蘭州百合不同層次鱗片無介質催培效果的差異
2020-03-08 12:53:00曹彩霞王娟唐楠唐道城劉高峰巨秀婷
江蘇農業科學 2020年21期
曹彩霞 王娟 唐楠 唐道城 劉高峰 巨秀婷
摘要:為提高蘭州百合的繁殖系數,解決種鱗莖供應量不足問題,以及為蘭州百合小鱗莖工廠化繁殖提供理論依據和技術參考。選擇蘭州百合不同層次鱗片為試驗材料,利用無介質催培方法進行小鱗莖繁殖,分別從鱗片催培效果(疑似發病率、分化率)以及小鱗莖催培效果(分化數、生根率、質量)進行測定,并對分化得到的小鱗莖按鱗片層次進行分級,以期明確蘭州百合不同層次鱗片在小鱗莖繁殖過程中的差異,擬篩選催培效果最佳鱗片的層次。在催培整個周期,蘭州百合鱗片的催培效果在不同層次間均存在差異。催培初期,外、中、內3層鱗片均有不同程度的疑似發病情況,并在21 d時達到高峰,28 d時疑似發病率逐漸下降并穩定;此時3層鱗片均有不同程度的小鱗莖分化。催培結束后,外、中、內3層鱗片的分化率分別達到82.67%、93.00%、93.33%,每層鱗片分化小鱗莖數分別為192.33、213.67、194.67粒/100片,生根率分別達到59.00%、74.33%、79.67%,外、中、內3層鱗片產生小鱗莖(100粒)的質量分別為109.55、87.20、60.93 g。綜合分析,整個催培周期中14~21 d是小鱗莖數量快速增長的時期,經過49 d的無介質催培發現,中層鱗片各項指標綜合表現最好,分化率達到93.00%,小鱗莖分化數(100片鱗片)達到213.67粒,分化1~4粒小鱗莖的鱗片數約占總數的91%以上。3層鱗片的繁殖系數從高到低依次為中層>內層>外層。
關鍵詞:蘭州百合;無介質催培;小鱗莖;快繁;分級;繁殖系數
中圖分類號:S644.104+.3?文獻標志碼:A?文章編號:1002-1302(2020)21-0181-04
蘭州百合[Lilium davidii var. unicolor (Hoog) Cotton]是百合科(Liliaceae)百合屬(Lilium)多年生球根作物,屬單子葉草本植物,其鱗莖瓣厚、豐潤、口感甜美,具有很高的營養及經濟價值[1],是我國的最佳食用百合品種[2],深受消費者青睞,市場需求量極大。蘭州百合生長周期長,且對種植區域的海拔、氣候、土壤均有特殊要求,使得蘭州百合在市場上常常出現供不應求的局面,因此選擇適宜的繁殖方法可以解決種球供應量問題。目前鱗片繁殖法常見的有苗床扦插法、室內砂培法、鱗片氣培法。蘭州百合的鱗片氣培法最早由高彥儀等提出,該方法利用鱗片剝傷處維管束的薄壁細胞在刺激下恢復細胞分生能力,使鱗片剝傷處生根發芽萌發小鱗莖[3-4]。何舉忠等認為氣培法較基質扦插法有很多優點,如產率高、生長周期短、單位面積再生籽球數量多、適宜工廠化生產等[5]。蘭州百合最常用且繁殖系數最高的材料是鱗片[6],小鱗莖由鱗片下部的組織細胞形成[7],且不同鱗瓣部位對小鱗莖繁殖具有一定的影響[8]。目前關于蘭州百合鱗片催培效果的報道相對較早,且針對不同層次鱗片催培方式的研究鮮見報道。為了提高蘭州百合的繁殖系數,解決種鱗莖供應量,本研究擬在蘭州百合的不同鱗片層次之間進行無介質催培效果篩選試驗,利用鱗片催培效果(疑似發病率、分化率)以及小鱗莖催培效果(分化數、生根率、小鱗莖質量),以期獲得相同的無介質催培條件下各鱗片之間的催培效果差異,篩選最佳的無介質催培鱗片層次。
1 材料與方法
1.1 試驗材料
選取色澤潔白、品相良好且飽滿的蘭州百合,先將外層腐爛有破損的鱗片剝除,然后依次從外向內取1~2層設為外層鱗片,3~4層設為中層鱗片,5~7層設為內層鱗片。鱗片沖洗干凈并浸泡消毒(50%甲基硫菌靈100倍液)30 min,清水沖洗并瀝干備用;催培盤用84消毒液浸泡消毒 10 min,40%甲醛溶液消毒表面,瀝干備用。
1.2 試驗設計
試驗于2018年4月在青海省園林植物與觀賞園藝重點實驗室的氣培室進行,分別將蘭州百合外、中、內3層鱗片置于催培盤中,每盤放置1 kg鱗片(外層約380片、中層約580片、內層約860片)進行催培試驗,每個鱗片層次設3個重復。催培室溫度白天控制在25 ℃,夜間控制在10 ℃,濕度控制在60%~80%。共催培49 d。
1.3 測定項目及方法
催培開始后分別對鱗片疑似發病率、分化率以及小鱗莖分化數、生根率進行測定,每隔7 d每個處理隨機取100片鱗片觀察并計數,共記錄49 d。催培結束后每個處理隨機選取30片鱗片并分離小鱗莖進行質量檢測,按照小鱗莖直徑分為一級(直徑≥7 mm)、二級(4 mm≤直徑<7 mm)、三級(直徑<4 mm)。
1.4 統計分析
試驗數據整理利用Micosoft Excel 2010軟件,數據統計分析利用SPSS 17.0版軟件。
2 結果與分析
2.1 蘭州百合各層次鱗片的催培效果分析
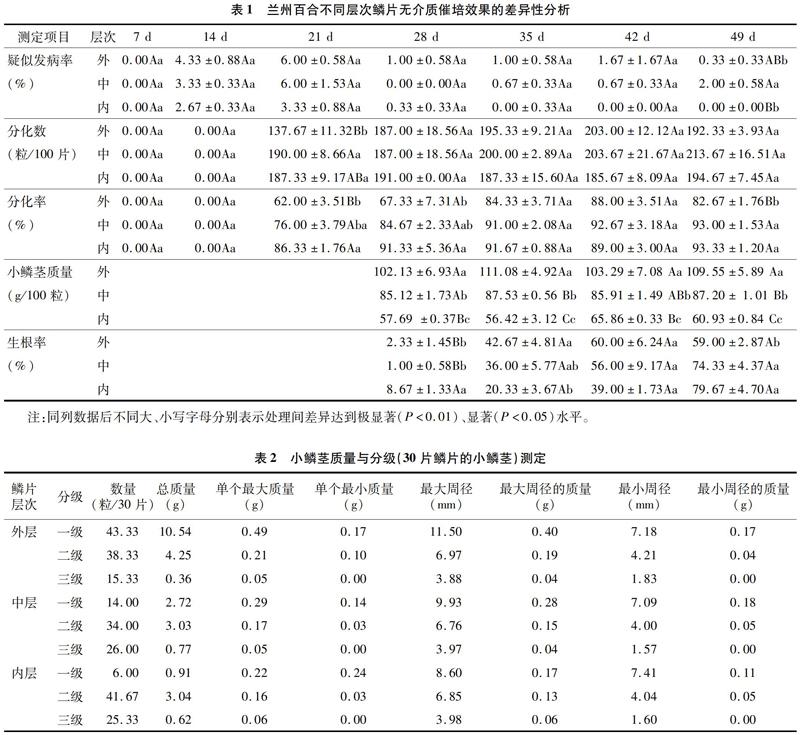
由表1可以看出,蘭州百合的不同層次鱗片在相同催培條件下的鱗片疑似發病率不同,隨著催培時間的延長,鱗片疑似發病率總體呈先上升后下降的變化趨勢。0~21 d,各層次鱗片的疑似發病率快速上升,21 d時達到最大值,且鱗片層次間差異不顯著;21 d后各層次鱗片愈傷開始形成,部分疑似病狀逐漸消失,疑似發病率逐漸下降,到催培結束(49 d)時,各層次鱗片的疑似發病率均低于3.00%。
催培過程中,鱗片基部出現白色小凸點即鱗片開始分化小鱗莖,催培14 d后3層鱗片均開始分化,內層鱗片在21 d時就達到分化高峰,中層和外層鱗片在35 d時才達到分化高峰,高峰期之后,各層次鱗片均保持相對穩定。說明在無介質催培中鱗片在35 d內完成全部分化。催培結束時,中層、內層鱗片的小鱗莖分化率極顯著高于外層,均達到93.0%左右。
2.2 蘭州百合各層次鱗片產生小鱗莖的質量檢測
催培結束后,3層鱗片的小鱗莖分級情況詳見表2。外層鱗片形成的一級小鱗莖數量最多,達43.33粒/30片,內層鱗片的一級小鱗莖最少(6.00粒/30片),表現為與催培鱗片的質量和體積相關,鱗片從外向內一級小鱗莖數逐漸減少;以內層鱗片形成的二級小鱗莖數最多(41.67粒/30片),中層鱗片的最少(34.00粒/30片);各層次鱗片的三級小鱗莖,以中層鱗片最多,達26.00粒/30片;外層鱗片最少,僅15.33粒/30片。
外層鱗片的一級小鱗莖質量占總質量的69.57%,二級小鱗莖質量占28.05%,三級小鱗莖質量僅占2.38%;中層鱗片的一級小鱗莖質量占總質量的41.72%,二級小鱗莖質量占總質量的46.47%,三級小鱗莖質量占總質量的11.81%;內層鱗片的一級小鱗莖質量占總質量的19.91%,二級小鱗莖質量占總質量的66.52%,三級小鱗莖質量占總質量13.57%。由此可以看出,鱗片由外向內,分化的小鱗莖盡管在數量上差異不明顯,但分化形成的小鱗莖質量差異明顯,一級小鱗莖的質量急劇下降,二級小鱗莖和三級小鱗莖的質量在大幅增加。
3 討論與結論
3.1 鱗片疑似發病率對催培效果的影響
在整個催培周期中不同層次的蘭州百合鱗片疑似發病情況各不相同,鱗片疑似發病率0~49 d的總和由高到低依次是外層>中層>內層。此現象可能是因為外層鱗片處于種球的外層,最容易受到破損和病菌感染,在催培開始前,鱗片內部已存在部分病原菌[9-10],同時外層鱗片的淀粉含量高,營養物質較多[11],這些都是造成外層鱗片疑似發病率較高的原因;內層鱗片因處于快速生長時期,能抵抗外界的病原菌,且無機械損傷,故其疑似發病率較低。鱗片疑似發病率隨著催培時間的延長出現變化,各層鱗片的疑似發病率均在21 d時出現高峰,原因可能是在進行蘭州百合鱗片剝離時產生不可避免的傷口,在催培初期鱗片傷口未及時愈合暴露在空氣中,容易感染病原菌從而使鱗片出現疑似發病現象,同時為保持濕度采用的鱗片浸水方式也會使病原菌更易擴散。催培期間,鱗片基部未出現疑似發病的組織細胞形成小鱗莖,當小鱗莖逐漸膨大時,鱗莖生長周圍疑似發病的細胞組織漸漸愈合,鱗片疑似發病率逐漸降低。
3.2 不同鱗片層次之間的小鱗莖分化差異
蘭州百合不同層次鱗片的分化率總體為內層鱗片最好,中層鱗片次之,外層鱗片最差;小鱗莖分化數總體為中層鱗片最高,內層鱗片次之,外層鱗片最低。綜合比較得到,蘭州百合鱗片繁殖系數從高到低依次為中層>內層>外層。前人研究認為,不同層次鱗片的小鱗莖分化率和分化數存在差異與鱗片處于不同的休眠狀態[12]、鱗片所含的營養狀況、相關酶活性及含量相關。因內層鱗片位于鱗莖內側,鱗片發育較外層和中層不完全,營養物質基礎相對外、中層較差,導致其分化小鱗莖個數較少;外層鱗片的疑似發病率較高、遭受更多病原菌侵染的同時影響了外層鱗片的分化能力[13],使得外層鱗片的分化率略低于中層鱗片。蘭州百合的鱗莖是營養物質的儲藏器官,也是進行無性繁殖的器官[14],選擇質厚、儲存營養物質豐富的鱗片,更利于形成小鱗莖。隨著小鱗莖的逐漸增長,鱗片逐漸干萎,其細胞體積也相應減小[15],導致蘭州百合不同層次的鱗片產生小鱗莖的能力不同。
3.3 催培效果對小鱗莖質量與分級的影響
催培結束(49 d)時,不同層次蘭州百合鱗片生根率由高到低依次是內層>中層>外層,根是小鱗莖和葉、芽形成后能否繼續生長的關鍵[16];不同層次的蘭州百合鱗片產生一級小鱗莖最多的鱗片層次依次是外層>中層>內層;二級小鱗莖的鱗片層次從高到低依次是內層>外層>中層;三級小鱗莖的鱗片層次從高到低依次是中層>內層>外層;催培期間,隨著鱗片上鱗莖的分化、數量的增多及體積的擴大[17],不同層次鱗片的厚度及營養物質的存儲量與其形成小鱗莖的大小和質量相關。周生壇等研究發現,百合形成小子球的能力與單鱗片在母球上所處的位置有較密切的關系,不同部位鱗片所含養分不同,不同層次鱗片營養物質的存儲量不同,導致各層次鱗片產生小鱗莖的質量不同[18-19]。孫紅梅等研究發現,外層和中層鱗片繁殖能力較強,內層鱗片細胞再生能力強,但由于基部面積較小,營養積累少,不能為分化提供充足的能量,因此繁殖系數較低[20]。
綜合分析,整個催培周期中14~21 d是小鱗莖數量快速增長的時期,蘭州百合經過49 d的鱗片無介質催培后,中層鱗片各項指標綜合表現最好,分化率達到93.00%,小鱗莖分化數(100片鱗片)達到213.67粒,分化1~4粒小鱗莖的鱗片數約占91%,3層鱗片的繁殖系數從高到低依次為中層>內層>外層。
參考文獻:
[1]張紅巖,周 興,莫勇生,等. 蘭州百合組織培養及快速繁殖技術研究[J]. 廣西科學院學報,2015,31(1):49-53.
[2]劉 靜. 蘭州百合快速繁殖研究[J]. 南方農業學報,2011,42(8):839-842.
[3]高彥儀,張金娣,劉德義,等. 百合“氣培法”生產籽球新技術的研究總結[J]. 甘肅農業科技,1988(1):19-21,23.
[4]馬君義,趙小亮,張 繼,等. 蘭州百合的研究進展[J]. 塔里木大學學報,2005,17(4):53-56.
[5]何舉忠,李云亞,徐學民. 百合鱗瓣氣培法工廠化生產籽球技術開發研究[J]. 甘肅農業科技,2002(2):29-30.
[6]鄭愛珍,張 峰. 百合的繁殖方法[J]. 北方園藝,2004(4):43.
[7]鄭 鑫. 亞洲百合無性繁殖小鱗莖技術的研究[D]. 哈爾濱:東北農業大學,2017.
[8]李云亞,張 丹,趙 潔,等. 蘭州百合鱗瓣氣培繁育小鱗莖技術研究[J]. 中國農學通報,2014,30(16):248-251.
[9]田麗麗. 甘肅省切花百合病害及種球包衣劑的研究[D]. 蘭州:甘肅農業大學,2008.
[10]胡新穎,楊迎東,顏津寧,等. 百合鱗片全基質包埋試驗[J]. 江蘇農業科學,2014,42(2):147-149.
[11]劉仁坤. “索邦”百合鱗片扦插繁殖技術研究[J]. 安徽農業科學,2010,38(31):17419-17420.
[12]王愛勤,何龍飛,盛玉萍,等. 百合鱗片不同處理與鱗莖形成關系的研究[J]. 廣西農業生物科學,2003,22(3):182-185.
[13]毛軍需,梁建國,孫福慶,等. 百合鱗片扦插繁殖研究[J]. 安徽農業科學,2007,35(31):9884-9885,9887.
[14]陳麗靜,馬 爽,李麗麗,等. 東方百合“索蚌”離體培養快繁體系建立[J]. 西南農業學報,2010,23(5):1652-1655.
[15]韋梅琴,唐 蓉,沈寧東,等. 觀賞百合氣培過程中鱗片的細胞形態學觀察[J]. 青海大學學報(自然科學版),1998,16(3):8-11.
[16]桑 林,林衛東,謝慶華. 激素對百合鱗片扦插繁殖的影響研究[J]. 西南農業學報,2006,19(3):473-475.
[17]唐 蓉,沈寧東,韋梅琴,等. 觀賞百合鱗片氣培過程中器官分化[J]. 北方園藝,1998(3):96-98.
[18]周生壇,劉世安. 東方百合鱗片催培繁殖研究[J]. 北方園藝,2013(3):85-88.
[19]單 艷,李枝林,趙 輝. 百合鱗片扦插繁殖技術研究綜述[J]. 中國農學通報,2006,22 (8):365-368.
[20]孫紅梅,賈子坤,陸 陽,等. 百合鱗片扦插繁殖的研究進展[J]. 北方園藝,2009(2):141-146.薛晨晨,寶雨欣,繆亞梅,等. 早春鮮食大豆設施栽培適宜播種期[J]. 江蘇農業科學,2020,48(21):185-187.
