克氏原螯蝦生長的損傷性補償效應
2020-03-08 12:53:00徐韻苓陳友明王山黃成黃亞紅
江蘇農業科學 2020年21期
徐韻苓 陳友明 王山 黃成 黃亞紅

摘要:為研究斷螯對克氏原螯蝦生長的影響,以亞成體克氏原螯蝦為試驗材料,研究了克氏原螯蝦失去大螯后的個體存活率與大螯再生、體質量、攝食量和餌料轉化效率等的生長狀態變化。結果表明,失去大螯后的亞成體螯蝦在單養條件下存活率與全肢螯蝦無顯著性差異(P>0.05),但在非單養條件下存活率顯著大于全肢螯蝦(P<0.05);螯蝦在斷螯后有近80%的個體可實現大螯再生,且攝食量、體質量增長率極顯著大于全肢螯蝦(P<0.01);雌性斷螯螯蝦餌料轉化效率顯著大于全肢螯蝦(P<0.05)。綜上所述,克氏原螯蝦斷螯后攝食量較多、生長較快,且雌性斷螯螯蝦餌料利用效率比全肢螯蝦高,同時斷螯的發生可以有效降低螯蝦打斗的死亡率。本研究從生理學和行為學角度對螯蝦斷螯后的損傷性補償生長進行了討論分析,以期對克氏原螯蝦的基礎理論研究和生產實踐提供參考。
關鍵詞:克氏原螯蝦;斷螯再生;補償效應;攝食量;體質量增長
中圖分類號:S966.12?文獻標志碼:A?文章編號:1002-1302(2020)21-0207-04
克氏原螯蝦(Procambarus clarkii)屬甲殼綱、十足目、螯蝦科,因其具有個體大、生長快、肉質鮮美和營養豐富等特點[1],已成為養殖戶和消費者極其青睞的特色水產品種。但是克氏原螯蝦性情兇猛、生性好斗,且種內斗爭較為常見[2],在養殖過程中經常出現因相互打斗導致大螯缺失甚至個體死亡等現象。本研究發現一個有趣的情況,在大多數的養殖生產中,無論是選擇幼蝦還是選擇繁育后代的親蝦,蝦農普遍認為肢體完整的個體更好;在理論研究中,無論是關于克氏原螯蝦繁殖行為的研究,比如張麗等研究的婚配后雄性螯蝦的擇偶行為,或是關于社會環境與行為方面的研究[3],如王陳路等研究的不同地位螯蝦對社會環境改變的行為反應,又或是關于螯蝦的食性研究[4],如陳翔宇等研究的食性對螯蝦格斗行為的影響[5],在其選擇實驗動物時都著重強調選用的是螯肢完整的個體,認為斷螯個體是不健康的或處于弊端的劣勢個體。但在實際生產中有人認為螯肢完整與否并無影響,也有人認為螯肢殘缺個體反而更好,那么克氏原螯蝦斷螯后,個體生長實際情況到底如何呢?
國內外關于生物體斷肢現象的研究,如在節肢動物門中蛛形綱的蜘蛛類和甲殼綱的蟹類的斷肢研究中發現,斷肢后機體發育會延長[6];招潮蟹(Uca arcuata)能通過延長下一次的蛻殼期而使斷肢在蛻殼時重新長出[7];譚梁飛等發現,中華真地鱉(Eupolyphaga sinensis)斷肢會延長蛻皮間期時間,且斷肢數量越多、斷肢期日齡越大,間期時間越長,說明斷肢會導致生長周期變長,從而保證足夠的時間促進機體修復[8];斷肢也會延緩甲殼類蛻殼,以便促進斷肢恢復,如在甲殼綱十足目的種類中就發現斷肢會延長蛻殼間期,使斷肢再生后再進行蛻皮[9];鋸緣青蟹(Scylla paramamosain)、陸地蟹等在發生肢體缺失現象后,其恢復會受到溫度、光周期和動物發育狀況等因素影響[10-11]。
目前,關于克氏原螯蝦斷螯后機體變化等的具體研究未見報道,本研究以亞成體克氏原螯蝦為試驗材料,通過去螯操作,研究斷螯對螯蝦生長狀況等的影響,不僅可以豐富克氏原螯蝦基礎生物學研究,同時對克氏原螯蝦養殖業也有一定的指導作用。
1 材料與方法
1.1 材料
本試驗于2018年5—9月在江蘇省淡水水產研究所揚中基地進行。試驗用蝦來源于江蘇省淡水水產研究所揚中基地,試驗前將蝦置于基地內水泥池暫養3 d,待蝦生長穩定后再進行試驗。試驗時,從同一批螯蝦中選出一批無病無傷、附肢完整、活力強的亞成體螯蝦,平均體質量為(3.41±0.68) g,根據試驗需要分別置于規格為48 cm×27 cm×28 cm 的長方體塑料箱和上口直徑 17.5 cm、下底直徑13.0 cm、高10.0 cm的圓臺形塑料容器中進行飼養觀察,飼養期間投喂嘉吉牌河蟹飼料(SC/T 1078—2004)。
1.2 方法
1.2.1 試驗設計 試驗分為2個部分,第一部分為單養試驗,每個碗狀容器飼養1尾克氏原螯蝦。將試驗用蝦進行編號、稱量體質量,隨機分為2組:試驗組、對照組各60尾(雌雄各半)。第二部分為非單養試驗,每個長方體容器放置2尾性別相同的螯蝦。將試驗用蝦進行編號、稱量體質量,隨機分為2組:試驗組、對照組各40尾(雌雄各半)。
試驗組克氏原螯蝦的大螯用人工壓力法去掉大螯后再次稱質量記錄體質量。所有螯蝦分別放置在各組相應的實驗容器中飼養。每天定時投喂飼料并在2 d投喂前記錄死亡數、攝食量后,換掉 1/2 的水,清理殘余飼料同時觀察殘肢的生長恢復情況。試驗期間保持各個容器的養殖環境基本一致。單養試驗進行45 d,非單養條件試驗進行30 d。
1.2.2 指標測定與計算 體質量增長率、餌料轉化效率和死亡率計算公式如下:
(3)死亡率的百分比檢驗公式為:
式中:m0、mt分別為試驗開始時和結束時的螯蝦體質量,t為試驗天數(d);F為攝食總量;a為試驗組死亡數;A為試驗組樣本容量;b為對照組死亡數;B為對照組樣本容量。百分比檢驗結果,u>1.96,P<0.05(表示差異顯著);u>2.58,P<0.01(表示差異極顯著)。
1.3 數據處理
試驗結果采用“平均數±標準差(x±s)”表示,利用Excel 2013、SPSS 25.0軟件進行獨立樣本t檢驗分析,顯著性水平設為0.05。另外還采用百分比檢驗分析。
2 結果與分析
2.1 單養分析
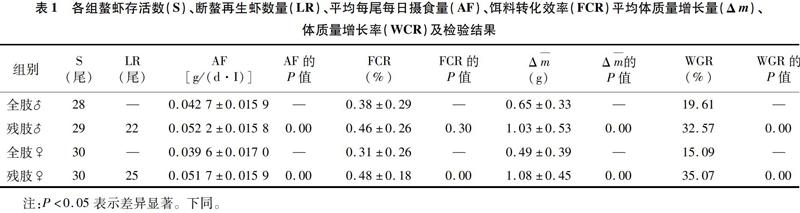
螯蝦存活尾數(S,Survival)、斷螯再生尾數(LR,Limb Regeneration)、平均攝食量[AF,Average Food Intake(g/(d·I))]、餌料轉化效率(FCR,Food Conversion Ratio)和平均體質量變化(Δm)、體質量增長率[WGR,Weight Growth Rate(%)]等情況及檢驗結果見表1。
2.1.1 存活與再生 由表1可知,試驗過程中雄性全肢螯蝦存活28尾,雄性殘肢螯蝦存活29尾;雌性全肢及殘肢螯蝦均無死亡,這表明斷螯對螯蝦死亡率并無影響。亞成體殘肢螯蝦最終存活59尾,螯肢再生的共有47尾,占存活數量的比例為79.67%。
2.1.2 攝食量分析 由表1可知,殘肢螯蝦攝食量大于全肢螯蝦。比較試驗組和對照組間的平均攝食量情況,雙側檢驗結果表明,亞成體克氏原螯蝦殘肢螯蝦攝食量極顯著大于全肢螯蝦(P<0.01),說明亞成體螯蝦在斷螯后攝食量較全肢螯蝦多。
2.1.3 餌料轉化效率 由表1可知,殘肢螯蝦餌料轉化效率高。雙側檢驗結果表明,雄性殘肢螯蝦餌料轉化效率與雄性全肢無顯著差異(P=0.30>0.05),雌性殘肢螯蝦餌料轉化效率顯著高于雌性全肢螯蝦(P=0.00<0.05)。綜上所述,斷螯會提高雌性克氏原螯蝦的餌料轉化效率。
2.1.4 體質量增長量、體質量增長率 由表1可知,殘肢螯蝦的平均體質量增長量和體質量增長率皆比全肢螯蝦大。比較試驗組和對照組間體質量情況,雙側檢驗結果表明,殘肢螯蝦體質量增長量極顯著大于對照組全肢螯蝦(P<0.01)。考慮到全肢螯蝦最終體質量包含大螯質量而殘肢螯蝦沒有,所以以體質量增長率為標準衡量體質量變化情況更為合理,雙側檢驗結果表明,殘肢螯蝦體質量增長率極顯著大于對照組全肢螯蝦(P<0.01),表明斷螯促進了克氏原螯蝦體質量的增加,試驗組殘肢螯蝦出現了超補償生長。
2.2 非單養條件下存活試驗
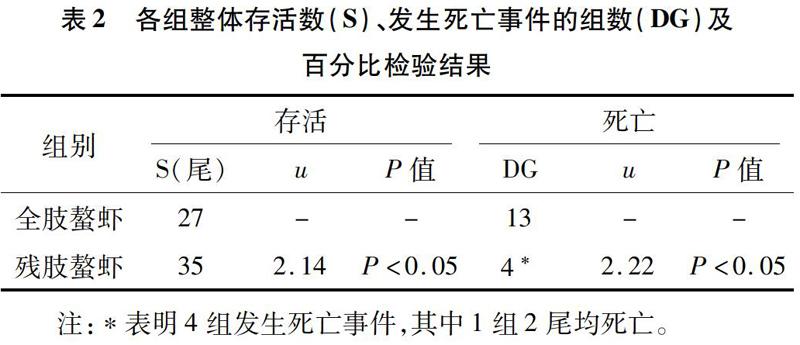
在非單養條件下,為探究斷螯對克氏原螯蝦的影響,采取2尾螯蝦共養模式,觀察殘肢螯蝦、全肢螯蝦在社會社區壓力和斷螯損傷雙重壓力的應激狀態下的存活率。試驗以發生死亡的組數和死亡個體數量作為衡量指標,通過百分比檢驗比較顯著性差異。因為螯蝦生性好斗,在非單養條件下存在同類相殘相食現象,所以無法準確測定螯蝦的攝食量,非單養試驗將不再探究螯蝦的攝食量和體質量變化情況。試驗各組存活螯蝦尾數(S)、發生死亡組數(DG,Died Group)及相應百分比檢驗結果見表2。
2.2.1 死亡率統計與檢驗 由表2可知,試驗組螯蝦有4組出現死亡事件,對照組有13組出現死亡事件,對其進行百分比檢驗,u=2.22>1.96,P<0.05,即有顯著性差異,說明試驗組發生死亡事件的概率顯著小于對照組。且死亡13尾克氏原螯蝦中有6尾打斗致殘,說明是經過激烈打斗致死的,統計個體死亡情況,對非單養條件下試驗的死亡率數據進行百分比檢驗,殘肢螯蝦與全肢螯蝦死亡率百分比檢驗結果為u=2.14>1.96,P<0.05,即有顯著性差異,說明非單養條件下殘肢螯蝦比正常螯蝦死亡率要低。
由表2可知,非單養條件下殘肢螯蝦及全肢螯蝦死亡率分別與表1中單養條件下殘肢螯蝦及全肢螯蝦死亡率進行百分比檢驗,結果分別為u=2.21>1.96(P<0.05)和u=3.98>2.58(P<0.01)(表2未顯示該數據)。這說明只是斷螯壓力不會對螯蝦存活產生應激刺激,生存不會受到影響(單養條件下全肢與殘肢螯蝦死亡率無顯著性差異);但非單養條件下在社會社區壓力和斷螯雙重壓力應激下,特別是社會競爭壓力的增大顯著,甚至極顯著(全肢螯蝦死亡率對比檢驗)提高了螯蝦的死亡率。
3 討論與展望
3.1 存活與再生
在單養試驗中,殘肢螯蝦與全肢螯蝦死亡率無顯著差異,這表明螯蝦并不會因斷螯而增加死亡率。在本研究和已有文獻對克氏原螯蝦探討中均發現,克氏原螯蝦存在自切現象[2],即螯蝦的大螯在外力脅迫下會從關節處自我切斷,這可能是螯蝦為防止血液外流、擴大傷口感染,避免因斷螯而導致死亡,這種現象是大多數節肢動物在漫長進化過程中形成的自我保護機制[11]。而在非單養條件下,殘肢組死亡率顯著低于全肢組,這可能是因為克氏原螯蝦性格兇猛,生性好斗[2],斷螯有效避免了螯蝦因為利用大螯相互打斗而導致死亡。非單養條件下螯蝦死亡率顯著大于單養條件,表明只有斷螯不會對螯蝦存活產生應激刺激,其生存不會受影響(單養條件下全肢與殘肢螯蝦死亡率無顯著性差異),但在非單養條件下社會社區壓力和斷螯雙重壓力應激特別是社會競爭壓力的增大,顯著甚至極顯著地提高了螯蝦死亡率。但在社會壓力下,斷螯的螯蝦存活率又顯著高于全肢螯蝦,說明斷螯提高了螯蝦社會競爭力,有效降低了螯蝦因為相互打斗而導致的死亡率,斷螯可有效提高存活率。
本研究中有近80%殘肢螯蝦重新長出大螯,但在中華絨螯蟹(Eriocheir sinensis)斷肢研究中殘肢蟹全部長出完整肢[12],作者猜測可能是因為中華絨螯蟹和克氏原螯蝦斷肢再生恢復機制有所不同,也可能是本次試驗周期不夠長,不足以讓全部螯蝦完成斷螯再生。
3.2 斷螯對攝食影響
本試驗中克氏原螯蝦雖然缺失了大螯,在一定程度上減少了運動的消耗量,但是通過螯蝦攝食量數據分析發現,大螯的缺失反而提高了殘肢螯蝦的攝食量。生物界不同生物在斷肢后會有不同的攝食策略來應對大螯損失的能量代價,如對多數昆蟲的研究中特別是普列熱蝽,在肢體發生殘缺后會大量進食,進入蛻皮期通過蛻皮來促進斷肢再生[13],本試驗結果與之類似。但在尾鰭缺失對3種鯉科魚類影響的研究中結果相反[14],3種鯉科魚類在尾鰭缺失后體內攝食代謝模式會減少,而把更多的能量分配到運動上,補償因為尾鰭缺失而造成的運動能力減弱這個現象。考慮到大螯是克氏原螯蝦主要的打斗器官,同時大螯的缺失也不會影響克氏原螯蝦的運動能力,所以在發生斷螯現象后,運動不會更多消耗體內能量。有的動物斷肢后攝食量不變,如麗斑麻蜥斷尾既不促進也不抑制該種蜥蜴攝食量[15],該試驗研究者認為在該種蜥蜴中補償斷尾能量代價對策不是增加攝食,猜測可能是以減少活動來節約體內能量損耗。但考慮到麗斑麻蜥屬于爬行綱,它們的斷肢再生是屬于直接生長出來,而不是像甲殼綱的蝦、蟹等,昆蟲綱的蜘蛛等以蛻皮促進生長,恢復斷肢。因此,本研究認為克氏原螯蝦在斷肢后恢復斷肢的對策基本上是通過增加攝食來實現補償生長。
3.3 體質量增長及餌料轉化效率
生命有機體的損傷性修復是一種生長過程。克氏原螯蝦在受到斷螯脅迫后,以大量攝食對策來彌補損失大螯的能量代價實現快速生長,這種機制與饑餓性損傷補償生長類似。補償生長分為超補償生長、完全補償生長、部分補償生長和不補償生長4種類型[16]。在克氏原螯蝦對多重周期饑餓研究中發現,克氏原螯蝦可以在脅迫環境下產生補償生長[17]。克氏原螯蝦在經歷斷螯脅迫后迅速適應,體質量增加、平均體質量增長量和體質量增長率較全肢螯蝦呈極顯著差異,殘肢螯蝦表現出超補償生長,這與對海星研究的結果類似,殘肢海星會極顯著增加體質量,以盡快恢復覓食能力,同時恢復攻擊的武器[18]。斷螯刺激克氏原螯蝦大量積累能量,外在表現就是體質量增加,然后將能量分配至損失的斷螯處,從而恢復斷螯,對狼蛛研究中也發現類似結果[19],同時狼蛛會減少其他方面能量消耗,集聚體內能量到下一個蛻皮期,以減輕體質量和減小體型為代價來恢復斷肢。
在本研究中雌性殘肢克氏原螯蝦的餌料轉化效率顯著大于全肢螯蝦組,說明雌性殘肢螯蝦對飼料的利用率和轉化率提高了。在攝食量相同情況下,雌性斷螯螯蝦的體質量增加量較多。筆者猜測可能是亞成體雌性螯蝦為了即將到來的生殖活動,所以要更加高效地儲存能量同時還要促進性腺的進一步發育。當生物體遭遇外力脅迫特別是不可抗的強大因素時都會選擇在最后將體內能量更多分配到生殖活動上,從而繁衍后代。如淡水蟹在斷肢刺激后會促進卵巢的發育[20],說明在遭遇外界不良環境時會將更多的能量傾向于生殖活動上。本試驗中則體現在雌性螯蝦斷螯后的體質量超補償性增加。
3.4 展望
亞成體克氏原螯蝦對斷螯產生的機體應激機制還不完全清楚,下一步還可以探究體內哪些激素分泌和相關基因表達以及微小核酸作用于攝食量、體質量等,從而揭示其內部變化。成體克氏原螯蝦的生長雖然已經定型,但是在受到斷螯損傷脅迫后機體是否會產生相應應激狀態還未可知,也可探究成年螯蝦在斷螯后的生理生化狀態。
參考文獻:
[1]何金星,竇 寅,唐建清,等. 飼料添加螺旋藻對克氏原螯蝦生長性能的影響[J]. 江蘇農業科學,2009(2):209-211,250.
[2]王 慶. 克氏原螯蝦繁育機制及養殖生態學研究[D]. 南京:南京師范大學,2012:2-44.
[3]張 麗,謝 春,唐建清,等. 婚配后雄性克氏原螯蝦的擇偶行為[J]. 南京大學學報(自然科學版),2014,50(6):890-894.
[4]王陳路,曾小翠,沈佳琰,等. 不同地位克氏原螯蝦對社會環境改變的行為反應[J]. 動物學雜志,2015,50(4):555-562.
[5]陳翔宇,王陳路,黃 成,等. 食性對克氏原螯蝦格斗行為的影響[J]. 飼料工業,2014(增刊1):117-120.
[6]González M,Barría E. Effect of autotomy and regeneration of the chelipeds on growth and development in Petrolisthes laevigatus (Guérin,1835) (Decapoda,Anomura,Porcellanidae)[J]. Crustaceana,2008,81(6):641-652.
[7]Holland C A,Skinner D M. Interactions between molting and regeneration in the land crab[J]. The Biological Bulletin,1976,150(2):222-240.
[8]譚梁飛,朱 芬,熊 強,等. 中華真地鱉斷足再生對發育的影響[J]. 昆蟲知識,2007,44(1):101-104.
[9]Donald L M. Interactions between limb regeneration and molting in decapod crustaceans[J]. American Zoologist,2001,41(3):399-406.
[10]Jie Gong,Yu Kun,Ling Shu,et al. Evaluating the effects of temperature,salinity,starvation and autotomy on molting success,molting interval and expression of ecdysone receptor in early juvenile mud crabs,Scylla paramamosain[J]. Journal of Experimental Marine Biology and Ecology,2015,464:11-17.
[11]Mattoni C I,García-Hernández S,Botero-Trujillo R,et al. Scorpion sheds ‘tail to escape:Consequences and implications of autotomy in scorpions (Buthidae:Ananteris)[J]. PLoS One,2015,10(1):e0116639.
[12]岳武成,陳 嬌,慈元吉,等. 斷肢再生對中華絨螯蟹蛻殼、生長及相關基因表達的影響[J]. 浙江大學學報(農業與生命科學版),2016,42(4):502-508.
[13]王孟卿,彩萬志. 昆蟲肢體再生的研究進展[J]. 昆蟲知識,2004,41(2):127-131.
[14]付 成,曹振東,付世建. 尾鰭缺失和再生對三種鯉科魚類游泳運動能力和攝食與運動代謝交互作用的影響[C]//中國海洋湖沼學會魚類學分會、中國動物學會魚類學分會2012年學術研討會.蘭州,2012.
