、丁字櫻群、櫻桃群、深山櫻群。本研究有54個(gè)品種屬于山櫻群,有18個(gè)品種屬于江戶(hù)彼岸群,有12 個(gè)品種屬于寒緋櫻群,有4 個(gè)屬于豆櫻群,依據(jù)ITS+trnH-psbA構(gòu)建的系統(tǒng)發(fā)育樹(shù)的結(jié)果,Ⅰ、Ⅱ和Ⅲ主要是山櫻群的櫻花品種,Ⅳ主要是江戶(hù)彼岸群和寒緋櫻群的櫻花品種,而豆櫻群的品種不構(gòu)成單系,與山櫻群混合在一起。因此,利用ITS+trnH-psbA可以較好地將不同品種的櫻花品種合理歸類(lèi),作為形態(tài)分類(lèi)的輔助手段。
大島櫻為日本原生種,因其花最大,花量最多,且花朵帶有香味,被廣泛喜愛(ài),培育出的園藝品種也最多(包括變異品種和雜交品種)[1],我國(guó)所說(shuō)的日本晚櫻其實(shí)指的就是具有大島櫻血統(tǒng)的櫻花雜交品種,因此,日本晚櫻中的雜交品種拉丁文本應(yīng)為Cerasus speciosa,但國(guó)內(nèi)習(xí)慣將日本晚櫻花拉丁文描述為Cerasus serrulata,因此本文對(duì)日本晚櫻中的雜交品種采用了后一種描述。山櫻群的櫻花品種比較復(fù)雜,其絕大多數(shù)品種原始親本是大島櫻、山櫻、大山櫻和霞櫻,然后通過(guò)彼此間反復(fù)雜交形成了今天諸多的山櫻群櫻花品種,從系統(tǒng)發(fā)育樹(shù)可知,它們彼此間的系統(tǒng)發(fā)育關(guān)系比較混亂,無(wú)規(guī)律可循。在第Ⅰ類(lèi)的Ⅰa 中包含3 個(gè)豆櫻(變異)品種,說(shuō)明ITS+trnH-psbA 組合序列對(duì)豆櫻、山櫻(紅山櫻)和大島櫻的分辨率較低,而冬櫻為雜交品種,其系統(tǒng)發(fā)育關(guān)系與豆櫻變異品種相差較大。江戶(hù)彼岸群大部分品種能夠單獨(dú)構(gòu)成一系,但其中有部分雜交品種混入到山櫻群,如十月紅櫻、嘉獎(jiǎng)櫻、澳博拉大葉早櫻、雨晴枝垂櫻、十月櫻、御帝吉野櫻和潘多拉櫻,因此,這些雜交品種遺傳物質(zhì)更多受到山櫻、大山櫻、大島櫻、霞櫻和豆櫻的影響,這7個(gè)江戶(hù)彼岸群雜交品種分類(lèi)地位有待進(jìn)一步確認(rèn),此外,江戶(hù)彼岸群還混有白花真櫻和鵯櫻,這2 個(gè)品種可能具有江戶(hù)彼岸櫻的血統(tǒng)。寒緋櫻群的12 個(gè)品種全部單獨(dú)構(gòu)成一系,但其中混有青肌櫻、千里香、兼六園菊櫻和高砂櫻,其中青肌櫻為我國(guó)原生種(山櫻),與寒緋櫻關(guān)系較近,其余兼六園菊櫻和高砂櫻均是花色淡紅的重瓣櫻花,推測(cè)均具有寒緋櫻血統(tǒng),而千里香花色潔白又與青肌櫻較近,且香氣濃郁,推測(cè)其是大島櫻或其品種與寒緋櫻品種的雜交品種。本研究中系統(tǒng)不確定的白山2號(hào)與大島櫻關(guān)系最近,可將其歸類(lèi)為大島櫻影響的日本晚櫻,其拉丁名可更改為Cerasus serrulata‘Hakusan-nigo’,弘前三段咲櫻拉丁名可更改為Cerasus serrulata‘Hirosaki-sandanzaki’,陽(yáng)春櫻拉丁名可更改為Cerasus subhirtella‘Yoshun’,粉巨人櫻拉丁名可更改為Cerasus campanulata‘Pin giant’,而潘多拉櫻與山櫻群混在一起,因此建議將其拉丁名更改為Cerasus serrulata‘Pandora’。白雪櫻一般認(rèn)為是江戶(hù)彼岸櫻反復(fù)雜交所得,這與本團(tuán)隊(duì)前期利用ISSR 分子標(biāo)記法所得結(jié)果一致,但本研究結(jié)果與前期研究結(jié)果有所出入,表明利用ITS 和trnH-psbA 兩個(gè)條形碼序列不能將白雪正確分類(lèi),做出了錯(cuò)誤的結(jié)論,也可能是白雪櫻遺傳背景比較復(fù)雜,也表明DNA 條形碼用于物種系統(tǒng)發(fā)育有其不足和缺陷。
ITS 和trnH-psbA 可用于種以及以上水平的系統(tǒng)發(fā)育研究,可以取得較好結(jié)果,但對(duì)于種下水平結(jié)果往往不理想[29]。本研究結(jié)果表明,山櫻群中的各系櫻花品種彼此混亂的聚合在一起,尤其是各類(lèi)的日本晚櫻,除了山櫻群大部分品種遺傳背景比較復(fù)雜以外,還可能是因?yàn)樯綑讶旱母飨灯贩N其原始親本系統(tǒng)發(fā)育關(guān)系本就很近造成的,這也是雜交育種的基礎(chǔ),因此,我們推測(cè)日本的山櫻、大山櫻、大島櫻、霞櫻、豆櫻等原種為近期分化出的物種,系統(tǒng)發(fā)育關(guān)系較近,并未完全出現(xiàn)生殖隔離。但很多江戶(hù)彼岸群和寒緋櫻群的櫻花品種很多也是與山櫻、大山櫻、大島櫻、霞櫻、豆櫻等雜交所得,但能夠與山櫻群品種彼此分離,由此可知,江戶(hù)彼岸櫻和寒緋櫻與山櫻、大山櫻、大島櫻、霞櫻、豆櫻系統(tǒng)發(fā)育關(guān)系相對(duì)較遠(yuǎn),但日本櫻屬植物原種彼此間都能夠雜交形成可育后代,嚴(yán)格來(lái)說(shuō)都可并為一個(gè)種,但形態(tài)分類(lèi)專(zhuān)家依據(jù)其形態(tài)差異分為了不同種,櫻屬植物是個(gè)比較年輕的屬,屬間物種大多可以彼此雜交,根據(jù)種的定義,嚴(yán)格而言都是亞種。
4 結(jié)論
本研究利用ITS和trnH-psbA對(duì)88個(gè)櫻花品種進(jìn)行了系統(tǒng)發(fā)育關(guān)系研究,結(jié)果表明,山櫻群櫻花品種系統(tǒng)發(fā)育比較混亂,這與其遺傳背景比較復(fù)雜以及原始親本系統(tǒng)發(fā)育關(guān)系較近有關(guān),江戶(hù)彼岸群和寒緋櫻群的大多數(shù)櫻花品種能夠聚類(lèi)在一起,與山櫻群品種能分得開(kāi),但涉及到很多雜交品種,ITS 和trnH-psbA 難以區(qū)分,表明DNA 條形碼技術(shù)難以應(yīng)用到雜交品種的系統(tǒng)發(fā)育關(guān)系,因此,對(duì)于櫻花的雜交品種可采用SSR、ISSR 等分子標(biāo)記技術(shù),探討彼此間親緣關(guān)系。本研究結(jié)果可作為櫻花品種分類(lèi)、嫁接繁殖、系統(tǒng)發(fā)育等研究的參考依據(jù)。
