小麥RCAR基因家族的鑒定、進化與逆境響應(yīng)
2020-03-15 03:31:38史夢夢溫思鈺趙佳佳喬玲武棒棒鄭興衛(wèi)鄭軍
中國農(nóng)業(yè)科技導(dǎo)報 2020年8期
史夢夢, 溫思鈺, 趙佳佳, 喬玲, 武棒棒, 鄭興衛(wèi),*, 鄭軍,*
(1. 山西大學(xué)生物工程學(xué)院,太原030006;2.山西農(nóng)業(yè)大學(xué)小麥研究所,山西 臨汾 041000)
脫落酸(abscisic acid,ABA)是調(diào)節(jié)植物生長發(fā)育的重要激素,參與植物的胚胎發(fā)生、種子發(fā)育、側(cè)根發(fā)生、葉片衰老和氣孔關(guān)閉等過程,是植物響應(yīng)干旱脅迫的關(guān)鍵信號分子[1],而ABA受體是ABA信號通路領(lǐng)域研究的核心。2009年,中國、美國、日本和歐洲的研究小組幾乎同時證實了RCAR(regulatory components of ABA receptor)家族是ABA的直接受體[2-3],是ABA調(diào)控的核心元件。RCAR(也稱PLY)家族成員具有RCAR-like結(jié)構(gòu)域,屬于結(jié)合疏水配體保守結(jié)構(gòu)域SRPBCC(START/RHO-α-C/PITP/Bet-v1/CoxG/CalC)家族,通過深層疏水配體結(jié)合囊與ABA結(jié)合。逆境脅迫下ABA與RCAR結(jié)合后抑制下游蛋白磷酸酶(PP2C)的活性,從而恢復(fù)蛋白磷酸激酶活性,參與抵御逆境脅迫[4]。
擬南芥中RCAR家族共有14個成員,分別為RCAR1~RCAR14,除了AtRCAR7以外,可在植物體內(nèi)激活A(yù)BA下游基因的表達[5-9],提高植物抵御逆境脅迫的能力,不同成員間表達水平不同,功能上存在冗余。AtRCAR5過表達能顯著提高種子萌發(fā)、幼苗生長和氣孔開閉等過程中對ABA 的敏感性,還能提高抗旱能力[10-11];AtRCAR1過表達誘導(dǎo)葉片提前衰老,使水分流向發(fā)育中組織,顯著提高擬南芥抗旱能力[12];同時敲除AtRCAR1和AtRCAR3使側(cè)根生長靜止期延長,與僅敲除AtRCAR3相比,初生根和側(cè)根都對ABA的敏感性降低[13]。可以看出,RCAR基因家族可顯著提高擬南芥的抗旱能力,且不同成員參與調(diào)控機制存在差異。
水稻中RCAR基因也參與逆境脅迫調(diào)控,多數(shù)研究主要集中在旱脅迫方面。Kim等[14-15]從水稻基因組中篩選出13個RCARs,其中OsRCAR5能提高種子萌發(fā)和幼苗階段對ABA的敏感性,增強成株期抗旱和抗鹽堿能力。OsRCAR3和OsRCAR9也參與水稻的非生物脅迫,過表達能夠提高水稻耐旱性和耐冷性[16]。此外,玉米中有13個RCARs,ZmRCAR3、ZmRCAR9、ZmRCAR10和ZmRCAR13的過表達均可提高ABA敏感性,增加脯氨酸積累,進一步證實RCARs在旱脅迫中具有重要作用[17-19]。小麥?zhǔn)侵匾募Z食作物,非生物脅迫是小麥減產(chǎn)的主要因素,而小麥?zhǔn)钱愒戳扼w,抵御非生物脅迫的調(diào)控機制較為復(fù)雜[20],隨著高質(zhì)量基因組序列的公布,抗逆相關(guān)基因資源的挖掘亟待開展。本研究從小麥基因組中分離RCAR家族基因,分析了家族成員序列、表達以及進化特性,以期為闡明小麥RCAR基因家族在非生物脅迫中的調(diào)控機制奠定基礎(chǔ)。
1 材料與方法
1.1 數(shù)據(jù)來源
擬南芥、水稻的RCARs基因和蛋白序列信息參考TAIR數(shù)據(jù)庫(https://www.arabidopsis.org/index.jsp)和國家水稻數(shù)據(jù)中心(http://www.ricedata.cn/gene/index.htm),玉米和大麥的基因組數(shù)據(jù)參考JGI數(shù)據(jù)庫(https://phytozome.jgi.doe.gov/pz/portal.html),六倍體小麥基因組序列信息來源于IWGSC數(shù)據(jù)庫(http://www.wheatgenome.org/)。
1.2 小麥RCAR基因的鑒定
利用擬南芥和水稻RCAR基因序列對小麥全基因組進行本地Blast,設(shè)置E-value<1e-5,初步篩選出小麥RCAR家族候選基因。Pfam數(shù)據(jù)庫(http://pfam.xfam.org)和SMART數(shù)據(jù)庫(http://smart.embl-heidelberg.de/)驗證蛋白保守結(jié)構(gòu)域。利用ExPASy ProtParam(https://web.expasy.org/protparam/)分析小麥RCAR蛋白的分子量、等電點和疏水性等理化性質(zhì),CELLO(http://cello.life.nctu.edu.tw/)進行亞細(xì)胞定位預(yù)測。
1.3 基因系統(tǒng)進化分析
利用MEGA6.0軟件的MUSCLE默認(rèn)參數(shù)設(shè)置生成多序列比對,使用MEGA6.0軟件最大似然法(maximum likelihood,ML),bootstrap number設(shè)為1 000次重復(fù),構(gòu)建系統(tǒng)發(fā)育樹。
1.4 基因結(jié)構(gòu)和蛋白質(zhì)序列分析
GSDS2.0(http://gsds.cbi.pku.edu.cn/)分析TaRCARs基因結(jié)構(gòu),MEME Suite 4.12.0(http://meme-suite.org/index.html)搜索蛋白質(zhì)序列保守基序,最大保守基序數(shù)目設(shè)為10,每一基序長度設(shè)為6~300個氨基酸殘基。
1.5 啟動子順式作用元件分析
截取起始密碼子上游3 000 bp的基因組序列作為啟動子區(qū),利用PlantCARE(http://bioinformatics.pAet.ugent.be/webtools/plantcare/html)數(shù)據(jù)庫分析順式作用元件。
1.6 基因家族的表達模式分析
從expVIP數(shù)據(jù)庫檢索TaRCARs在14個組織器官中的表達數(shù)據(jù),相關(guān)表達數(shù)據(jù)進行l(wèi)og2轉(zhuǎn)換處理,分析家族成員的表達模式。
1.7 基因進化和正選擇檢測
在構(gòu)建進化樹的基礎(chǔ)上,通過PAML軟件利用樹枝模型法(branch model)分析系統(tǒng)發(fā)育樹中每個Group的進化速率,似然比測驗(LRTs)用于檢測無效模型和備擇模型;在檢測選擇位點時,利用BEB(bayes empirical bayes)方法計算每個位點在不同選擇模型的后驗概率。
1.8 基因表達檢測
挑選完整無損、大小均勻一致的“中國春”種子(本實驗室保存),將露白種子種于育苗盤,在光溫培養(yǎng)箱中16 h光照/8 h黑暗,23 ℃條件下生長一周,進行脅迫(30%PEG和42 ℃)處理。處理0、6、12、24和48 h取根、莖和葉組織,液氮速凍,-80 ℃ 保存。RNA提取和cDNA的制備參考Zhao等[12]方法。不同組織樣品RNA提取后,采用SpectraMax QuickDrop 超微量分光光度計(Molecular Devicess, America)測定核酸濃度,取不同組織RNA等濃度混合反轉(zhuǎn)錄。Realtime-PCR按照TaKaRa SYBR PremixExTaq使用說明,在ABI7300 Realtime PCR儀(Thermo Fisher, America)上運行反應(yīng)程序,甘油醛-3-磷酸脫氫酶基因GAPDH(GenBank登錄號: EF592180.1)作為內(nèi)參,2-ΔΔCT[21]計算目標(biāo)基因的相對表達量。
2 結(jié)果與分析
2.1 TaRCAR基因家族信息
小麥基因組中共檢索到27個RCARs,除第5和6同源群外,其余同源群均有分布。在染色體上分布并不均勻,第1同源群中分布最多,為3個,第3和7同源群中均為2個,第2和4同源群中分布的數(shù)目均為1個,且部分同源群間無缺失,將RCAR基因命名為TaRCAR1-1AS~TaRCAR9-7DL(表1)。TaRCAR分子量為19 177.66~30 169.25 Da之間,等電點差異較大,介于4.97~9.17之間。亞細(xì)胞定位預(yù)測發(fā)現(xiàn)除了TaRCAR8定位于細(xì)胞質(zhì)上和TaRCAR5-3BL定位于細(xì)胞核上外,其余TaRCARs均定位于葉綠體上。此外,TaRCAR1-1AS、TaRCAR4-2DS、TaRCAR5-3DS、TaRCAR7-4AS和TaRCAR9還定位于細(xì)胞核上,TaRCAR3-1AL、TaRCAR3-1BL、TaRCAR4-2AS、TaRCAR4-2BL和TaRCAR6-3DL還定位于線粒體上。TaRCAR6-3BL、TaRCAR7-4BL和TaRCAR7-4DL均是疏水蛋白,其余為親水蛋白。
2.2 TaRCAR蛋白的系統(tǒng)進化分析
選取擬南芥、水稻、玉米、大麥和小麥的RCAR蛋白構(gòu)建系統(tǒng)發(fā)生樹,結(jié)合擬南芥和水稻基因分析發(fā)現(xiàn),RCAR蛋白主要分為6個亞組(圖1),分別為GroupA~F,HvRCAR與TaRCAR親緣關(guān)系最近, 其次為玉米、水稻和擬南芥。其中GroupD成員最多,為22個,含有9個TaRCARs;GroupA、GroupB和GroupF成員分別為14、10和21個,TaRCARs數(shù)目均有6個;GroupC和GroupE成員最少,是擬南芥特有分支,GroupC中AtRCAR8~10較為特殊,均以單體的形式存在,在沒有ABA的情況下能夠抑制PP2Cs[22-23];GroupE分支中的AtRCAR7不依賴ABA就能抑制PP2C家族的成員PP2CA,但不能抑制成員ABI1、HAB1和AHG1[24],推測擬南芥的這6個成員在進化的過程中發(fā)生了功能分化。GroupA和GroupB組中僅包含禾本科植物RCAR蛋白序列,暗示其可能分化于雙子葉植物分化之后。GroupD和GroupF組中同時包含擬南芥和禾本科植物RCAR蛋白,且有2組旁系同源AtRCAR,推測這類基因的分化在單、雙子葉植物分化之前以物種特有的方式進行擴張。
2.3 TaRCAR基因結(jié)構(gòu)與蛋白質(zhì)保守基序分析
27個TaRCARs大多沒有內(nèi)含子,只有TaRCAR1(TaRCAR1-1AS、TaRCAR1-1BS、TaRCAR1-1DS)和TaRCAR8(TaRCAR8-7AL、TaRCAR8-7BL、TaRCAR8-7DL)中有2個內(nèi)含子(圖2)。TaRCARs保守基序個數(shù)為4~6,其保守基序類型及排列順序基本一致,基序1、2、3和4的保守性最高,除TaRCAR3-1BL和TaRCAR6-3BL的基序3外,所有TaRCARs均具有基序1、2、3和4。基序8、9和10的保守性相對較低,基序8只在TaRCAR7-4BL和TaRCAR7-4DL存在,基序9只在TaRCAR3中發(fā)現(xiàn),基序10僅存在于TaRCAR1和TaRCAR5。

表1 小麥RCAR家族成員鑒定及蛋白質(zhì)理化分析Table 1 Identification and protein physic-chemical analysis of the wheat RCAR gene family
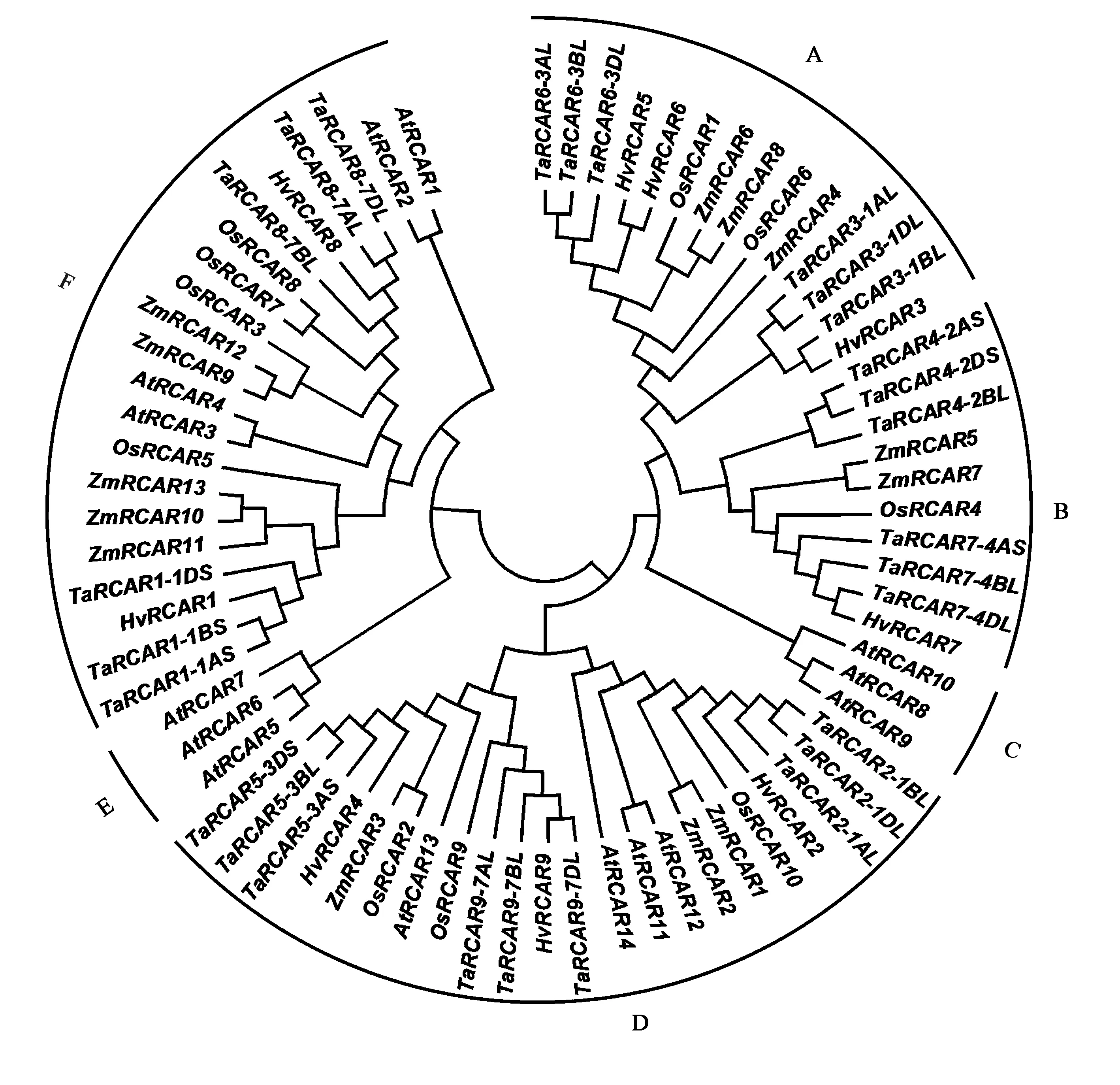
注:At—擬南芥;Os—水稻;Zm—玉米;Hv—大麥;Ta—小麥。Note: At—A. thaliana; Os—O. sativa; Zm—Z. mays; Hv—H. vulgare; Ta—T. aestivum.圖1 不同物種RCAR蛋白進化樹分析Fig.1 Phylogenetic tree of RCAR proteins in different species

Motif 1: REVTVVSGLPATTSTERLEILDDERHVLSFRVVGGEHRLRNYRSVTTVHE (50 aa); Motif 2: HAPGAGQCTSAVVQRIAAPV AAVWSVVRRFDQPQAYKHFVRSCALVAGDG (50 aa); Motif 3: DGPPGTVVVESYVVDVPDGNTEEDTRMFVDTVVKCNLQSLARVAEK LAAA (50 aa); Motif 4: AEVPGEVARHH (11 aa); Motif 5: MPYPASRPSLQQHARIASGSG (21 aa); Motif 6: GGGLGGGGD RIWRPWDEAAVG (21 aa); Motif 7: MEAHMERALREGVTE (15 aa); Motif 8: MPCIPVSSPSIQHHNHNHHHRVLAGVG VGMGCGAEAVVAAAGTAGMRCRE (50 aa); Motif 9: GWKAAAHAASC (11 aa); Motif 9: SPPPPDEQ (8 aa).圖2 TaRCAR基因結(jié)構(gòu)和蛋白質(zhì)保守基序分析Fig.2 Gene structure and conserved motifs of TaRCAR
2.4 TaRCARs表達模式分析
分析TaRCARs在胚芽鞘、根、莖、葉、節(jié)間、旗葉、小穗、雄蕊、雌蕊、穎殼、外稃、芒、籽粒和胚乳組織表達數(shù)據(jù)并繪制熱圖(圖4)。TaRCARs存在兩種表達模式:其中TaRCAR1、TaRCAR2、TaRCAR4和TaRCAR7是組成型表達,TaRCAR3、TaRCAR5、TaRCAR6、TaRCAR8和TaRCAR9為組織特異性表達。TaRCAR1和TaRCAR2在葉、節(jié)間和旗葉中的表達量較高,籽粒發(fā)育中表達量較低;TaRCAR5和TaRCAR6只在根中表達,TaRCAR8僅在籽粒和胚乳中表達。此外,TaRCARs部分同源群間也存在差異,TaRCAR8-7AL和TaRCAR8-7DL在胚乳中表達,而TaRCAR8-7BL在胚乳中未檢測到;TaRCAR9中僅有TaRCAR9-7DL在雌蕊中有表達。TaRCARs表達模式不同,部分同源群間也發(fā)生了分化,暗示TaRCARs功能具有一定的多樣化,在不同生長發(fā)育過程中行使功能。
2.5 TaRCARs的進化速率分析
利用PAML軟件計算了每個亞組的進化速率,研究基因的進化機制是否存在差異。整體上看TaRCARs在進化過程中亞組間進化速率差異較大(表2)。GroupC、GroupD和GroupF進化速率較低,其ω值分別為0.162、0.179和0.000 01,亞組進化速率極小暗示組內(nèi)成員功能相同或相近。GroupF中OsRCAR5(Os05g12260)是水稻中有功能的ABA受體,在種子萌發(fā)和幼苗生長時對ABA極為敏感,過表達可增強水稻的抗旱性和耐鹽性[14],由此推測TaRCAR1和TaRCAR8在小麥響應(yīng)干旱和鹽脅迫的過程中可能發(fā)揮重要作用。GroupA、GroupB和GroupE均出現(xiàn)了快速進化,ω值分別為0.349、0.321和1.280,GroupE是擬南芥特有分支,GroupA和GroupB是禾本科特有分支,進化速率較快。
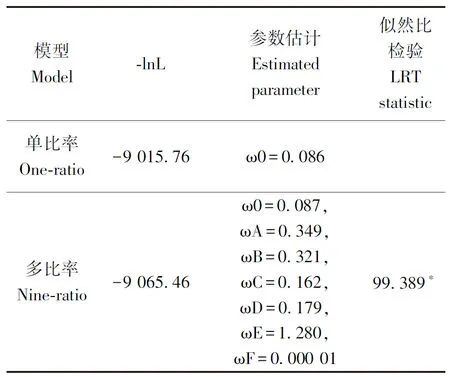
表2 TaRCARs不同亞組的進化速率分析Table 2 Evolutionary rate analysis of TaRCARs
2.6 TaRCARs正選擇檢驗
為了適應(yīng)新的環(huán)境,物種會選擇有利的突變保留并穩(wěn)定遺傳給后代。TaRCARs在GroupA進化速率(ωA=0.349)最快,使用branch-site model檢測GroupA是否存在正選擇位點(表3)。GroupA有3個位點的后驗概率P>0.99,說明相關(guān)位點經(jīng)歷了正選擇。GroupA中OsRCAR1(Os01g61210)的表達在干旱脅迫后顯著上調(diào),而ABA處理后顯著下調(diào)[16];AtRCAR10作為OsRCAR1同源基因,干旱脅迫后表達量降低,過表達能提高擬南芥在種子萌發(fā)、幼苗的生長時期對ABA的敏感[25]。因此,GroupA中TaRCARs出現(xiàn)正選擇,可能導(dǎo)致基因突變產(chǎn)生新的功能,或者喪失原來的功能。
2.7 TaRCARs干旱和熱脅迫下的表達模式
根據(jù)TaRCARs的編碼序列設(shè)計表達引物進行表達特性研究。除TaRCAR8外,成功檢測到其余8個成員的表達特性。在干旱脅迫下,TaRCARs具有不同的表達模式(圖5),TaRCAR1、TaRCAR3、TaRCAR4、TaRCAR5和TaRCAR6在處理6 h達到最高,隨后逐漸降低,趨于正常。而TaRCAR2的表達量先下降,然后恢復(fù)到初始水平。TaRCAR7和TaRCAR9的表達幾乎不受影響。在熱脅迫下,TaRCAR1和TaRCAR2在處理6 h表達量最高,而TaRCAR3、TaRCAR6和TaRCAR9表達量明顯增加,而TaRCAR3、TaRCAR5、TaRCAR6和TaRCAR9在6 h基本無變化,12 h時表達量升高,達到最大,表明不同TaRCARs對熱脅迫響應(yīng)的時間存在差異。TaRCAR7和TaRCAR4的熱處理后分別在6 h和12 h表現(xiàn)出下調(diào)趨勢,說明TaRCARs在響應(yīng)干旱和熱脅迫的機制存在差異。
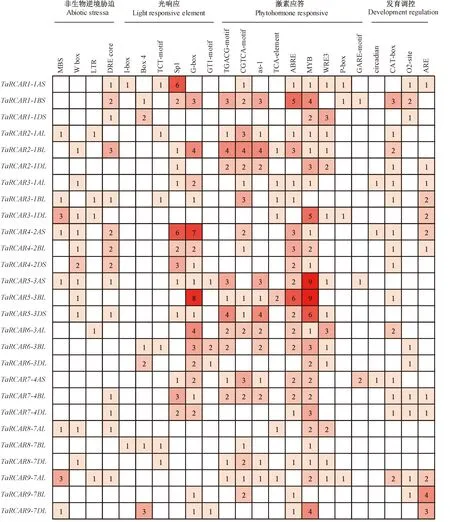
注:數(shù)字代表順式作用元件個數(shù),數(shù)目越多,標(biāo)注的顏色越深。Note: Numbers in the table represent the number of cis-acting elements, the greater the number, the darker the color of the label.圖3 TaRCARs順式作用元件和功能Fig.3 TaRCARs cis-acting elements and functions
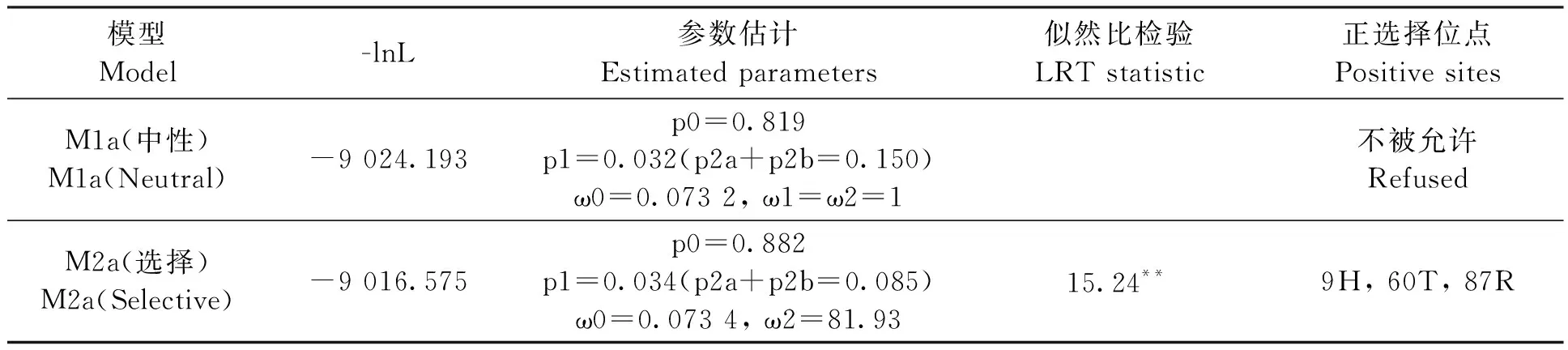
表3 GroupA正選擇位點的檢測Table 3 Positive selection detection of GroupA

圖4 TaRCAR基因表達譜Fig.4 Expression profile of RCAR genes in wheat
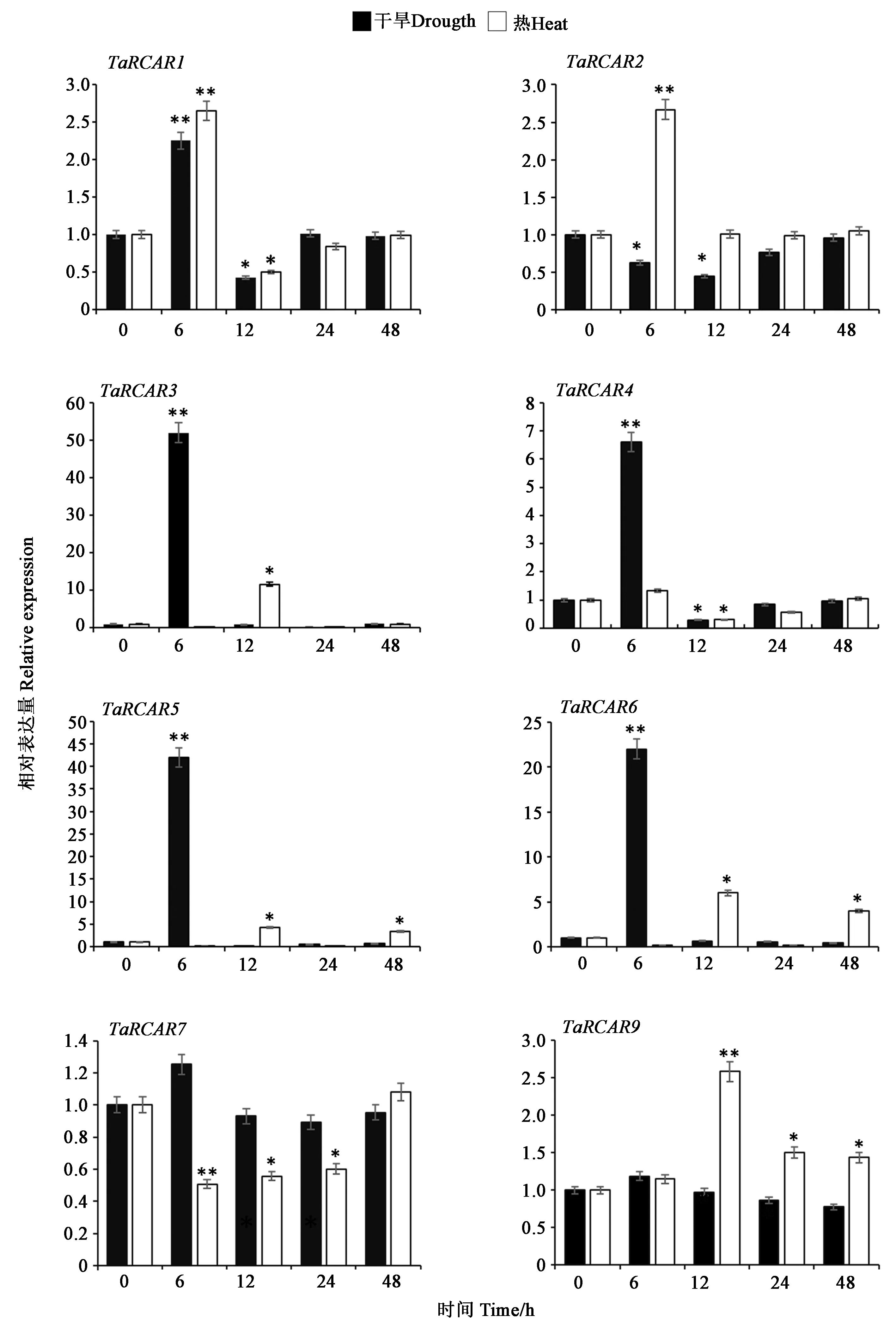
注:t-test分析基因在不同脅迫時間下與對照的差異具有統(tǒng)計學(xué)意義(**: P<0.01;*: P<0.05)。Note: t-test for significant difference between CK and samples(**: P<0.01,*: P<0.05).圖5 小麥RCARs在干旱脅迫和熱脅迫條件下的表達Fig.5 Expression of RCARs under drought stress and heat stress in wheat
3 討論
TaRCARs在部分同源群間分布不均勻,在小麥第5和6同源群中沒有TaRCARs分布,而第1同源群則有9個TaRCARs,第3和7同源群有6個TaRCARs。禾谷類祖先種有5條古染色體,分別為A5、A7、A11、A8和A4,90 MYA (million years ago)時發(fā)生過一次全基因組復(fù)制事件,導(dǎo)致染色體加倍由5條變?yōu)?0條,增加了A1、A10、A12、A9和A6[26-28];隨后兩條染色體發(fā)生斷裂和融合,從而產(chǎn)生兩條新的染色體,分別是由A10和A7組成的A3以及A4和A6組成的A2,導(dǎo)致過渡態(tài)的祖先染色體為12條;在50~70 MYA時Panicoideae亞科與另外兩個亞科分化,Ehrhartoideae亞科約在46 MYA與Pooideae亞科分化[29]。在小麥形成過程中還發(fā)生了一次染色體融合事件,其中A8和A6染色體發(fā)生斷裂重組,形成7號染色體的祖先。除第3、6同源群的祖先染色體未發(fā)生斷裂和重組外,其余同源群在進化過程中基本上都發(fā)生了重組和融合,這可能是導(dǎo)致TaRCARs在部分同源群間發(fā)生偏分布的原因。此外,RCARs參與小麥的適應(yīng)性的進化,在多倍體化過程中拷貝數(shù)變異是重要的進化方式,有些TaRCARs可能發(fā)生了功能性的復(fù)制和刪除。
擬南芥和水稻中RCARs研究較為深入,可以為TaRCARs研究提供參考。物種間同一進化分支的基因互為同源基因或親緣較近,生物學(xué)功能相似。分析發(fā)現(xiàn),擬南芥、水稻、玉米、大麥和小麥的RCAR蛋白進化分析可劃分為6個亞組,GroupD所在分支中包括4個AtRCARs、3個OsRCARs、3個HvRCARs、3個ZmRCARs和9個TaRCARs,其中AtRCAR14(AT2G26040)過表達能夠提高擬南芥種子的萌發(fā)、幼苗形態(tài)建立階段對ABA敏感度,從而增強抗旱性[30];OsRCAR4作為AtRCAR14的同源基因過表達后明顯提高水稻的耐旱性[16],推測TaRCAR2、TaRCAR5和TaRCAR9具有相同功能。GroupA所在分支的OsRCAR1在干旱和冷脅迫下顯著上調(diào)[31],GroupA中的TaRCAR3和TaRCAR6在干旱和熱脅迫下表達顯著升高,暗示它們參與小麥抵御逆境脅迫。在漫長的進化過程中,結(jié)構(gòu)相似的基因在種內(nèi)或種間有時也發(fā)生功能分化。如CCT基因調(diào)控植物開花[32],然而Yang等[33]發(fā)現(xiàn)在玉米中無轉(zhuǎn)座子插入ZmCCT為抗病等位基因,同時介導(dǎo)玉米對低氮和高鹽的抗性。GroupF分支中OsRCAR5正調(diào)控水稻幼苗對ABA的敏感度,可提高成株期抗鹽堿脅迫能力[14-15],而OsRCAR3過表達卻能提高水稻的耐冷性[16]。可見RCARs在物種間雖具有較高的保守性,在進化過程中也會發(fā)生功能分化以適應(yīng)環(huán)境變化。
正選擇是物種進化的動力,是物種適應(yīng)環(huán)境的遺傳基礎(chǔ)。在微生物中,太古菌有一些基因的進化速率加快,而加速進化的基因使太古菌能更好的適應(yīng)地球降溫,證實基因加速進化提高了物種適應(yīng)環(huán)境的能力[34]。在植物中,黃瓜、楊樹、大豆和擬南芥的11S球蛋白在進化過程中受到的正選擇效應(yīng)并不一致,在大豆中顯著高于其他物種,進一步研究證實大豆中11S球蛋白快速進化促進了大豆合成蛋白的能力[35]。TaRCARs在GroupA出現(xiàn)了快速進化的現(xiàn)象,且TaRCAR3和TaRCAR6受到正選擇,可能是小麥適應(yīng)不同逆境脅迫,保留了較多的有利變異所致。小麥自北緯18~50°,從平原到高海拔地區(qū)均有種植,在擴大種植范圍的馴化過程中就要求小麥能夠抵御更多的逆境脅迫。RCAR基因是ABA信號通路中的重要調(diào)控因子,參與植物對逆境的抵御,相關(guān)成員必然受到選擇,而差異選擇為小麥適應(yīng)性提供了遺傳基礎(chǔ)。因此,對TaRCARs進行更深入的研究,有助于解析小麥抗逆調(diào)控和馴化的分子機制。
猜你喜歡
青少年科技博覽(中學(xué)版)(2022年6期)2022-12-27 19:44:27
今日農(nóng)業(yè)(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農(nóng)業(yè)(2021年14期)2021-10-14 08:35:40
民用飛機設(shè)計與研究(2020年4期)2021-01-21 09:15:02
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
中國中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年11期)2014-08-08 13:23:44
