鎘脅迫對瘤突蒼耳、蒼耳及其雜合體種子萌發與幼苗生長的影響
2020-03-13 03:16:08薛晨陽高英美
生態學報 2020年1期
關鍵詞:植物
薛晨陽,高英美,曲 波,*
1 沈陽農業大學生物科學技術學院, 沈陽 110161 2 遼寧省全球變化與生物入侵重點實驗室, 沈陽 110161
污水灌溉、金屬開采、冶煉及加工是造成我國土壤重金屬污染的最大原因[1]。日益增加的土壤重金屬污染不僅嚴重影響了作物的產量和質量,而且通過食物鏈和食物網向上傳遞,影響了動物和人類健康[2]。對受重金屬污染的土壤進行修復迫在眉睫,植物修復技術是指利用植物吸收、降解、揮發、過濾、固定等作用來凈化土壤、水體中的重金屬元素,其具有成本低,操作簡單,對土壤擾動低,無二次污染等優點[3- 4]。目前在實際應用中已經取得一定的成效。比如Ji等在沈陽張士污灌區使用龍葵(Solanumnigrum)進行了為期兩年的原位植物修復試驗,發現地表20 cm土層中的重金屬鎘濃度由最初的2.75 mg/kg降低到1.53 mg/kg,降幅達到24.9%[5]。
生物量大、生長迅速、適應性廣且能夠富集或超富集一種或多種重金屬的植物是植物修復技術的關鍵[6]。目前發現的一些重金屬富集植物一般都存在一定的缺點,不利于大規模推廣應用。例如楊肖娥等發現東南景天(Sedumalfredii)對土壤中高含量的Zn有很強的忍耐、吸收和積累能力,當土壤中Zn含量為2269—3858 mg/kg,植株地上部分Zn含量達到4065—5000 mg/kg,富集系數為1.25—1.94[7],但東南景天在寒冷高海拔地區不開花,生長緩慢。同樣,Visoottiviseth等發現蜈蚣草(Pterisvittata)對土壤中高含量的As具有很強的吸收能力,當土As壤濃度為810—1400 mg/kg,蜈蚣草羽片As含量達到4240—6030 mg/kg,富集系數達到4.31—5.23[8],但蜈蚣草在淮河以北及沈陽地區不能生長。
現有的超富集植物仍難以滿足我們的實踐需求,超富集植物種質資源庫仍需要持續擴充。雜交是自然界中廣泛存在的基因交流現象,也是植物育種中常用的技術,其涉及到完整染色體上大量基因的重組,這很容易引起某些生物性狀的改變,比如更快的生長速度、更大的體積以及生物量等特征,從而使植物適應復雜多變的環境,提高后代的適合度[9- 10]。Lexer等發現入侵種矛盾向日葵(Helianthusparadoxs)是由向日葵(H.annuus)和原野向日葵(H.petiolaris)經天然雜交產生,獲得了雙親均不具有的耐鹽性狀,因此能夠成功入侵到鹽堿性的濕地生境中[11];Lin等從四川兩個不同的氣候-生態區采集了龍葵進行雜交,在盆栽實驗中發現雜合體提取效率和生物量均高于親本,在田間實驗中,雜合體生物量高于親本且莖稈的鎘提取效率顯著增加[12]。
通過野外調查,我們發現沈陽張士污灌區同時存在瘤突蒼耳(Xanthiumstrumarium)和蒼耳(Xanthiumsibiricum)兩種植物,且兩種植物之間有可能存在雜交現象。Jeelani等[13]和胡小英等[14]研究結果分別顯示蒼耳和瘤突蒼耳對重金屬鎘的轉移系數均小于1,二者將大量的重金屬鎘聚集在根部。這種類型植物可以用來穩定土壤中重金屬,防治其遷移[15]。瘤突蒼耳原產北美洲,由于抗逆性強、生長快、生物量大的特點,在我國東北地區已成為危害較重的入侵植物之一。目前,對瘤突蒼耳和蒼耳雜合體的研究主要集中在種子萌發特性[16]、對養分響應[17- 18]方面,關于鎘脅迫下二者雜合體種子萌發及幼苗生長的研究尚未見報道。
種子是植物生長周期中特殊而重要的階段,其對個體繁殖、種群擴展和抵抗不良環境有著重要的意義,而種子萌發是將植物種群從幕后轉變為現實種群的關鍵環節[19];幼苗生長階段是植物生活史中對環境生態因子反應最為敏感的時期之一,植物在該階段的表現能初步反映其抵抗外界干擾的能力[20]。因此,本試驗通過比較蒼耳、瘤突蒼耳及其雜合體種子在不同鎘濃度處理下萌發及其幼苗生長情況,以期初步判斷雜合體對重金屬鎘的耐性是否優于親本,為超富集植物的篩選提供一種思路。
1 材料與方法
1.1 瘤突蒼耳、蒼耳及其雜合體種子蛋白質電泳實驗
1.1.1實驗材料
雜合體親本瘤突蒼耳和蒼耳種子分別于2012年11月采于遼寧省凌海市右衛滿族鎮小黃村(121°34′ E,41°04′ N)大凌河河岸沙地的自然成熟植株。2017年在沈陽農業大學教學科研基地分別進行瘤突蒼耳和蒼耳自交及雜交試驗,獲得瘤突蒼耳自交種子1000粒(采自5株植株),蒼耳自交種子1000粒(采自5株植株),雜合體(X.strumarium♀ ×X.sibiricum♂,簡稱雜交瘤突)種子591粒(采自5株植株),雜合體(X.sibiricum♀ ×X.strumarium♂,簡稱雜交蒼耳)種子935粒(采自5株植株)。用于收集自交種子的瘤突蒼耳和蒼耳在花期時選擇健康的花序進行套袋處理,防止受到干擾作為母本的瘤突蒼耳和蒼耳在花期時去除雄蕊,待柱頭呈羽毛狀分叉并帶有光澤時馬上分別授以蒼耳和瘤突蒼耳的花粉,授粉后的花序套袋隔離。待花期過后去除自交袋和雜交袋,待果實自然成熟后,按株采集,放置于通風避光處晾干,室溫保存備用。
野外疑似雜合體種子于2017年11月采于遼寧省朝陽市建平縣沙海鎮(119°27′ E,41°29′ N),按株采集,共收集55株。
1.1.2實驗方法
參考Osbome[21]對蛋白質的分類,將種子蛋白分為水溶蛋白(清蛋白)、鹽溶蛋白(球蛋白)、剩余蛋白(膠蛋白和谷蛋白)。取瘤突蒼耳、蒼耳自交及雜合體種子2—4粒(約0.15 g)。放于-20℃預冷的研缽,加入0.1 g SiO2,加入9 mL去離子水于冰盒上研磨,隨后轉入10 mL離心管,8000 r/min低溫離心4 min,取上清200 μL,移入1.5 mL 離心管,按照1∶1比例加入事先準備好的蛋白質提取液(0.15% SDS)200 μL,振蕩混勻,放入25℃光照培養箱中提取30 min后,配平,8000 r/min低溫離心4 min,取上清200 μL,移入1.5 mL離心管中4℃保存,作為水溶蛋白提取液;棄掉剩余上清液,加入9 mL去離子水,振蕩混勻,在25℃光照培養箱中提取30 min,配平,8000 r/min低溫離心4 min,去上清,再重復一次,去上清后,加入9 mL 1 mol/L NaCl溶液,振蕩混勻,放入25℃光照培養箱中提取30 min,配平,8000 r/min低溫離心4 min,取上清200μL,移入1.5 mL離心管中4℃保存,作為鹽溶蛋白提取液;棄掉剩余上清液,加9 mL 1mol/L NaCl溶液,振蕩混勻,在25℃光照培養箱中提取30 min,配平,8000 r/min低溫離心4 min,去上清,再重復一次,加入9 mL蛋白提取液放入25℃光照培養箱中提取30 min,配平,8000 r/min低溫離心4 min,取上清200 μL,移入1.5 mL離心管中4℃保存,作為剩余蛋白提取液。
采用10.2%醋酸尿素聚丙烯酰胺凝膠溶液制膠,2%冰乙酸為電極緩沖液(4℃保存),水溶、鹽溶、剩余蛋白點樣量分別為12、5、5 μL,接通電源,恒定100V下4小時。
水溶蛋白和剩余蛋白膠片染色12 h,鹽溶蛋白膠片染色5 h;水溶蛋白和剩余蛋白膠片加入脫色液(50%甲醇,7%的冰醋酸水溶液)洗脫1 min倒掉,重復3次,隨后加入脫色液,于恒溫25℃搖床 45 min;鹽溶蛋白膠片加入脫色液洗脫1 min,重復兩次,加入去離子水洗脫1 min,重復兩次,隨后加入去離子水,于恒溫25℃搖床 45 min。將膠片置于膠片觀察燈上并照相[22]。實驗結果進行4次重復。
野外疑似雜合體種子從55株中隨機選擇11株,試驗方法同上。
1.2 鎘脅迫對瘤突蒼耳、蒼耳及其雜合體種子萌發影響實驗
1.2.1實驗材料
選擇瘤突蒼耳、蒼耳自交種及雜合體種子各450粒(按株平均隨機選擇90粒),由于瘤突蒼耳和蒼耳種子具有二形性,相同條件下,上位種子休眠時間較下位種子長5—7 d[23]。為了消除上下位種子的影響,本實驗均選擇下位種子作為實驗材料。
1.2.2實驗方法
首先用5%的NaClO溶液浸泡種子10 min,去離子水沖洗5—6次,然后將消過毒的種子放入鋪有2層濾紙的培養皿中(直徑 9 cm,使用前均高溫高壓蒸汽120℃,30 min滅菌),每個培養皿放30粒種子。隨后將培養皿置于恒溫光照培養箱中,在25℃,12 h光照/12h 黑暗的條件下進行萌發。采用CdCl2·2.5H2O(分析純)設置0(CK)、10 、40、 80、120 mg/L 5個鎘濃度,每個處理3次重復。每個濃度均采用蒸餾水作為基礎液,對照組施以蒸餾水。為保持溶液濃度恒定,每隔24 h更換1次處理液和濾紙。
1.2.3指標測定
發芽勢=4 d內正常發芽種子數/供試種子數×100%
(1)
萌發率=7 d內正常發芽的種子數/供試種子數×100%
(2)
發芽指數 (GI)=∑(Gt/Dt)
(3)
式中,GI為在t日的發芽種子數;Dt為發芽天數[24]。
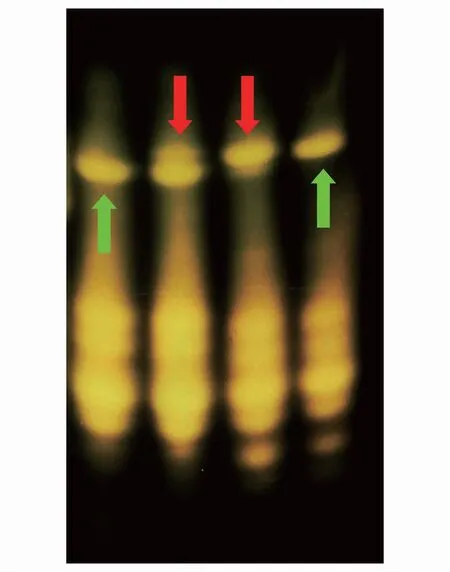
圖1 人工雜合體和親本種子鹽溶蛋白聚丙烯酰胺凝膠電泳圖 Fig.1 Salt-protein polyacrylamide gel electrophoresis diagram of artificial hybrids and parental seeds從左往右依次為瘤突蒼耳X. strumarium, 雜交瘤突X. strumarium♀ × X. sibiricum♂, 雜交蒼耳X. sibiricum♀ × X. strumarium♂, 蒼耳 X. sibiricum
1.3 鎘脅迫對瘤突蒼耳、蒼耳及其雜合體幼苗生長影響實驗
1.3.1實驗材料
待種子萌發實驗進行7 d后,將不同鎘處理和對照的種子萌發材料轉移至直徑15 cm培養皿,施以相同濃度的鎘處理液和蒸餾水,繼續培養3天,每隔24 h更換1次處理液和濾紙。
1.3.2實驗方法
第10天,用游標卡尺(精度0.01 mm)測量不同鎘濃度處理下瘤突蒼耳、蒼耳及其雜合體幼苗子葉長度和根長,隨后120℃殺青15 min,60℃烘箱中烘干后稱重,測定地上部分干重、地下部分干重并計算總干重;采用硫代巴比妥酸(TBA)比色法測定植物子葉丙二醛含量[25];采用愈創木酚法測定植物子葉愈創木酚過氧化物酶(GPX)活性[25];采用80%丙酮提取植物子葉葉綠素,并利用紫外光分光光度計測定470、646、663 nm波長下吸光度,根據Lichtenthaler和Wellburn[26]公式計算植物葉綠素含量。以上指標均隨機選擇3株幼苗。
1.4 數據處理
采用雙因素方差分析(two-way ANOVA)分析雜交、鎘濃度及二者交互作用對瘤突蒼耳、蒼耳種子萌發和幼苗生長的影響;隨后,在雙因素方差分析的基礎上進行多重比較(LSD)。采用SPSS 22.0進行統計分析,Origin 9.0作圖。
2 結果與分析
2.1 瘤突蒼耳、蒼耳及其雜合體與野外疑似雜合體種子蛋白質電泳
瘤突蒼耳與蒼耳鹽溶蛋白條帶存在顯著差異(圖1綠色箭頭),雜交瘤突和雜交蒼耳的水溶蛋白、剩余蛋白分別與瘤突蒼耳和蒼耳無明顯差異(未放入)。雜交瘤突和雜交蒼耳的鹽溶蛋白同時具有親本瘤突蒼耳和蒼耳獨有的蛋白條帶(圖1紅色箭頭),說明雜合體種子確實為瘤突蒼耳和蒼耳雜交而來。
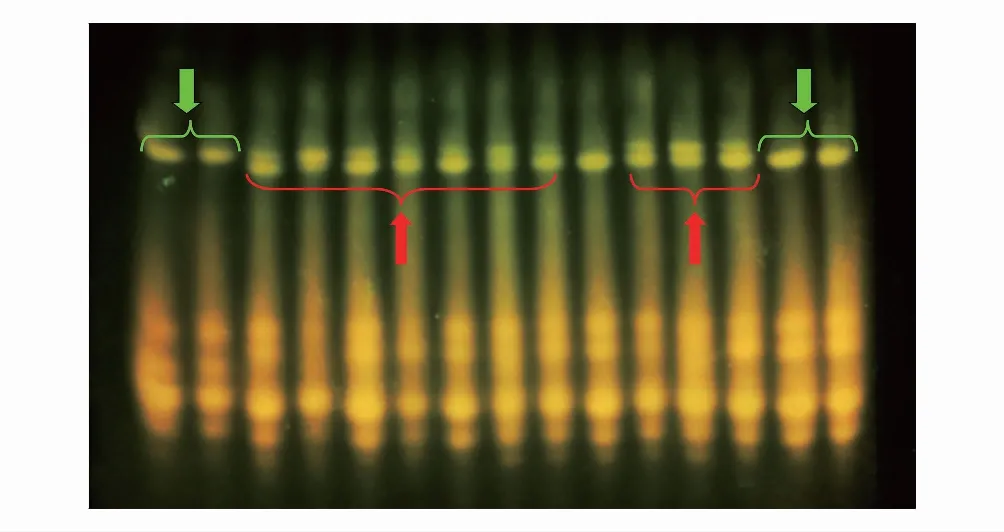
圖2 遼寧省朝陽市建平縣沙海鎮疑似雜合體種子鹽溶蛋白電泳圖Fig.2 Salt-soluble protein electrophoresis map of suspected hybrids in Shahai Town, Jianping County, Chaoyang City, Liaoning Province從左往右前2個條帶: 蒼耳; 中間條帶: 疑似雜交種; 最后2個條帶: 瘤突蒼耳 The first two strip from left to right: X. sibiricum; Middle strip: Suspected hybrid; Last two strip: X. strumarium
疑似雜合體種子蛋白質鹽溶蛋白電泳結果如圖2所示,部分疑似雜合體種子的鹽溶蛋白條帶與人工雜合體情況類似(圖2紅色箭頭),其水溶蛋白和剩余蛋白無明顯差異(未放入),說明在自然界中,瘤突蒼耳和蒼耳之間存在雜交現象。
2.2 鎘脅迫對瘤突蒼耳、蒼耳及其雜合體種子萌發和幼苗生長的影響
雜交對瘤突蒼耳種子的萌發率、根長、地下部鮮重、總鮮重、地上部干重、總干重及子葉丙二醛含量有顯著影響(P< 0.05, 表1),鎘濃度則對除發芽指數、子葉長、地上部鮮重及總干重外的其余指標均有顯著形影(P< 0.05, 表1),二者交互作用對瘤突蒼耳的根長、丙二醛、葉綠素含量存在顯著影響(P< 0.05, 表1)。雜交對蒼耳的根長、子葉長、丙二醛含量、愈創木酚過氧化物酶活性、葉綠素含量有顯著的影響(P< 0.05, 表1),鎘濃度對根長、子葉長、地上、地下及總鮮重、丙二醛含量、愈創木酚過氧化物酶活性、葉綠素含量均有顯著影響(P< 0.05, 表1),二者交互作用對蒼耳的根長、丙二醛含量、愈創木酚過氧化物酶活性、葉綠素含量存在顯著影響(P< 0.05, 表1)。
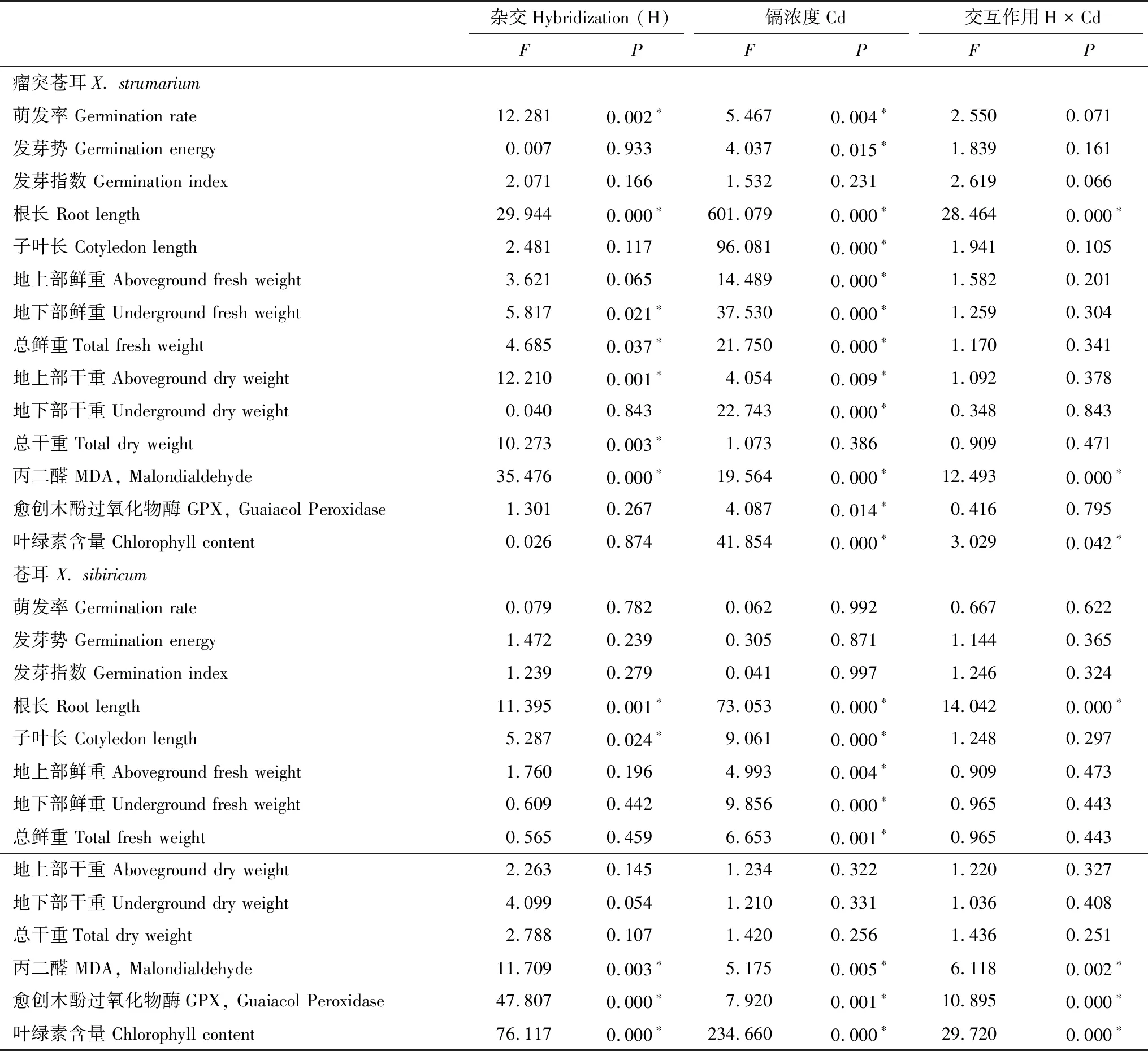
表1 雜交和重金屬鎘對瘤突蒼耳、蒼耳種子萌發和幼苗生長的影響
*:P< 0.05
蒼耳、瘤突蒼耳及雜交瘤突蒼耳種子萌發率隨著Cd濃度的增加而增加。在5個Cd濃度下,瘤突蒼耳和雜交瘤突蒼耳種子萌發率均顯著高于蒼耳和雜交蒼耳(P< 0.05),蒼耳和雜交蒼耳種子萌發率無顯著差異(P> 0.05);在80 mg/L Cd處理下,雜交瘤突蒼耳種子萌發率顯著高于瘤突蒼耳,其余處理下則無顯著性差異(P< 0.05)。在5個Cd濃度下,瘤突蒼耳和雜交瘤突蒼耳種子發芽指數均顯著高于蒼耳和雜交蒼耳(P< 0.05);在80 mg/L Cd處理下,雜交瘤突蒼耳種子發芽指數顯著高于瘤突蒼耳,其余處理下則無顯著性差異(P> 0.05);在120 mg/L Cd處理下,雜交蒼耳種子發芽勢顯著低于蒼耳(P< 0.05),其余處理下則無顯著性差異(P> 0.05)。蒼耳、瘤突蒼耳及其雜合體種子發芽勢表現出與發芽指數一致的規律性。
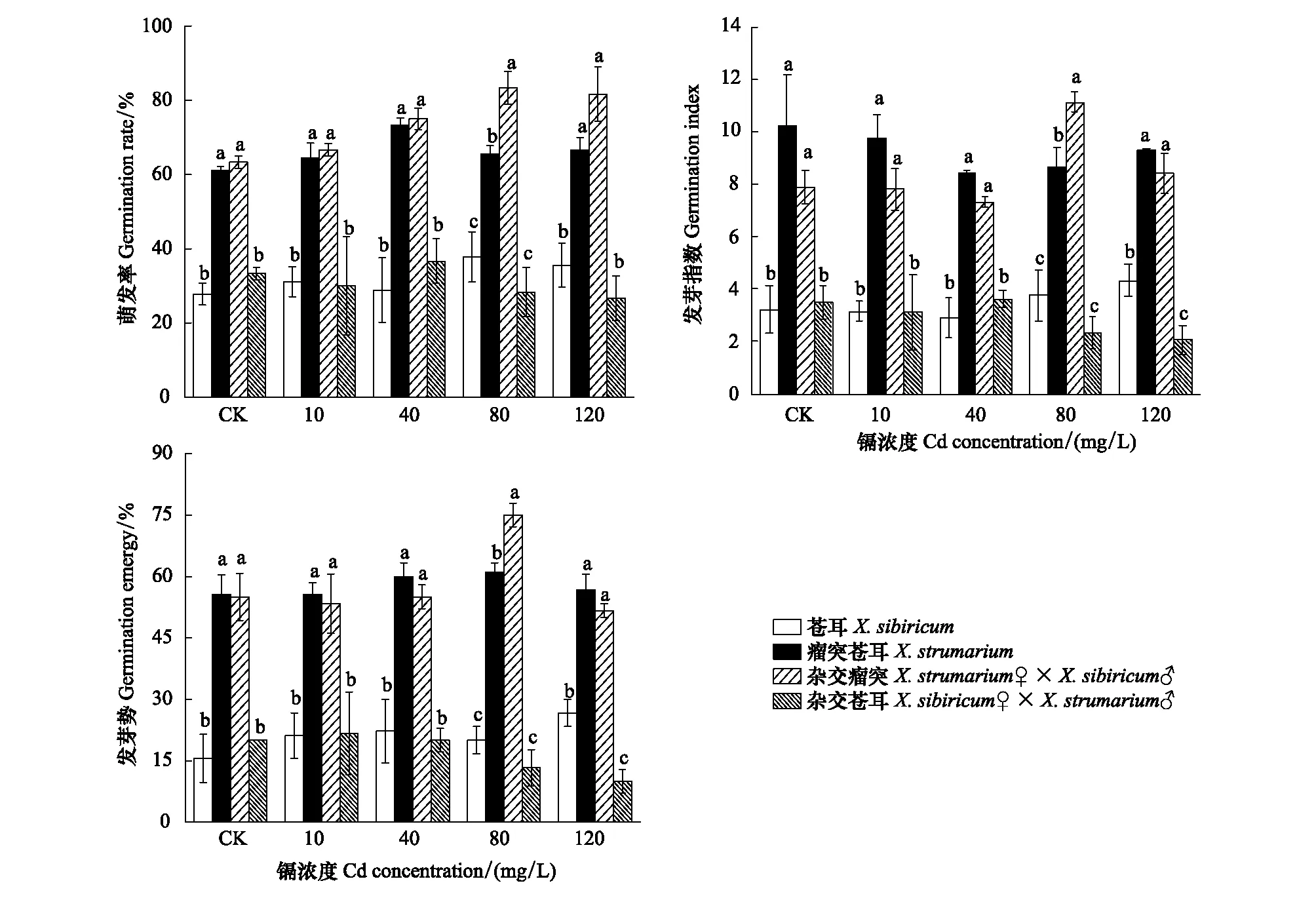
圖3 不同鎘濃度下瘤突蒼耳、蒼耳及其雜合體種子萌發特征Fig.3 Germination feature of X. strumarium, X. sibiricum and their hybrids under different Cd concentration (means±SE)在同一鎘濃度處理下, 不同小寫字母代表蒼耳、瘤突蒼耳及其雜合體存在顯著差異(P < 0.05)。CK: 蒸餾水; 10: 鎘濃度10 mg/L; 40: 鎘濃度40 mg/L; 80: 鎘濃度80 mg/L; 120: 鎘濃度120 mg/L
蒼耳、瘤突蒼耳及其雜合體幼苗根長和子葉長度隨著Cd濃度的增加而降低(圖4)。雜交瘤突蒼耳和雜交蒼耳子葉長在Cd濃度40 mg/L處理下分別顯著低于瘤突蒼耳和蒼耳(P< 0.05);雜交瘤突蒼耳根長在CK和10 mg/L處理下顯著高于瘤突蒼耳 (P< 0.05),在40—120 mg/L處理下顯著低于瘤突蒼耳(P< 0.05),在80和120 mg/L下顯著高于蒼耳(P< 0.05);雜交蒼耳根長在40 mg/L處理下顯著低于蒼耳(P< 0.05)。
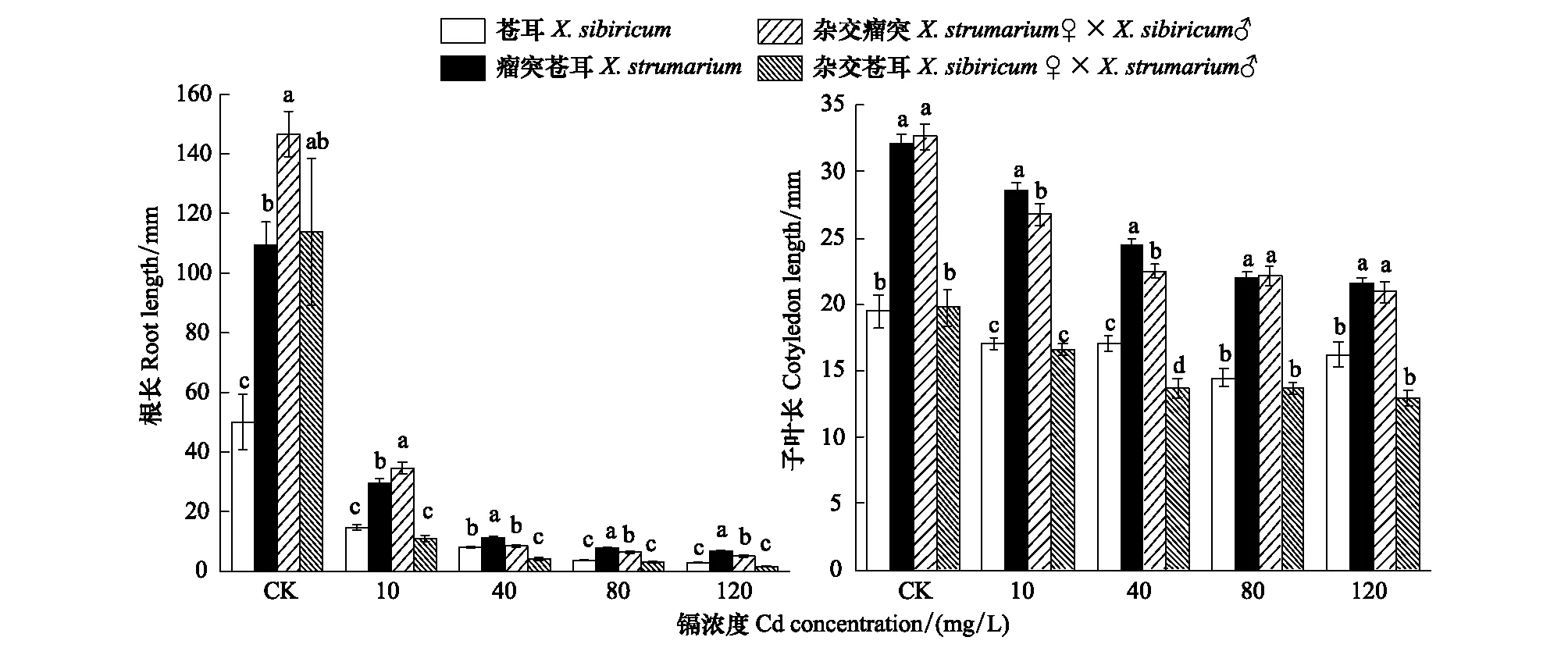
圖4 不同鎘濃度下瘤突蒼耳、蒼耳及其雜合體根長和子葉長Fig.4 Root length and cotyledon length of X. strumarium, X. sibiricum and their hybrids under different Cd concentration (means±SE)
瘤突蒼耳、蒼耳及其雜合體的地上鮮重、地下鮮重、總鮮重及地下干重隨著Cd濃度的增加而降低,其地上干重和總干重則隨著Cd濃度的增加而增加。雜交蒼耳的地上鮮重和總鮮重僅在鎘濃度為120 mg/L時顯著低于蒼耳(P< 0.05),地上、地下及總鮮重在CK下與蒼耳無顯著差異(P> 0.05),但親本瘤突蒼耳顯著高于蒼耳(P< 0.05),說明在CK處理下,雜交使得蒼耳鮮重增加;雜交蒼耳的地上干重和總干重在CK和80 mg/L時與蒼耳無顯著差異(P> 0.05),其余濃度處理均顯著低于蒼耳(P< 0.05);雜交瘤突蒼耳幼苗地上鮮重和總鮮重在鎘濃度為10 mg/L時顯著低于瘤突蒼耳(P< 0.05),地上干重和總干重在鎘濃度為40和120 mg/L時顯著低于瘤突蒼耳(P< 0.05),地下干重在120 mg/L時顯著低于瘤突蒼耳(P< 0.05) 。

圖5 不同鎘濃度下瘤突蒼耳、蒼耳及其雜合體生物量特征Fig.5 Biomass characteristics of X. strumarium, X. sibiricum and their hybrids under different Cd concentration (means±SE)
蒼耳、瘤突蒼耳及其雜合體幼苗子葉丙二醛含量和愈創木酚過氧化物酶活性隨著Cd濃度的增加而增加,葉綠素含量則相反(圖6)。對照組中,雜交瘤突蒼耳和雜交蒼耳的葉綠素總含量分別顯著高于瘤突蒼耳和蒼耳(P< 0.05),在80 mg/L處理下則相反(P< 0.05);在10、40、80 mg/L處理下,瘤突蒼耳的丙二醛含量顯著高于雜交瘤突(P< 0.05),僅在80 mg/L處理下,雜交蒼耳的丙二醛含量顯著高于蒼耳(P< 0.05);雜交瘤突蒼耳和雜交蒼耳的愈創木酚過氧化物酶活性在對照處理下分別顯著高于瘤突蒼耳和蒼耳(P< 0.05);在10、80、120 mg/L處理下,雜交蒼耳的愈創木酚過氧化物酶活性顯著低于蒼耳(P< 0.05)。
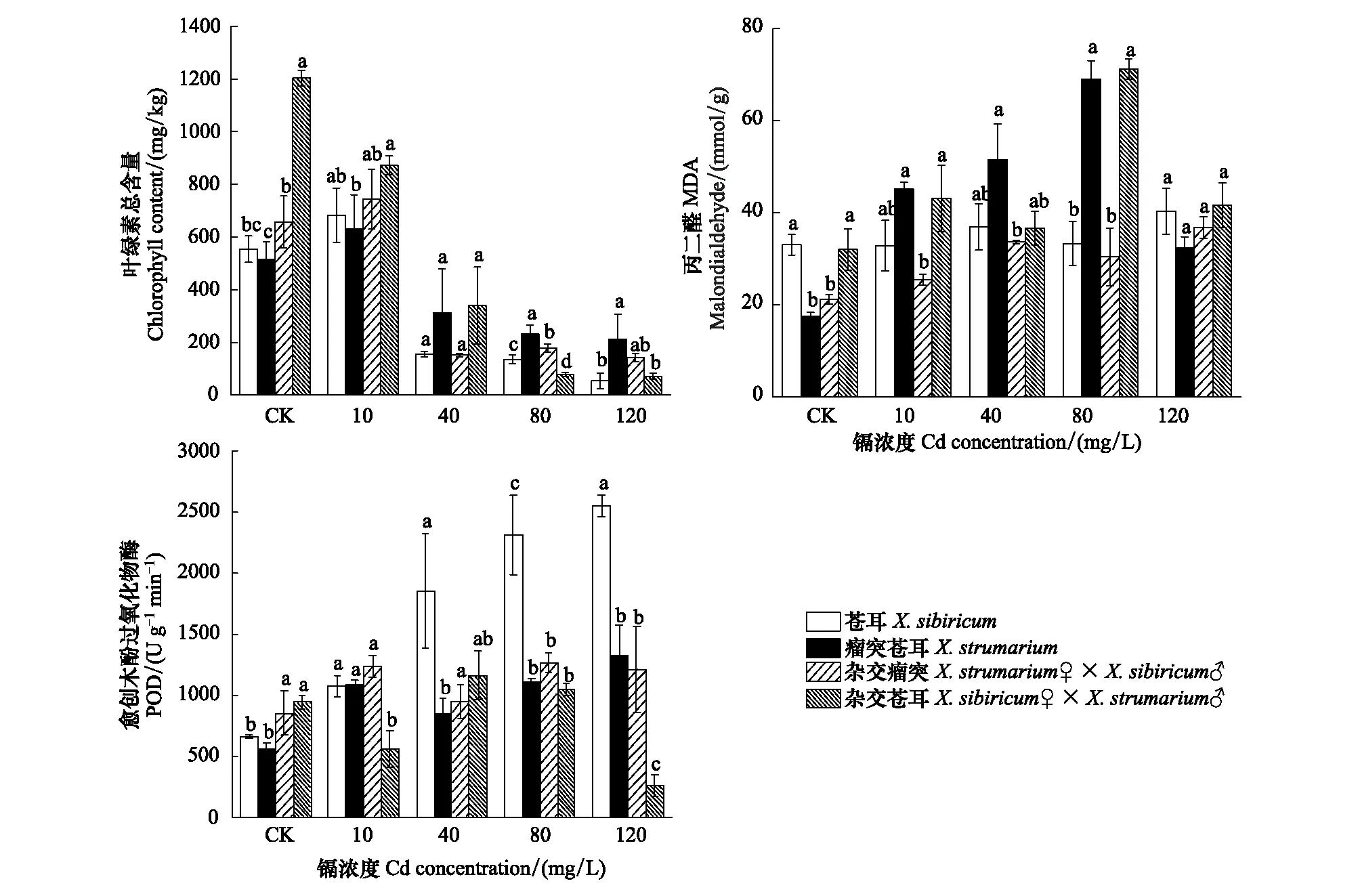
圖6 不同鎘濃度下瘤突蒼耳、蒼耳及其雜合體葉綠素含量、丙二醛及愈創木酚過氧化物酶活性Fig.6 Chlorophyll content, malondialdehyde (MDA) and Guaiacol peroxidase activity (POD) of X. strumarium, X. sibiricum and their hybrids under different Cd concentration (means±SE)
3 討論
種子蛋白質電泳實驗結果顯示,蒼耳和瘤突蒼耳的雜交后代具有親本獨有的某些鹽溶蛋白,這說明雜交使得二者基因之間發生交換和重組,隨后雙因素方差分析表明雜交確實影響了瘤突蒼耳和蒼耳的某些性狀,但這些變化性狀是否有增強了植物對重金屬鎘的耐受性,我們仍需要繼續進行驗證。由于雜交親本篩選比較困難、雜交后代形狀不穩定、雜種優勢受重組、雜合性下降等因素隨世代的增加而消失以及較長的研究周期,目前很少有研究通過入侵植物與本地種雜交來進行超富集植物的篩選,但植物對重金屬的積累受控于體內特有基因表達。比如Liu等[27]對鎘鋅超積累植物伴礦景天Cd耐性基因進行研究,發現伴礦景天體內的ATP酶3基因(SpHMA3)具有Cd專一運輸活性,在非超積累型景天中超量表達可增加其對鎘的耐性和積累性。因此利用雜交改變植物的遺傳特征,這值得我們進行嘗試。
萌發率反映了種子的生命力,發芽勢反映種子出苗快慢,兩者均為鑒定種子質量最直接和穩定的指標,發芽指數則反映了整個發芽期種子活力的綜合特征[28]。本試驗結果顯示,瘤突蒼耳的發芽率、發芽勢及發芽指數隨著重金屬鎘濃度的增加而增加,說明鎘脅迫并未抑制其種子活力,相反還提高了其發芽速度,這與胡小英等[29]研究結果一致。雜交瘤突(X.strumarium♀ ×X.sibiricum♂)種子的萌發情況與瘤突蒼耳類似,且當鎘濃度為80 mg/L時,雜交瘤突(X.strumarium♀ ×X.sibiricum♂)種子的萌發率顯著高于瘤突蒼耳并達到最大(83.33%),這說明了以瘤突蒼耳為母本,蒼耳為父本的雜交后代種子在萌發情況上得到提高,并未受到抑制。相反,雜交蒼耳(X.sibiricum♀ ×X.strumarium♂)不僅在種子萌發率上低于蒼耳,而且在高濃度(120 mg/L)時種子發芽勢和發芽指數均顯著低于蒼耳,說明以蒼耳為母本,瘤突蒼耳為父本的雜交后代種子在高濃度鎘脅迫下的表現弱于蒼耳。
幼苗的生長狀況是植物對逆境脅迫適應的綜合體現,脅迫環境對植物生長的抑制程度直觀地反映了植物體的受傷程度[30]。本研究結果表明,蒼耳、瘤突蒼耳及雜合體幼苗的根長、子葉長、鮮重、葉綠素含量隨著鎘濃度的增加而降低,說明重金屬鎘對幼苗的生長具有明顯的抑制作用。這與張大鵬等[31]、韓寶賀等[32]的研究結果一致。隨著鎘濃度增加,蒼耳、瘤突蒼耳及其雜合體根長降幅遠高于子葉,這是因為在種子萌發過程中,胚根是主要吸收水分和養分的器官,長時間與鎘接觸,所以根長受到的抑制程度高于子葉。
重金屬鎘會導致植物體內自由基和活性氧含量大幅度上升,進而氧化蛋白質、DNA、脂類和碳水化合物,從而擾亂許多生理生化過程[33- 34]。藺永和等研究了野生垂穗披堿草(Elymusnutans)對鋁脅迫的響應,發現隨著鋁濃度的增加,丙二醛含量不斷增加[35]。張虎等研究了紫花苜蓿種子萌發(Medicagosativa)對鈷脅迫的生理生化響應,發現MDA含量隨著鈷濃度的增加而增加[24]。本研究結果與這些研究基本一致。蒼耳和雜交蒼耳的GPX活性持續增加,但在10、40、80、120 mg/L下,蒼耳的GPX卻高于雜交蒼耳(僅在40 mg/L處理下不顯著),說明蒼耳的抗氧化能力高于雜交蒼耳。
本試驗基礎液均采用蒸餾水,排除了不同離子對幼苗生長的影響,植物生長發育取決于植物本身。結果顯示,在對照和低濃度(10 mg/L)脅迫下,雜交瘤突幼苗根長和子葉葉綠素含量均高于瘤突蒼耳;在高濃度鎘(80、120 mg/L)脅迫下,雜交瘤突幼苗根長和子葉葉綠素含量低于瘤突蒼耳。Mobin和Khan研究了Varuna和RH- 30兩種芥菜(Brassicajuncea)在鎘脅迫下的光合活性、色素組成及抗氧化反應,結果顯示當土壤鎘濃度為25、50、100 mg/kg時,RH- 30根和葉中的鎘濃度均顯著高于Varuna,但Varuna中葉綠素總含量分別降低20.0%,35.0%和50.0%,RH- 30則分別降低31.0%,43.0%和57.0%[36]。相似的結果也在Shi等[37]、He等[38]、Zhang等[39]、Fang等[40]研究中發現。前人研究結果已經證明鎘可以通過抑制葉綠素合成酶和CO2固定酶的活性[36, 41]或光系統色素蛋白復合物的聚集[42]來擾亂葉綠體功能。結合我們的實驗結果,我們推測雜交有可能提高了瘤突蒼耳在高濃度鎘脅迫時對重金屬鎘的轉運能力,幼苗根系吸收大量的鎘并向上運輸給子葉,這導致根系生長受到抑制、子葉葉綠素含量下降。同時我們發現當鎘濃度為10、40、80 mg/L時,雜交瘤突幼苗MDA含量顯著低于瘤突蒼耳,其GPX活性高于瘤突蒼耳,但并不顯著,因此雜交瘤突抗氧化酶系統是否在這一過程中發揮了作用仍需要我們繼續研究。雜交是否提高了瘤突蒼耳對重金屬鎘的吸收與轉運,我們仍然需要后續盆栽實驗給出確切的證據,同時深入探究雜交在這一過程中發揮的作用。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
