金屬納米材料的植物生物效應(yīng)及其多組學(xué)研究進(jìn)展
2020-03-14 08:01:54韋革宏
陳 春 ,劉 爽 ,韋革宏 *
金屬納米材料(MNMs)是目前應(yīng)用最為廣泛的一類(lèi)納米材料,按其化學(xué)成分的不同可分為:金屬、金屬合金及金屬化合物3大類(lèi)[1],商品化MNMs多以金屬單質(zhì)與金屬氧化物居多[2]。近年來(lái),伴隨著納米技術(shù)在農(nóng)業(yè)領(lǐng)域中的快速發(fā)展,肥料、農(nóng)藥、獸藥以及飼料等納米化農(nóng)業(yè)投入品的使用也日益增多,這類(lèi)投入品的直接使用將會(huì)引起MNMs在農(nóng)業(yè)生境中的暴露加劇[3];人類(lèi)日常使用的洗滌產(chǎn)品、化妝品及個(gè)人護(hù)理品等中的MNMs,首先會(huì)伴隨生活污水的排放進(jìn)入污水處理廠(chǎng),然后通過(guò)污水灌溉或污泥農(nóng)田施用等方式潛入土壤中;另外,大氣中存在的MNMs也可通過(guò)團(tuán)聚沉降或降水等方式遷移潛入土壤,最終土壤將成為大多數(shù)MNMs的最終歸宿[4]。存在于土壤中的MNMs會(huì)對(duì)土壤生態(tài)系統(tǒng)產(chǎn)生何種效應(yīng),已經(jīng)引起國(guó)內(nèi)外學(xué)者的廣泛關(guān)注,現(xiàn)有研究表明MNMs在植物體內(nèi)的生物富集會(huì)沿陸地食物鏈逐級(jí)放大,從而可能對(duì)整個(gè)陸地生態(tài)系統(tǒng)產(chǎn)生潛在的環(huán)境風(fēng)險(xiǎn)[5]。組學(xué)技術(shù)的快速發(fā)展則為探究MNMs植物毒性效應(yīng)提供了新的思路,結(jié)合傳統(tǒng)植物毒理學(xué)手段,有望更加準(zhǔn)確與深入闡釋MNMs毒性的分子機(jī)制。
1 MNMs傳統(tǒng)植物毒理學(xué)研究
關(guān)于MNMs植物效應(yīng)方面的研究,傳統(tǒng)毒理學(xué)方法主要采用葉面噴施、水培或土壤基質(zhì)染毒的方式,觀(guān)察MNMs暴露脅迫下植物生長(zhǎng)、亞細(xì)胞以及生理生化水平層次上的生物效應(yīng)。
1.1 植物生長(zhǎng)水平
植物生長(zhǎng)效應(yīng)的評(píng)價(jià)指標(biāo)一般包括發(fā)芽率、莖長(zhǎng)、根長(zhǎng)、葉片數(shù)和生物量等。早期國(guó)內(nèi)外學(xué)者陸續(xù)開(kāi)展了典型納米顆粒(ZnO、Ag、CuO、CeO2等)對(duì)大豆、水稻、小麥、苜蓿、黃瓜等作物生長(zhǎng)影響的研究[6-9]。在營(yíng)養(yǎng)液、砂質(zhì)或土壤等不同暴露基質(zhì)中,某些納米顆粒(NPs)暴露濃度超過(guò)某一閾值時(shí),便會(huì)對(duì)植物生長(zhǎng)造成一些負(fù)面效應(yīng),主要表現(xiàn)為種子萌發(fā)和根伸長(zhǎng)受到抑制以及生物量的顯著減少等。通常水培基質(zhì)中NPs的效應(yīng)濃度水平要遠(yuǎn)低于土壤,這是由于NPs進(jìn)入土壤之后會(huì)發(fā)生溶解、團(tuán)聚等化學(xué)變化。而土壤復(fù)雜的理化因素也會(huì)影響NPs的穩(wěn)定性與生物可利用性[10-11]。其中,土壤pH值是影響土壤中可溶性金屬離子類(lèi)MNMs毒性效應(yīng)的關(guān)鍵因子,研究發(fā)現(xiàn)酸性土壤中NPs對(duì)植物的毒性強(qiáng)度要高于堿性土壤,這是由于酸性條件MNMs金屬離子的釋放速率與移動(dòng)性增強(qiáng)[12-13];此外,土壤離子強(qiáng)度或有機(jī)質(zhì)含量也會(huì)影響NPs在土壤中的環(huán)境行為,離子強(qiáng)度的升高可引起NPs團(tuán)聚速率的增加[14],有機(jī)質(zhì)中腐植酸等物質(zhì)可吸附于NPs顆粒表面,增加NPs表面靜電斥力,從而減緩NPs的團(tuán)聚效應(yīng)[15-16],促使NPs在植物體內(nèi)遷移,提高生物可利用性,最終加劇植物的毒性水平。研究發(fā)現(xiàn)粗砂暴露條件下,500 mg·kg-1CuO NPs和500 mg·kg-1CeO2NPs不會(huì)對(duì)胡蘿卜生長(zhǎng)產(chǎn)生脅迫,但是相同濃度ZnO NPs暴露則顯著抑制了胡蘿卜總生物量的積累,這表明同一種植物對(duì)于不同種類(lèi)MNMs暴露的響應(yīng)與耐受程度存在差異[17];同一種MNMs暴露對(duì)于不同種類(lèi)植物生長(zhǎng)方面的效應(yīng)壓力也有所不同,例如水培條件下1600 mg·L-1ZnONPs暴露可導(dǎo)致紫花苜蓿和番茄種子的萌發(fā)率受到顯著抑制,而黃瓜種子的萌發(fā)率反而提高,表明在種子萌發(fā)階段不同種類(lèi)植物對(duì)于MNMs暴露脅迫的響應(yīng)表現(xiàn)與耐受力存在差異[18]。
1.2 亞細(xì)胞水平
植物亞細(xì)胞結(jié)構(gòu)主要包括細(xì)胞壁、細(xì)胞膜、細(xì)胞核和細(xì)胞器(線(xiàn)粒體、葉綠體、內(nèi)質(zhì)網(wǎng)等)。對(duì)于植物亞細(xì)胞結(jié)構(gòu)的觀(guān)察,不僅能幫助判別NPs是否進(jìn)入植物體內(nèi),了解NPs的入胞方式以及掌握其在細(xì)胞內(nèi)的轉(zhuǎn)運(yùn)等關(guān)鍵信息,還能確定MNMs作用的靶標(biāo)器官并定位損傷組織,這也是探析NPs植物毒性機(jī)理的必要基礎(chǔ)。目前研究發(fā)現(xiàn)植物細(xì)胞可通過(guò)吞噬或胞飲等內(nèi)吞作用攝取NPs,并且證實(shí)了ZnO、Ag、CuO、TiO2、ZVI(納米零價(jià)鐵)等多種MNMs能夠進(jìn)入植物根細(xì)胞并在細(xì)胞內(nèi)積累[19-22]。NPs的納米尺寸效應(yīng)與其獨(dú)特的表面活性效應(yīng),賦予其更易穿過(guò)植物細(xì)胞屏障的能力。NPs首先附著于植物初生根表面,隨植物的生長(zhǎng)發(fā)育進(jìn)入根尖、側(cè)根原基及根毛區(qū)域中,并且通過(guò)細(xì)胞間隙或胞間連絲,伴隨養(yǎng)分運(yùn)輸?shù)侥举|(zhì)部,進(jìn)而縱向易位到莖葉;粒徑稍大的NPs可能會(huì)堵塞細(xì)胞孔隙或誘導(dǎo)細(xì)胞表面形成更大孔隙,從而影響根部的水分及養(yǎng)分運(yùn)輸[23-25]。此外,NPs也可直接通過(guò)角質(zhì)層或氣孔被植物葉片吸收,憑借維管組織運(yùn)輸?shù)街仓昶渌课籟26-27]。進(jìn)入葉肉細(xì)胞的NPs會(huì)導(dǎo)致葉綠體腫脹,類(lèi)囊體損傷及質(zhì)體小球增多變大,這些生物學(xué)現(xiàn)象皆反映出葉綠體可能是MNMs作用的靶位點(diǎn)之一,但具體作用機(jī)制有待于進(jìn)一步研究[28-29]。植物生理生化變化與形態(tài)結(jié)構(gòu)緊密相連,MNMs的暴露脅迫除了影響植物細(xì)胞形態(tài)結(jié)構(gòu)發(fā)生變化外,還伴隨著復(fù)雜的生理生化變化。
1.3 生理生化水平
大量研究證實(shí)MNMs暴露脅迫對(duì)植物的毒性機(jī)制之一是產(chǎn)生氧化脅迫。植物在抵御氧化脅迫時(shí),一方面會(huì)通過(guò)產(chǎn)生如還原型谷胱甘肽(GSH)等小分子有機(jī)物來(lái)消耗體內(nèi)的活性氧簇(ROS),另一方面會(huì)通過(guò)誘導(dǎo)體內(nèi)一系列抗氧化酶的活性來(lái)清除ROS。ROS主要包括單線(xiàn)態(tài)氧(1O2)、超氧陰離子自由基(O-2·)、過(guò)氧化氫(H2O2)和羥基自由基(HO·)等,其通常產(chǎn)生于植物細(xì)胞葉綠體、線(xiàn)粒體及過(guò)氧化物酶體等亞細(xì)胞結(jié)構(gòu)中[30]。植物抗氧化的第一道防線(xiàn)是通過(guò)超氧化物歧化酶(SOD)催化O-2·產(chǎn)生歧化產(chǎn)物H2O2,而后由過(guò)氧化氫酶(CAT)、過(guò)氧化物酶(POD)或抗壞血酸過(guò)氧化物酶(APX)等進(jìn)一步清除H2O2。同時(shí),抗壞血酸、谷胱甘肽、類(lèi)胡蘿卜素、生育酚及酚類(lèi)化合物等小分子代謝物也參與植物ROS的清除[31]。低水平ROS可作為信號(hào)分子參與植物防御反應(yīng),傳遞信息至抗氧化防御系統(tǒng),從而誘導(dǎo)抗氧化酶活性升高;然而,當(dāng)ROS水平超出抗氧化防御系統(tǒng)的清除能力時(shí),便會(huì)導(dǎo)致抗氧化酶蛋白組分發(fā)生氧化損傷,從而使抗氧化酶水平逐漸下降,進(jìn)而破壞抗氧化酶系統(tǒng)。同時(shí),植物體內(nèi)積累的ROS會(huì)進(jìn)一步與脂類(lèi)、蛋白質(zhì)、核酸等生物大分子直接反應(yīng)導(dǎo)致機(jī)體氧化損傷。測(cè)定植物體內(nèi)抗氧化酶酶活和脂質(zhì)過(guò)氧化水平,通常可以表征植物抵御MNMs脅迫的能力以及機(jī)體氧化損傷的程度。多數(shù)研究皆發(fā)現(xiàn)MNMs暴露可誘導(dǎo)大豆、豌豆、水稻、擬南芥等植物細(xì)胞中ROS的產(chǎn)生,進(jìn)而引起一系列的植物防御反應(yīng),主要表現(xiàn)為丙二醛(MDA)水平升高、抗氧化酶活性改變、葉綠素水平降低以及氧化應(yīng)激損傷程度加劇等[32-35]。蘭麗貞等[36]研究發(fā)現(xiàn)在水培條件下,隨著TiO2NPs暴露濃度增加(0.05~1.5 mg·L-1)與暴露時(shí)間(0、7 d和14 d)延長(zhǎng),擬南芥葉片及根部中SOD水平呈逐漸下降的趨勢(shì),同時(shí)植物體內(nèi)MDA含量呈遞增趨勢(shì);?eki?等[37]發(fā)現(xiàn)高劑量Ag NPs(80 mg·L-1)在水培暴露條件下可引起番茄體內(nèi)SOD、APX活性顯著降低,并引發(fā)DNA損傷和脂質(zhì)過(guò)氧化作用,繼而對(duì)番茄植物造成一定的毒害。
2 MNMs植物生物效應(yīng)的多組學(xué)研究
2.1 轉(zhuǎn)錄組學(xué)(Transcriptomics)
轉(zhuǎn)錄組學(xué)是基于RNA水平研究細(xì)胞基因表達(dá)及調(diào)控規(guī)律的一門(mén)學(xué)科[38],目前檢測(cè)技術(shù)主要分為兩大類(lèi):(1)基于雜交的DNA微陣列(Microarray);(2)基于測(cè)序分析的大規(guī)模平行測(cè)序技術(shù)(MPSS)、表達(dá)序列標(biāo)簽技術(shù)(EST)、基因表達(dá)系列分析技術(shù)(SAGE)和RNA測(cè)序技術(shù)(RNA-Seq)等。其中,RNA-Seq由于其高通量、高分辨率、低背景且不依賴(lài)于基因組信息等優(yōu)勢(shì),而被廣泛應(yīng)用于轉(zhuǎn)錄組學(xué)研究[39-40]。近年來(lái),轉(zhuǎn)錄組學(xué)在植物逆境脅迫、抗病防御及發(fā)育調(diào)控等方面的應(yīng)用越來(lái)越廣泛。已經(jīng)有不少學(xué)者通過(guò)轉(zhuǎn)錄組學(xué)技術(shù),得到植物體在某些MNMs暴露脅迫下的植物差異轉(zhuǎn)錄圖譜和代謝通路,并篩選出某些關(guān)鍵表達(dá)差異基因,相關(guān)部分研究列于表1。MNMs的暴露可誘導(dǎo)植物某些基因在轉(zhuǎn)錄水平的差異表達(dá),這些差異基因主要與植物體內(nèi)響應(yīng)非生物脅迫(氧化脅迫、水分脅迫、鹽脅迫和滲透脅迫等)和生物脅迫(傷口刺激和病原體入侵),重金屬解毒及轉(zhuǎn)運(yùn)和DNA復(fù)制、轉(zhuǎn)錄及翻譯調(diào)控相關(guān)。在水培基質(zhì)中,ZnO NPs(4 mg·L-1與 100 mg·L-1)[41-42]、Ag NPs(5 mg·L-1)[43]、CuO NPs(10 mg·L-1)[44]暴露下的擬南芥,500 mg·L-1ZnO NPs[45]暴露下的玉米體內(nèi)SOD、POD等抗氧化酶基因的表達(dá)皆會(huì)受到誘導(dǎo)而顯著上調(diào);1 mg·L-1Cu NPs暴露脅迫下,小麥體內(nèi)參與苯丙烷代謝(合成酚類(lèi)化合物清除ROS)與脯氨酸代謝(植物體一種有效抗氧化劑[46])相關(guān)的基因表達(dá)顯著上調(diào)[47]。此外,暴露在含有1400 mg·kg-1ZnONPs、180 mg·kg-1Ag NPs和5000 mg·kg-1TiO2NPs的土壤中(老化6個(gè)月后),蒺藜苜蓿體內(nèi)與抗氧化脅迫相關(guān)的基因也會(huì)表現(xiàn)出顯著上調(diào)[48]。這些現(xiàn)象都表明MNMs暴露誘導(dǎo)了植物抗氧化防御系統(tǒng)的響應(yīng)。質(zhì)膜內(nèi)在蛋白與水通道蛋白可介導(dǎo)植物水分與養(yǎng)分的跨膜和長(zhǎng)距離運(yùn)輸,其相關(guān)基因的響應(yīng)表達(dá)可能是由于MNMs納米尺度的物理作用引起水通道堵塞,從而導(dǎo)致植物發(fā)生水分脅迫。同時(shí),4 mg·L-1ZnONPs[41]、10 mg·L-1CuONPs[44]水培暴露可導(dǎo)致擬南芥體內(nèi)與根發(fā)育相關(guān)的基因(包括細(xì)胞壁修飾、根形態(tài)發(fā)生和根毛發(fā)育等)下調(diào)表達(dá),表明了MNMs會(huì)破壞根表細(xì)胞并抑制植物根的發(fā)育;而10 mg·L-1Al2O3NPs水培暴露則顯著增加了擬南芥體內(nèi)與根發(fā)育和伸長(zhǎng)相關(guān)基因的轉(zhuǎn)錄表達(dá)[49],體現(xiàn)了植物對(duì)于不同種類(lèi)MNMs脅迫在轉(zhuǎn)錄水平響應(yīng)表達(dá)的特異性。重金屬轉(zhuǎn)運(yùn)及解毒蛋白(鋅轉(zhuǎn)運(yùn)蛋白、金屬硫蛋白和重金屬ATP酶等)在植物重金屬吸收及解毒調(diào)控中發(fā)揮著關(guān)鍵作用,這類(lèi)基因的顯著上調(diào)表明MNMs釋放的重金屬離子可能是其產(chǎn)生毒性的一種主要機(jī)制[50-52]。研究還發(fā)現(xiàn)ZnONPs在水培暴露(4、100 mg·L-1)下可以引起擬南芥微管蛋白相關(guān)基因的下調(diào)表達(dá),而擬南芥微管的重組與微管蛋白單體的加速降解存在一定的聯(lián)系,這些微管蛋白又參與細(xì)胞分裂過(guò)程,可能預(yù)示MNMs會(huì)影響植物的細(xì)胞分裂,從而給植物生長(zhǎng)帶來(lái)負(fù)面的影響[41-42,53]。
轉(zhuǎn)錄組學(xué)雖然可以展現(xiàn)轉(zhuǎn)錄水平植物對(duì)MNMs暴露的響應(yīng),但其僅能反映基因表達(dá)的中間狀態(tài),而蛋白質(zhì)作為基因的最終產(chǎn)物,在全面分析基因表達(dá)的過(guò)程中具有不可替代的作用。另外,由于存在轉(zhuǎn)錄及翻譯后的調(diào)控與修飾,基因的轉(zhuǎn)錄和其產(chǎn)物蛋白的表達(dá)有時(shí)存在不一致性,并且利用轉(zhuǎn)錄組學(xué)研究無(wú)參考基因組的非模式植物時(shí),一些測(cè)序轉(zhuǎn)錄本難以進(jìn)行注釋?zhuān)虼诵杞柚鞍踪|(zhì)組學(xué)進(jìn)一步完善,以期更加全面地闡釋植物對(duì)MNMs暴露的響應(yīng)機(jī)制[54]。

表1 植物響應(yīng)MNMs暴露的轉(zhuǎn)錄組學(xué)研究的主要發(fā)現(xiàn)Table 1 Transcriptomic response of plant exposed to metal nanomaterials(MNMs)
2.2 蛋白質(zhì)組學(xué)(Proteomics)
蛋白質(zhì)是生物體生命活動(dòng)的主要承擔(dān)者,是基因功能的執(zhí)行者。蛋白質(zhì)組學(xué)是通過(guò)鑒定生物體全套蛋白質(zhì)的組成及表達(dá)水平,從細(xì)胞水平研究其變化規(guī)律、修飾情況以及相互作用的一門(mén)學(xué)科[55-56]。蛋白質(zhì)組學(xué)分析的主要流程分為:蛋白質(zhì)的分離純化、鑒定以及生物信息學(xué)分析。蛋白質(zhì)的分離技術(shù)主要包括電泳和色譜。鑒定技術(shù)一般分為質(zhì)譜和同位素標(biāo)記定量技術(shù)。生物信息學(xué)分析主要是指通過(guò)數(shù)據(jù)庫(kù)對(duì)蛋白質(zhì)結(jié)構(gòu)和功能進(jìn)行科學(xué)預(yù)測(cè)。目前,蛋白質(zhì)氨基酸序列數(shù)據(jù)庫(kù)有PIR(國(guó)際蛋白質(zhì)序列數(shù)據(jù)庫(kù))、SWISS-PROT(瑞士,經(jīng)過(guò)注釋的蛋白質(zhì)序列數(shù)據(jù)庫(kù))等,蛋白質(zhì)結(jié)構(gòu)數(shù)據(jù)庫(kù)主要有PDB、SWISS-2DPAGE(瑞士,經(jīng)過(guò)注釋的二維聚丙烯酰胺凝膠電泳數(shù)據(jù)庫(kù))等[40,57-58]。近期采用蛋白質(zhì)組學(xué)技術(shù)研究MNMs植物效應(yīng)的相關(guān)結(jié)果歸納于表2。GSH在植物應(yīng)對(duì)外界脅迫過(guò)程中起著關(guān)鍵作用,不僅能夠清除細(xì)胞內(nèi)的活性氧自由基,還能通過(guò)與細(xì)胞內(nèi)的重金屬離子螯合起到解毒作用。Ag NPs水培暴露下的水稻(30~60 mg·L-1)[59]與蕓芥(10 mg·L-1)[60],其植物體內(nèi)多種與 GSH合成相關(guān)的蛋白酶水平皆顯著提升,表明植物通過(guò)增加GSH的合成來(lái)抵御MNMs引起的氧化脅迫;PYK10(β-葡萄糖苷酶23)是植物細(xì)胞為抵御外界環(huán)境脅迫而形成的一種新型內(nèi)質(zhì)網(wǎng)衍生結(jié)構(gòu),它是內(nèi)質(zhì)網(wǎng)體(ER Body)的主要組成成分,機(jī)械損傷或外源植物激素均可誘導(dǎo)ER Body的產(chǎn)生。10 mg·L-1Ag NPs水培暴露可導(dǎo)致蕓芥根部PYK10水平升高,表明MNMs暴露可能對(duì)蕓芥根部造成一定的機(jī)械損傷[60-63]。光合作用、三羧酸循環(huán)以及糖酵解等代謝途徑是植物體內(nèi)主要的產(chǎn)能途徑,25 mg·kg-1Cu NPs和Fe NPs土壤暴露下的小麥體內(nèi)參與糖酵解相關(guān)蛋白的上調(diào)表達(dá)[64]、10 mg·L-1Ag NPs水培暴露下的煙草[65]及小麥[66]體內(nèi)參與糖酵解及ATP合成相關(guān)蛋白的上調(diào)表達(dá),菜豆體內(nèi)光系統(tǒng)相關(guān)蛋白在250、2000 mg·kg-1CeO2NPs土壤暴露下被顯著激活[67],這些產(chǎn)能途徑的激活可以幫助細(xì)胞產(chǎn)生更多的ATP與還原力(如NADPH),從而迅速為植物合成防御化合物提供能量。此外,還有一些與蛋白質(zhì)折疊與易位相關(guān)的蛋白質(zhì)(例如蛋白酶體、NAC轉(zhuǎn)錄因子和內(nèi)質(zhì)網(wǎng)腔結(jié)合蛋白等)的表達(dá)水平也發(fā)生了變化,表明MNMs脅迫會(huì)干擾蛋白質(zhì)的轉(zhuǎn)運(yùn)及翻譯后修飾[59-60,66-68]。然而,由于轉(zhuǎn)錄、轉(zhuǎn)錄后及翻譯后調(diào)節(jié)機(jī)制、蛋白表達(dá)時(shí)間的滯后性以及測(cè)序背景噪音等原因,使轉(zhuǎn)錄組及蛋白組數(shù)據(jù)之間的相關(guān)性并不高。考慮到植物小分子代謝物是其上游基因與蛋白質(zhì)功能性變化的最終體現(xiàn),借助植物代謝組學(xué)可廣泛篩選一些差異代謝物,從而能夠更直接準(zhǔn)確地了解植物體在逆境脅迫下的生理狀態(tài),并建立代謝組學(xué)與植物表型之間的內(nèi)在聯(lián)系[70-71]。
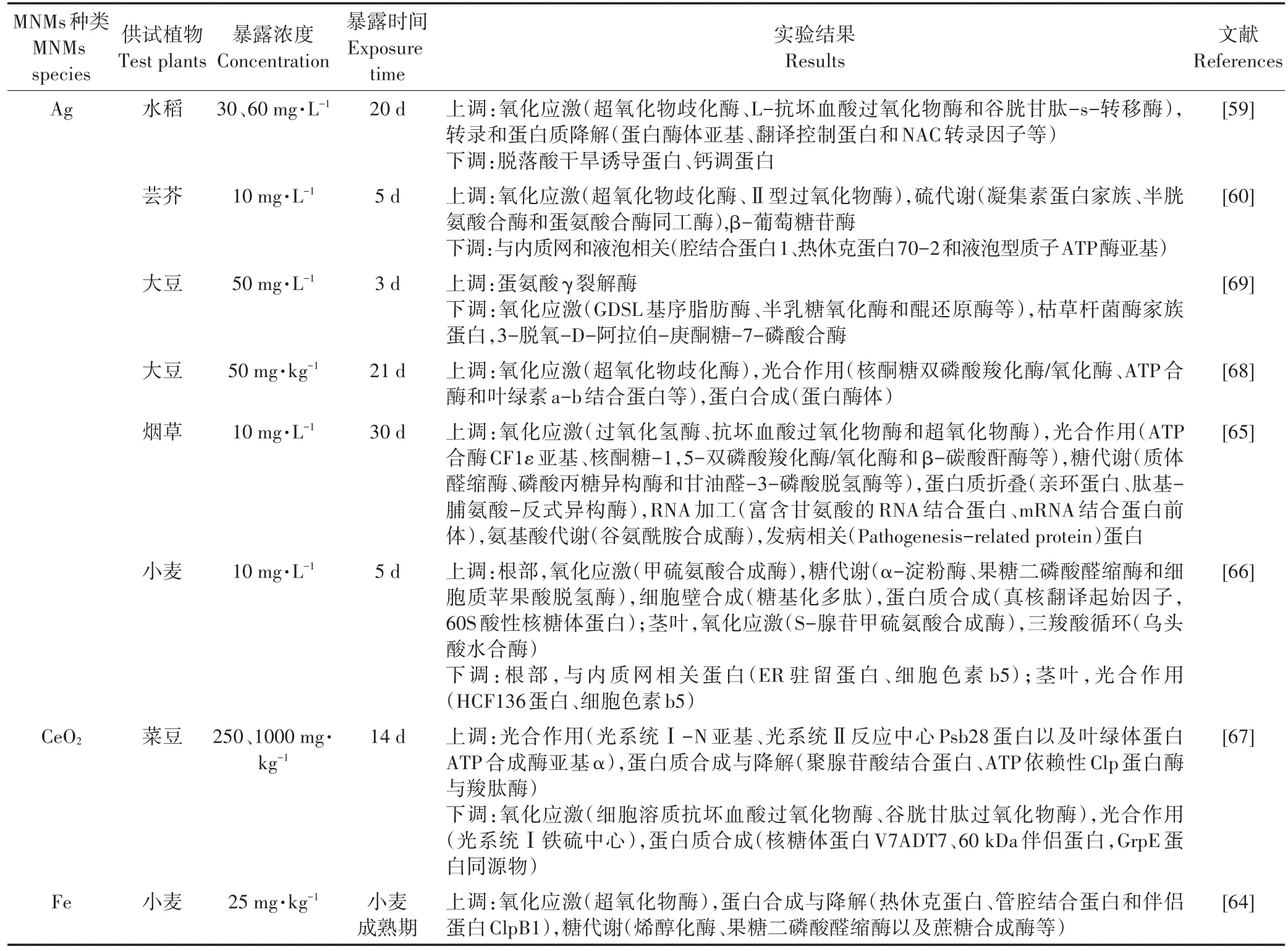
表2 植物響應(yīng)MNMs暴露的蛋白質(zhì)組學(xué)研究的主要發(fā)現(xiàn)Table 2 Proteomics response of plant exposed to metal nanomaterials(MNMs)
2.3 代謝組學(xué)(Metabolomics)
生物體的代謝物通常由多種性質(zhì)不同的小分子化合物組成,是細(xì)胞調(diào)節(jié)過(guò)程的終端產(chǎn)物,能直觀(guān)反映生物體對(duì)環(huán)境變化的響應(yīng)[72-73]。代謝組學(xué)可靈敏地反映生物體在逆境脅迫下代謝水平的細(xì)微變化,通過(guò)明確代謝物對(duì)各種應(yīng)激源的響應(yīng),建立與生物體表型之間的內(nèi)在關(guān)聯(lián)。代謝組學(xué)根據(jù)其研究目的主要分為靶向代謝組學(xué)和非靶向代謝組學(xué):靶向代謝組學(xué)只對(duì)特定的已知代謝物進(jìn)行定量分析,常用于對(duì)已知代謝途徑進(jìn)行深入探究;非靶向代謝組學(xué)可對(duì)生物體內(nèi)源性代謝物進(jìn)行系統(tǒng)分析,獲取特定實(shí)驗(yàn)條件下的差異代謝物,在生物標(biāo)志物與代謝通路的發(fā)現(xiàn)等方面應(yīng)用較為廣泛[74-75]。目前代謝組學(xué)普遍采用的研究手段包括兩類(lèi):核磁共振和質(zhì)譜-色譜聯(lián)用技術(shù)。核磁共振技術(shù)具有無(wú)需樣品前處理、無(wú)損性、無(wú)偏向性、實(shí)時(shí)動(dòng)態(tài)等優(yōu)點(diǎn),但靈敏度相對(duì)較低;而質(zhì)譜-色譜聯(lián)用技術(shù)因其靈敏度高、分辨率好,且能同時(shí)檢測(cè)多種組分等優(yōu)點(diǎn),逐漸被廣泛應(yīng)用于代謝組學(xué)領(lǐng)域,其中應(yīng)用最為廣泛的是氣相色譜-質(zhì)譜技術(shù)和液相色譜-質(zhì)譜技術(shù)[71,76-77]。有關(guān)MNMs植物效應(yīng)代謝組學(xué)近期研究的主要發(fā)現(xiàn)總結(jié)于表3。Ag NPs葉面噴施后(10、100 mg·L-1)的黃瓜[78]、Cu NPs土壤(200、400、800 mg·kg-1)[79-80]及水培暴露(10、20 mg·L-1)下的黃瓜[81]、CeO2NPs土壤暴露(250、2000 mg·kg-1)下的菜豆[67]以及TiO2NPs水培暴露(100、250、500 mg·L-1)下的水稻[82],這些植物體內(nèi)酚類(lèi)化合物的水平均顯著升高。酚類(lèi)化合物是植物體內(nèi)一類(lèi)具有抗氧化活性的次級(jí)代謝產(chǎn)物,它們的積累可能是植物抵御MNMs氧化脅迫的策略之一[78]。氨基酸是植物初級(jí)代謝的重要組成部分,也是植物體內(nèi)氮代謝的主要形式,Ag NPs葉面噴施(10、100 mg·L-1)[78]、Cu NPs土壤暴露(200、400、800 mg·kg-1)[79-80,83]和水培暴露(10、20 mg·L-1)[81]都會(huì)誘導(dǎo)黃瓜體內(nèi)氨基酸的組成發(fā)生變化,表明MNMs可能會(huì)干擾植物體的氮代謝過(guò)程。Ag NPs暴露后的擬南芥(12.5 mg·kg-1土壤暴露)[84]、黃瓜(10、100 mg·L-1葉面噴施)[78],Cu NPs水培暴露(10、20 mg·L-1)后的黃瓜[81]以及TiO2NPs水培暴露(100、250、500 mg·L-1)后的水稻[82]體內(nèi)蘋(píng)果酸、琥珀酸、異檸檬酸等三羧酸循環(huán)中間體發(fā)生積累,而三羧酸循環(huán)是植物體氧化產(chǎn)能的主要途徑之一,植物體可能通過(guò)這種方式為植物抵御MNMs脅迫和修復(fù)受損大分子等提供足夠的能量。此外,10、20 mg·L-1Cu NPs水培暴露與10、100 mg·L-1Ag NPs葉面噴施處理后,黃瓜體內(nèi)壬酸(膜損傷的指標(biāo))和十五烷酸水平(磷脂雙分子層的主要組分)分別受到顯著上調(diào),這些現(xiàn)象可能預(yù)示著細(xì)胞膜完整性遭到破壞[78-81]。
3 MNMs與植物相互作用機(jī)制
植物體在長(zhǎng)期適應(yīng)外界環(huán)境的過(guò)程中形成了多種防御與耐受機(jī)制,如改變自身形態(tài)結(jié)構(gòu)、調(diào)節(jié)基因表達(dá)和改變代謝物水平等,從而保護(hù)植物細(xì)胞免受各類(lèi)脅迫的影響。目前研究表明MNMs產(chǎn)生植物毒性的來(lái)源主要是顆粒的尺度效應(yīng)、表面活性以及自身釋放的游離金屬離子[86]。這種來(lái)源途徑的不同也造成了不同種類(lèi)MNMs的毒性差異。對(duì)于金屬離子溶解度大的MNMs如ZnONPs、Ag NPs等,其植物毒性主要與其釋放出的金屬離子所關(guān)聯(lián),而對(duì)于部分難溶性或溶解度低的MNMs如TiO2NPs、SiO2NPs,其與細(xì)胞的直接接觸以及表面光敏特性等被認(rèn)為是導(dǎo)致細(xì)胞毒性的主要因素[87]。

表3 植物響應(yīng)MNMs暴露的代謝組學(xué)研究的主要發(fā)現(xiàn)Table 3 Metabolomics response of plant exposed to metal nanomaterials(MNMs)
由此推測(cè)MNMs與植物體相互作用的機(jī)制主要包括以下幾個(gè)方面(圖1):(1)MNMs顆粒物理過(guò)程引發(fā)的機(jī)械效應(yīng)。進(jìn)入土壤中的MNMs一部分會(huì)黏附于植物根表,并在轉(zhuǎn)運(yùn)過(guò)程中與細(xì)胞屏障發(fā)生摩擦、阻塞等一系列相互作用,從而引起細(xì)胞壁、細(xì)胞膜組分破損以及胞間連絲等細(xì)胞通道的堵塞[88]。進(jìn)入細(xì)胞內(nèi)部的MNMs還可能與納米及微米尺寸的細(xì)胞結(jié)構(gòu)發(fā)生相互作用,這也被看作是納米顆粒產(chǎn)生毒性效應(yīng)的一種方式[89],通常這種相互作用會(huì)干擾膜表面的電子傳遞,造成線(xiàn)粒體和葉綠體等細(xì)胞器結(jié)構(gòu)損傷,直接影響植物能量及光合代謝,嚴(yán)重時(shí)甚至?xí)斐杉?xì)胞死亡[90]。(2)MNMs釋放的金屬離子的化學(xué)作用(氧化還原活性、催化活性等)。有研究發(fā)現(xiàn),MNMs與其對(duì)應(yīng)的金屬離子在相同暴露條件下誘導(dǎo)的植物轉(zhuǎn)錄組基因表達(dá)譜十分相似,這表明MNMs釋放的金屬離子引發(fā)的生物效應(yīng)是納米材料毒性的主要來(lái)源之一[42-44]。組成MNMs核心金屬的類(lèi)型可以影響MNMs被植物攝取的方式、在細(xì)胞內(nèi)作用的靶標(biāo)等關(guān)鍵因素,從而對(duì)MNMs的最終毒性起到?jīng)Q定性作用[91]。例如Fe2+會(huì)與H2O2反應(yīng)生成高反應(yīng)活性的HO·,從而導(dǎo)致細(xì)胞內(nèi)的有機(jī)大分子發(fā)生氧化損傷;Ag+會(huì)與蛋白質(zhì)的巰基結(jié)合導(dǎo)致蛋白質(zhì)變性失活;某些金屬離子會(huì)競(jìng)爭(zhēng)性替代植物體某些成分的核心金屬原子,破壞植物細(xì)胞金屬離子穩(wěn)態(tài),例如Ag+可以通過(guò)競(jìng)爭(zhēng)性替代質(zhì)體藍(lán)素中的Cu2+來(lái)影響光合作用[92],ZnONPs釋放的Zn2+可以替代葉綠素中心的Mg2+,導(dǎo)致光合核心的改變,進(jìn)而導(dǎo)致植物毒性[93]。(3)氧化脅迫損傷。這是目前普遍接受的MNMs產(chǎn)生植物毒性的主要機(jī)制之一,大多數(shù)研究也發(fā)現(xiàn)MNMs暴露后植物體抗氧化酶酶活以及相關(guān)編碼基因在轉(zhuǎn)錄表達(dá)水平的變化,這些皆已表明MNMs暴露可誘導(dǎo)植物體抗氧化防御系統(tǒng)的響應(yīng)。化學(xué)合成的MNMs表面通常殘留有還原基團(tuán),環(huán)境中氧原子或羥基的存在會(huì)使MNMs具有表面電荷,并且由于小尺寸而增大的比表面積增加了MNMs與細(xì)胞內(nèi)各種物質(zhì)反應(yīng)的幾率,這些因素都決定了MNMs具有極強(qiáng)的表面活性[94-95],在接觸植物細(xì)胞后,高反應(yīng)性的MNMs會(huì)誘導(dǎo)ROS產(chǎn)生,引發(fā)植物氧化脅迫。植物體內(nèi)存在一套復(fù)雜的抗氧化防御系統(tǒng),用于維持體內(nèi)ROS的動(dòng)態(tài)平衡,當(dāng)氧化還原穩(wěn)態(tài)失衡時(shí),ROS會(huì)進(jìn)一步造成蛋白質(zhì)、DNA、生物膜及其他組分等發(fā)生氧化損傷,并干擾細(xì)胞信號(hào)轉(zhuǎn)導(dǎo)[96-97]。此外,大量積累的ROS也會(huì)通過(guò)阻礙葉綠素合成及加速葉綠素降解兩種途徑最終導(dǎo)致植物葉綠素含量下降,從而對(duì)植物光合作用產(chǎn)生負(fù)面效應(yīng)[98]。(4)影響植物基因復(fù)制、轉(zhuǎn)錄及蛋白質(zhì)合成,干擾植物碳氮代謝。MNMs脅迫下植物體與DNA復(fù)制(組蛋白、核小體組裝等)相關(guān)基因在轉(zhuǎn)錄水平的下調(diào),蛋白水平上協(xié)助蛋白質(zhì)折疊與易位(熱休克蛋白、伴侶蛋白),調(diào)控轉(zhuǎn)錄及翻譯(轉(zhuǎn)錄因子、翻譯控制蛋白、翻譯起始因子等)的蛋白水平的波動(dòng),這些都印證了MNMs對(duì)植物體轉(zhuǎn)錄及翻譯過(guò)程的干擾。基于轉(zhuǎn)錄表達(dá)水平層次,1 mg·L-1Cu NPs水培暴露下的小麥[47]和500 mg·L-1ZnONPs水培暴露下的玉米[45]體內(nèi)與氮代謝有關(guān)基因的上調(diào),MNMs混合土壤暴露(1400 mg·kg-1ZnO NPs、180 mg·kg-1Ag NPs和5000 mg·kg-1TiO2NPs,老化6個(gè)月后)下蒺藜苜蓿根部與結(jié)瘤固氮和氮代謝相關(guān)的基因表達(dá)的下調(diào)[48],以及代謝水平層次上,Ag NPs(10、100 mg·L-1,葉面噴施)[78]、Cu NPs(10、20 mg·L-1,水培暴露[81];200、400、800 mg·kg-1,土壤暴露[83])均導(dǎo)致黃瓜體內(nèi)氨基酸組成發(fā)生變化,這些都表明MNMs對(duì)于植物體氮代謝過(guò)程產(chǎn)生了影響。蛋白質(zhì)組學(xué)與代謝組學(xué)研究還顯示碳代謝(光系統(tǒng)、三羧酸循環(huán)和糖酵解)相關(guān)的蛋白與其代謝物水平也會(huì)受到MNMs暴露脅迫的顯著干擾[64,82]。
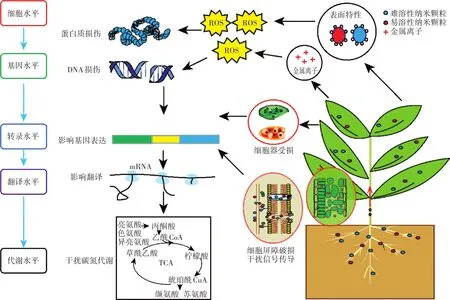
圖1 MNMs與植物的相互作用Figure 1 Schematic representation of MNMs and plants interactions
4 存在問(wèn)題及前景
近年來(lái),越來(lái)越多的學(xué)者將組學(xué)手段應(yīng)用于MNMs植物毒理研究中,現(xiàn)有的研究結(jié)果已經(jīng)發(fā)掘了部分植物響應(yīng)MNMs的差異基因、差異蛋白以及差異代謝物等,并預(yù)測(cè)了部分相關(guān)代謝通路,為我們后續(xù)進(jìn)一步研究MNMs植物毒性的分子機(jī)理提供了新的思路,就目前研究來(lái)看,仍有以下幾個(gè)問(wèn)題亟待解決:
(1)目前大部分研究?jī)H局限使用單一組學(xué)技術(shù),然而生物體代謝通路及調(diào)控錯(cuò)綜復(fù)雜,且基因、蛋白與代謝物的表達(dá)具有時(shí)空差異,單一組學(xué)數(shù)據(jù)通常具有一定的片面性,因而僅依靠一種組學(xué)研究手段難以獲取完整的生物學(xué)信息,未來(lái)開(kāi)展MNMs植物毒性?xún)?nèi)在分子機(jī)制的探究,仍需要采用多種組學(xué)整合手段。此外,如何將龐大的多組學(xué)數(shù)據(jù)與植物表型數(shù)據(jù)進(jìn)行科學(xué)的整合分析,建立植物基因、蛋白及內(nèi)源代謝物與表型變化之間的聯(lián)系也是未來(lái)研究的重點(diǎn)。
(2)當(dāng)前大多數(shù)研究采用水培或砂土作為培養(yǎng)基質(zhì)而非真實(shí)土壤,培養(yǎng)條件多是受控的實(shí)驗(yàn)室或溫室而非真正的自然條件,且研究多集中于MNMs高濃度急性短期暴露實(shí)驗(yàn),考慮到實(shí)際存在土壤中MNMs的現(xiàn)實(shí)濃度,并且由于MNMs進(jìn)入土壤中會(huì)發(fā)生溶解、聚集、吸附等一系列化學(xué)反應(yīng),目前亟需開(kāi)展更貼近實(shí)際環(huán)境中MNMs的低濃度長(zhǎng)期土壤暴露實(shí)驗(yàn),以獲取更加科學(xué)有效的數(shù)據(jù)。
(3)MNMs的植物生物效應(yīng)還取決于以下幾個(gè)因素:MNMs本身特有的性質(zhì)(MNMs種類(lèi)、形狀、粒徑、表面結(jié)構(gòu)以及穩(wěn)定性與溶解性等理化特征),植物生理因素(如植物的不同種類(lèi)、植株不同生長(zhǎng)階段和不同組織器官響應(yīng)與耐受程度各異),實(shí)驗(yàn)設(shè)計(jì)因素(如MNMs的施用方式、暴露濃度、暴露基質(zhì)和暴露時(shí)間等差異)。但目前絕大多數(shù)研究在實(shí)驗(yàn)所用MNMs的理化性質(zhì)、暴露濃度、暴露基質(zhì)、供試植物種類(lèi)及生長(zhǎng)階段等方面難以達(dá)到統(tǒng)一,導(dǎo)致大量實(shí)驗(yàn)結(jié)果不具可比性,需要對(duì)MNMs的來(lái)源、制備方法、供試植物的選擇以及暴露條件的設(shè)計(jì)等進(jìn)行規(guī)范化,建立一套標(biāo)準(zhǔn)系統(tǒng)的MNMs植物毒理研究方法。
(4)植物-土壤-微生物是一個(gè)不可分割的有機(jī)整體,MNMs對(duì)于植物的影響并不是孤立的。土壤提供植物與微生物生活的物質(zhì)基礎(chǔ),土壤微生物又促進(jìn)土壤養(yǎng)分的循環(huán)與轉(zhuǎn)化,并通過(guò)氮素固定、影響根發(fā)育以及抑制植物病原菌等一系列途徑,直接或間接地影響植物的生長(zhǎng),而植物通過(guò)產(chǎn)生根系分泌物選擇與調(diào)節(jié)根際微生物的數(shù)量種類(lèi)和定殖能力。關(guān)注MNMs暴露下植物應(yīng)激表現(xiàn)與土壤養(yǎng)分功能以及土壤微生物群落變化響應(yīng)之間的內(nèi)在耦聯(lián)作用,對(duì)于準(zhǔn)確表征真實(shí)環(huán)境中MNMs植物效應(yīng)具有現(xiàn)實(shí)意義。
猜你喜歡
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天津外國(guó)語(yǔ)大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
汽車(chē)工程學(xué)報(bào)(2017年2期)2017-07-05 08:13:02
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
