Fe2+和Fe3+對(duì)長(zhǎng)莖葡萄蕨藻生長(zhǎng)、鐵含量和生理特性的影響
2020-03-17 13:05:48王曉梁劉東超徐日升陳海昇
廣東海洋大學(xué)學(xué)報(bào) 2020年2期
王曉梁,劉東超,徐日升,陳海昇
Fe2+和Fe3+對(duì)長(zhǎng)莖葡萄蕨藻生長(zhǎng)、鐵含量和生理特性的影響
王曉梁1,2,劉東超1,2,徐日升1,2,陳海昇1,2
(1. 廣東海洋大學(xué)水產(chǎn)學(xué)院,廣東 湛江 524088;2. 廣東省藻類養(yǎng)殖及應(yīng)用工程技術(shù)中心,廣東 湛江 524025)
【】探索兩種無(wú)機(jī)鐵對(duì)長(zhǎng)莖葡萄蕨藻()的生長(zhǎng)、鐵含量及生理特性的影響。分別以三氯化鐵(FeCl3)和硫酸亞鐵(FeSO4)作為鐵源,采用單因素實(shí)驗(yàn)方法,研究在不同濃度Fe2+、Fe3+條件下,長(zhǎng)莖葡萄蕨藻的生長(zhǎng)、光合色素含量、鐵含量、兩種過(guò)氧化酶(CAT、T-SOD)活性,以及總蛋白、維生素C和可溶性糖含量。在水體中無(wú)論是添加Fe2+還是Fe3+,藻體的特定生長(zhǎng)率(SGR)、光合色素含量、鐵含量、過(guò)氧化物酶活性和維生素C含量均顯著高于對(duì)照組(< 0.05)。藻體的鐵含量隨著培養(yǎng)液中鐵離子濃度的升高而增加,以Fe2+和Fe3+處理后,藻體鐵最高值分別可達(dá)到1 522.54 μg/g和1 373.93 μg/g,增幅為1 369.34%和1 225.92%;藻體維生素C最高質(zhì)量分?jǐn)?shù)分別可達(dá)15.41 μg/mg和12.89 μg/mg,增幅為107.56%和73.58%。Fe2+實(shí)驗(yàn)組對(duì)藻體的可溶性糖含量有顯著影響(< 0.05)。實(shí)驗(yàn)藻體的總蛋白含量顯著低于對(duì)照組(< 0.05)。藻體在3 mg/L Fe2+或者6 mg/L Fe3+的條件下,長(zhǎng)莖葡萄蕨藻生長(zhǎng)最佳,其營(yíng)養(yǎng)品質(zhì)可得以改善。
長(zhǎng)莖葡萄蕨藻;生長(zhǎng);鐵含量;抗氧化酶;營(yíng)養(yǎng)成分
長(zhǎng)莖葡萄蕨藻(),俗稱海葡萄,是一種大型食用綠藻,主要生長(zhǎng)在潮間帶海區(qū)、水流較緩的沙地或礁石[1]。它不僅富含多種人體所需的氨基酸、維生素和礦物質(zhì)等營(yíng)養(yǎng)元素[2-4],還含有多種活性成分,在治療痛風(fēng)、抗病毒與癌癥等方面具有較高的醫(yī)藥價(jià)值[5-7]。
鐵是人體的一種必需微量元素,主要在細(xì)胞代謝和氧的轉(zhuǎn)運(yùn)中發(fā)揮重要生理作用[8-9]。人體所需的鐵主要通過(guò)食物鏈獲取,而自然食物中鐵含量通常不能滿足人體需要。據(jù)統(tǒng)計(jì),缺鐵性貧血依舊是一個(gè)世界性的公共健康問(wèn)題[10],有必要在食物中補(bǔ)充鐵來(lái)滿足人體的生理需要,以蒸煮谷物植物為主食的人群更需補(bǔ)鐵[11-12]。目前增加膳食中鐵供給途徑主要有直接補(bǔ)充、食物強(qiáng)化、飲食多樣化、作物生物強(qiáng)化(包括育種與施肥)[11]。許多學(xué)者根據(jù)作物生物強(qiáng)化的原理開(kāi)展了許多經(jīng)濟(jì)作物的富鐵研究,而作為功能食品的藻類的消費(fèi)不斷增加[13],因此研究開(kāi)發(fā)富鐵的藻類功能食品有重要價(jià)值。目前,關(guān)于食用藻類的富鐵研究?jī)H見(jiàn)鈍頂螺旋藻()和小球藻()等微藻的報(bào)道[14-15],而大型食用海藻類富鐵研究鮮有報(bào)道。
長(zhǎng)莖葡萄蕨藻為公眾喜愛(ài)的一種高端生鮮美食。筆者研究Fe2+、Fe3+兩種無(wú)機(jī)鐵對(duì)長(zhǎng)莖葡萄蕨藻的生長(zhǎng)、富鐵量和生理特性的影響,旨在為生產(chǎn)富鐵的長(zhǎng)莖葡萄蕨藻,開(kāi)發(fā)新型長(zhǎng)莖葡萄蕨藻功能食品提供參考。
1 材料與方法
1.1 長(zhǎng)莖葡萄蕨藻藻種
實(shí)驗(yàn)用長(zhǎng)莖葡萄蕨藻藻種購(gòu)自深圳市藍(lán)汀鼎執(zhí)生物科技有限公司,培養(yǎng)于廣東海洋大學(xué)水產(chǎn)學(xué)院藻類實(shí)驗(yàn)室。
暫養(yǎng)條件:溫度(26±1)℃,照度(2 000±100)lx,光暗周期12 L:12 D,營(yíng)養(yǎng)鹽為0.5 mmol/L 的NaNO3溶液和0.1 mmol/L的KH2PO4溶液,海水鹽度30±1,連續(xù)充氣培養(yǎng),換水周期為7 d[16]。
1.2 富鐵實(shí)驗(yàn)設(shè)計(jì)
挑選長(zhǎng)勢(shì)良好的長(zhǎng)莖葡萄蕨藻,剪取25 mm左右的直立枝作為實(shí)驗(yàn)對(duì)象,用干凈海水反復(fù)沖洗后,將直立枝放入滅菌不加營(yíng)養(yǎng)鹽的海水中進(jìn)行適應(yīng)培養(yǎng)1 d。
實(shí)驗(yàn)用鐵化合物為1 g/L FeCl3溶液、1 g/L FeSO4溶液。全部藥品均為分析純(AR)。
采用1.5 L的方形開(kāi)口塑料盒作為培養(yǎng)盒,添加1 L過(guò)濾滅菌自然海水,分別加入1 mL的NaNO3溶液與1 mL的KH2PO4溶液作為營(yíng)養(yǎng)鹽。根據(jù)預(yù)實(shí)驗(yàn)結(jié)果,2種價(jià)態(tài)(Fe2+和Fe3+)的鐵濃度分別設(shè)置為0(對(duì)照組)、3、6、9、12 mg/L。選取暫養(yǎng)后健康的直立枝,稱取質(zhì)量(0),隨機(jī)分至各組,每組放置5棵直立枝,每個(gè)實(shí)驗(yàn)組設(shè)置3個(gè)平行組。
培養(yǎng)條件:不充氣,其他條件與暫養(yǎng)條件一致。連續(xù)培養(yǎng)7 d,每天觀測(cè)生長(zhǎng)狀況,7 d后取出。
1.3 指標(biāo)測(cè)定
對(duì)培養(yǎng)結(jié)束后的藻體進(jìn)行充分沖洗,去除表面的鐵溶液,用吸水紙吸干表面水分,稱取質(zhì)量(m),計(jì)算藻體的特定生長(zhǎng)率(SGR)[16]。
將藻體烘干,研磨制成藻粉,進(jìn)行硝化,用電感耦合等離子體質(zhì)譜儀(ICP-MS)測(cè)其含鐵量(Fe)[17]。
按照南京建成生物工程研究所的試劑盒說(shuō)明書(shū)要求,將藻體進(jìn)行剪切、勻漿、稀釋等,測(cè)其光合色素含量、過(guò)氧化氫酶(CAT)活力、總超氧化物歧化酶(T-SOD)活力、總蛋白(TP)含量、維生素C(VC)含量和可溶性糖含量。
1.4 數(shù)據(jù)的處理和分析
SGR = [100 ln(m/0)] /;
Fe= (待測(cè)樣濃度-空白濃度)測(cè)/測(cè);
(Chl-a)=[13.95(665nm)-6.88(649nm)]測(cè)/測(cè);
(Chl-b)=[24.96(649nm)-7.32(665nm)]測(cè)/測(cè);
(類胡蘿卜素) = {[1 000(470nm) - 2.05×(Chl-a) - 114.8(Chl-b)] / 245}測(cè)/測(cè)。
式中,SGR為特定生長(zhǎng)率(%·d-1),0為初始藻體質(zhì)量(g),m為最終藻體質(zhì)量(g);測(cè)為待測(cè)樣硝化處理后的體積(L),測(cè)為待測(cè)樣的質(zhì)量(g)。
總蛋白質(zhì)量濃度= (待測(cè)OD值 -空白OD值) / (標(biāo)準(zhǔn)OD值-空白OD值) × 標(biāo)準(zhǔn)品質(zhì)量濃度(0.524 g/L)× 稀釋倍數(shù),標(biāo)準(zhǔn)品質(zhì)量濃度0.524 g/L,在= 595 nm處測(cè)量。
CAT活力= (對(duì)照OD值-測(cè)定OD值) × 271 × 1 /(60×取樣量)/ 待測(cè)樣本蛋白濃度,在= 405 nm處測(cè)量。
T-SOD活力= (對(duì)照OD值-測(cè)定OD值) / 對(duì)照OD值 / 50% ×(反應(yīng)液總體積 / 取樣量)/ 待測(cè)樣本蛋白濃度,在= 550 nm處測(cè)量。
VC質(zhì)量分?jǐn)?shù)= (待測(cè)OD值-空白OD值) / (標(biāo)準(zhǔn)OD值 -空白OD值) × 標(biāo)準(zhǔn)品質(zhì)量濃度× 稀釋倍數(shù)(4倍) / 待測(cè)樣本蛋白濃度,標(biāo)準(zhǔn)品質(zhì)量濃度6 μg/mL,在= 536 nm處測(cè)量。
可溶性糖質(zhì)量分?jǐn)?shù)= (待測(cè)OD值 -空白OD值) /(標(biāo)準(zhǔn)OD值- 空白OD值)× 標(biāo)準(zhǔn)品質(zhì)量濃度× 稀釋倍數(shù)/ 待測(cè)樣本蛋白濃度,標(biāo)準(zhǔn)品質(zhì)量濃度100 μg/mL,在= 620 nm處測(cè)量。
采用IBM SPSS Statistics 19軟件對(duì)數(shù)據(jù)進(jìn)行處理和分析。通過(guò)單因素方差分析和Duncan多重比較對(duì)數(shù)據(jù)進(jìn)行顯著性檢測(cè)(< 0.05),每組濃度結(jié)果,按照三組均值±標(biāo)準(zhǔn)誤用GraphPad Prism制圖。
2 結(jié)果與分析
2.1 兩種無(wú)機(jī)鐵對(duì)長(zhǎng)莖葡萄蕨藻SGR的影響
兩種無(wú)機(jī)鐵對(duì)長(zhǎng)莖葡萄蕨藻SGR的影響見(jiàn)圖1。圖1可見(jiàn),隨著Fe2+或Fe3+濃度的升高,長(zhǎng)莖葡萄蕨藻的特定生長(zhǎng)率均呈先升后降變化趨勢(shì)。當(dāng)Fe2+質(zhì)量濃度為3 mg/L時(shí),長(zhǎng)莖葡萄蕨藻的特定生長(zhǎng)率最大,為4.670%·d-1,F(xiàn)e3+質(zhì)量濃度為6 mg/L時(shí),藻體SGR最大,為5.627%·d-1,且均與對(duì)照組(2.746 %·d-1)有顯著差異(< 0.05)。
可見(jiàn),在實(shí)驗(yàn)濃度范圍內(nèi),無(wú)論添加Fe2+還是Fe3+,均可促進(jìn)長(zhǎng)莖葡萄蕨藻的生長(zhǎng)。最適添加的Fe2+質(zhì)量濃度為3 mg/L,F(xiàn)e3+質(zhì)量濃度為6 mg/L。
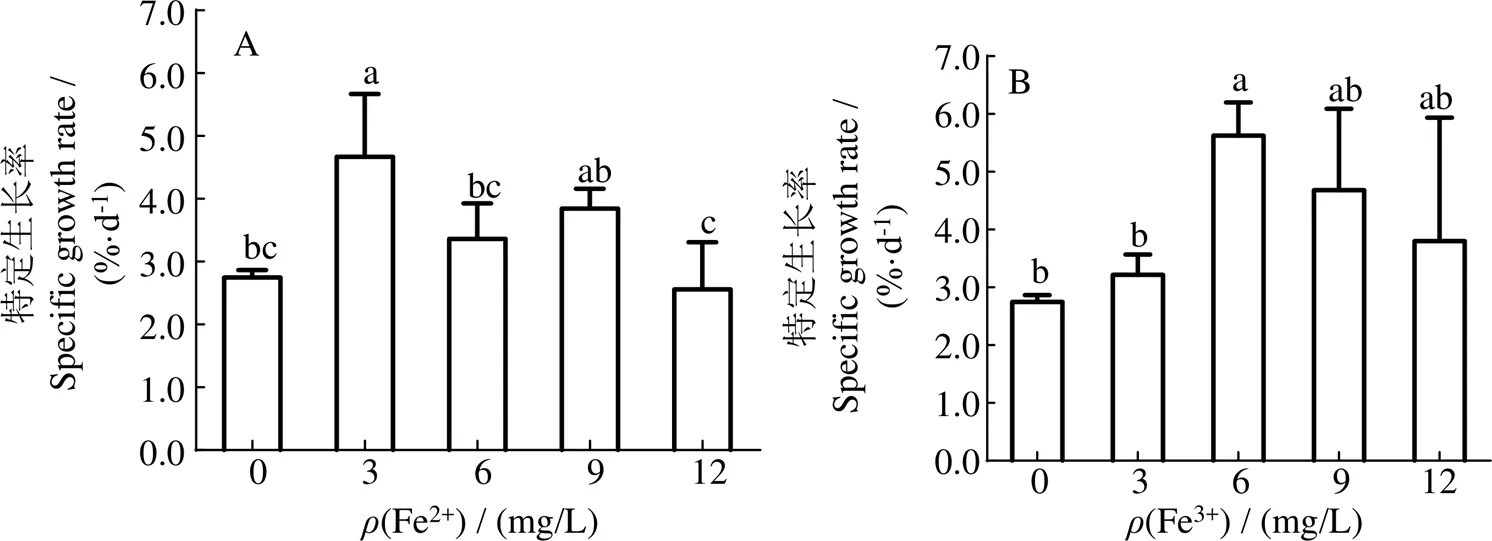
凡含有一個(gè)相同字母則組間差異無(wú)統(tǒng)計(jì)學(xué)意義(P>0.05)
2.2 兩種無(wú)機(jī)鐵對(duì)長(zhǎng)莖葡萄蕨藻光合色素的影響
圖2可見(jiàn),所有加鐵實(shí)驗(yàn)組的藻體葉綠素a含量均高于對(duì)照組,差異有統(tǒng)計(jì)學(xué)意義(< 0.05);鐵濃度變化對(duì)藻體葉綠素b含量影響不顯著(> 0.05);所有加鐵組藻體的類胡蘿卜素含量均值均高于對(duì)照組,但在Fe2+實(shí)驗(yàn)組中僅3 mg/L和6 mg/L兩組差異有統(tǒng)計(jì)學(xué)意義(< 0.05),在Fe3+中,除9 mg/L外,其余3組藻體的類胡蘿卜素含量均與對(duì)照組差異有統(tǒng)計(jì)學(xué)意義(< 0.05)。
可見(jiàn),兩種無(wú)機(jī)鐵主要對(duì)長(zhǎng)莖葡萄蕨藻葉綠素a和類胡蘿卜素的含量變化有顯著影響,對(duì)葉綠素b含量影響不顯著。
2.3 兩種無(wú)機(jī)鐵對(duì)長(zhǎng)莖葡萄蕨藻富鐵量的影響
兩種無(wú)機(jī)鐵對(duì)長(zhǎng)莖葡萄蕨藻富鐵量的影響見(jiàn)表1。表1可見(jiàn),隨著鐵離子濃度的升高,長(zhǎng)莖葡萄蕨藻的富鐵量呈升高趨勢(shì),與對(duì)照組差異有統(tǒng)計(jì)學(xué)意義(< 0.05)。當(dāng)添加Fe2+時(shí),除6 mg/L和9 mg/L兩組之間差異無(wú)統(tǒng)計(jì)學(xué)意義(> 0.05)外,其他各組間差異有統(tǒng)計(jì)學(xué)意義(< 0.05),在12 mg/L時(shí),藻體富鐵量達(dá)到最高值,為1 522.540 μg/g,各組增幅分別為528.80%、901.71%、1 080.21%、1 369.34%。當(dāng)添加Fe3+時(shí),各組間差異有統(tǒng)計(jì)學(xué)意義(< 0.05),在12 mg/L時(shí),藻體富鐵量達(dá)到最高值,為1 373.932 μg/L,各組增幅分別為402.06%、581.89%、903.08%、1225.92%。
綜合分析,在實(shí)驗(yàn)濃度范圍內(nèi),長(zhǎng)莖葡萄蕨藻對(duì)兩種價(jià)態(tài)的鐵均有較強(qiáng)的富集能力。在相同濃度下,F(xiàn)e2+實(shí)驗(yàn)組比Fe3+實(shí)驗(yàn)組的藻體富鐵量略高。

凡含有一個(gè)相同字母則組間差異無(wú)統(tǒng)計(jì)學(xué)意義(>0.05)
The date with a same letter indicate no significant difference between them (> 0.05)
圖2不同的無(wú)機(jī)鐵濃度下長(zhǎng)莖葡萄蕨藻光合色素含量變化
Fig.2 Changes of photosynthetic pigments content inunder different concentrations of inorganic iron
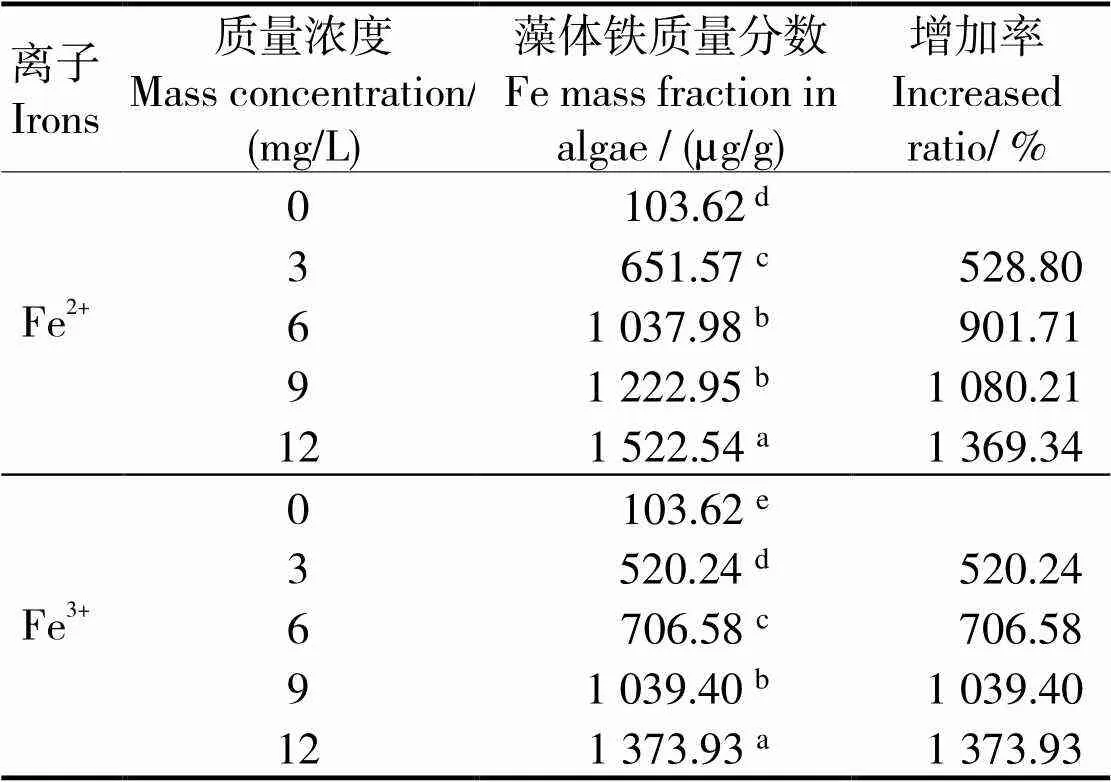
表1 不同的無(wú)機(jī)鐵濃度下長(zhǎng)莖葡萄蕨藻富鐵量
注:凡含有一個(gè)相同字母則組間差異無(wú)統(tǒng)計(jì)學(xué)意義(>0.05)
Note: The date with a same letter indicate no significant difference between them (> 0.05)
2.4 兩種無(wú)機(jī)鐵對(duì)長(zhǎng)莖葡萄蕨藻抗氧化酶CAT、T-SOD活力的影響
兩種無(wú)機(jī)鐵對(duì)長(zhǎng)莖葡萄蕨藻抗氧化酶CAT、T-SOD活力的影響見(jiàn)圖3。圖3可見(jiàn),F(xiàn)e2+處理組的藻體CAT活性顯著高于對(duì)照組(< 0.05)。Fe3+處理組,除3 mg/L組別外,其余處理組藻體的CAT活性顯著高于對(duì)照組(0.05)。在圖3-B中,F(xiàn)e2+處理組的藻體T-SOD活性均與對(duì)照組差異有統(tǒng)計(jì)學(xué)意義(<0.05),F(xiàn)e3+的處理組,除3 mg/L組外,其余處理組藻體T-SOD活性均顯著高于對(duì)照組(<0.05)。可見(jiàn),在實(shí)驗(yàn)濃度范圍內(nèi),除3 mg/L 的Fe3+外,添加兩種無(wú)機(jī)鐵可顯著提高藻體兩種抗氧化酶活力,且當(dāng)添加Fe2+濃度為3 mg/L時(shí),兩種抗氧化酶活力最強(qiáng);當(dāng)Fe3+濃度為6 mg/L時(shí),藻體的兩種抗氧化酶活力開(kāi)始相對(duì)穩(wěn)定。
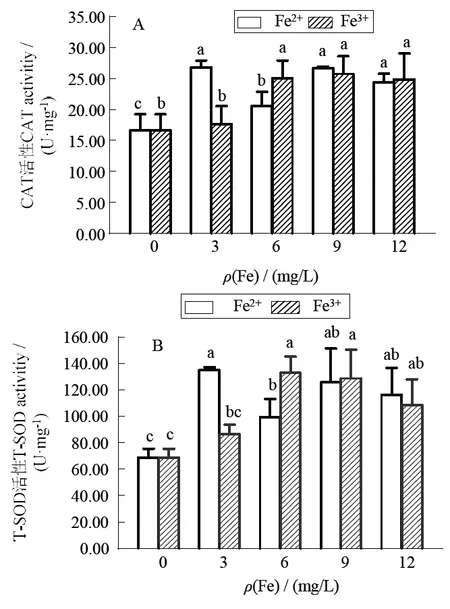
凡含有一個(gè)相同字母則組間差異無(wú)統(tǒng)計(jì)學(xué)意義(>0.05)
The date with a same letter indicate no significant difference between them (> 0.05)
圖3不同無(wú)機(jī)鐵濃度下長(zhǎng)莖葡萄蕨藻過(guò)氧化物酶活力變化
Fig.3 Changes of peroxidase activity inunder different concentrations of inorganic iron
2.5 兩種無(wú)機(jī)鐵對(duì)長(zhǎng)莖葡萄蕨藻總蛋白、VC和可溶性糖含量的影響
兩種無(wú)機(jī)鐵對(duì)長(zhǎng)莖葡萄蕨藻總蛋白、VC和可溶性糖含量的影響見(jiàn)表2。表2可見(jiàn),所有加鐵組的藻體總蛋白含量均顯著低于對(duì)照組(<0.05)。在Fe2+處理后,各實(shí)驗(yàn)組的藻體總蛋白含量比對(duì)照組分別降幅為36.47%、17.88%、31.82%、33.37%。在Fe3+處理后,各實(shí)驗(yàn)組的藻體總蛋白含量分別降幅為18.65%、34.15%、38.80%、30.27%。在Fe2+處理后,添加濃度為3 mg/L和9 mg/L組的VC含量與對(duì)照組相比顯著提高(<0.05),分別增幅107.56 %和128.55 %。而Fe3+處理組中,除3 mg/L組外,其他組均與對(duì)照組有顯著提高(<0.05),分別增幅52.68%、73.58%、71.21%。Fe2+處理后,藻體可溶性糖含量均顯著增加(<0.05),增幅分別為39.01%、5.87%、23.02%、25.12%。Fe3+處理后的藻體可溶性糖含量沒(méi)有顯著提高(>0.05)。
可見(jiàn),F(xiàn)e2+和Fe3+處理組的藻體總蛋白含量均顯著低于對(duì)照組。Fe2+處理后的藻體的VC與可溶性糖含量總體上顯著提高。Fe3+處理組的藻體的VC含量總體上顯著高于對(duì)照組,但藻體的可溶性糖含量與對(duì)照組差異不大。
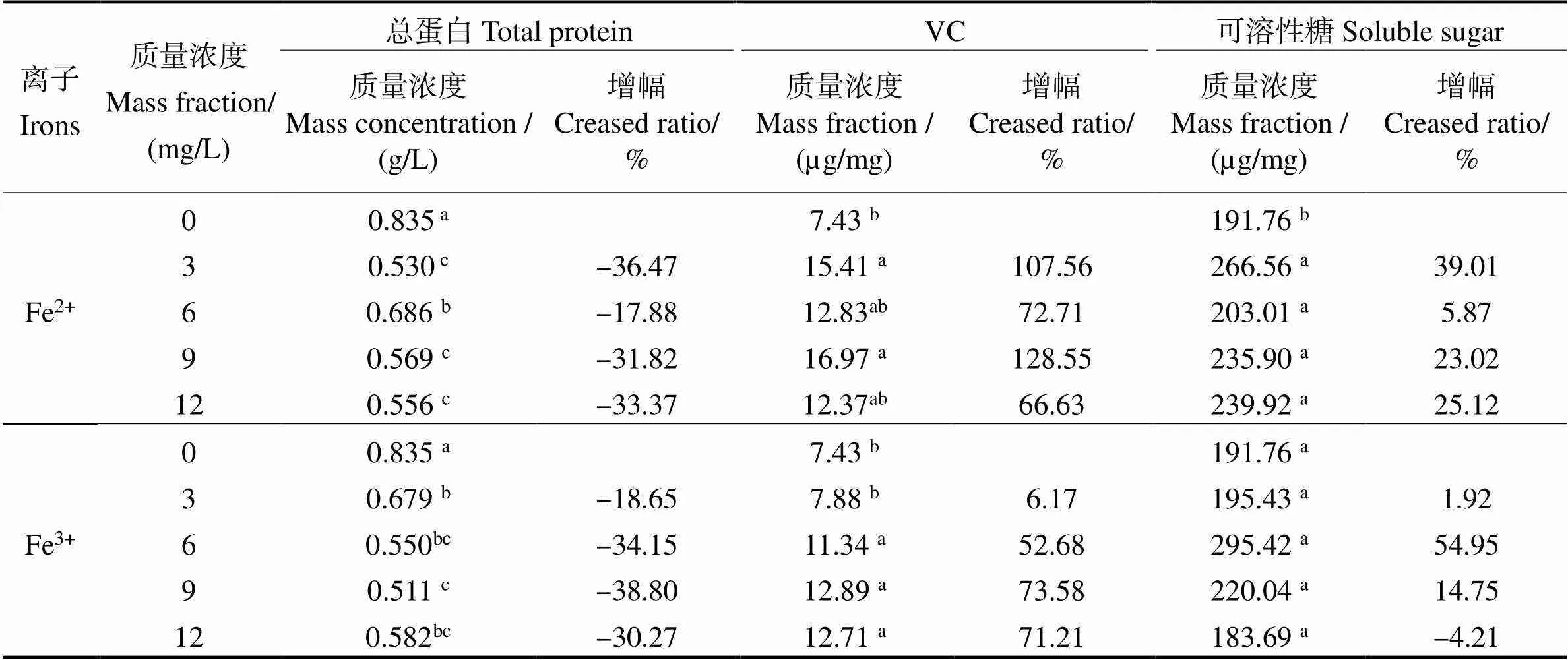
表2 不同的無(wú)機(jī)鐵濃度下長(zhǎng)莖葡萄蕨藻的總蛋白、VC和可溶性糖含量變化
凡含有一個(gè)相同字母則組間差異無(wú)統(tǒng)計(jì)學(xué)意義(>0.05)。
The date with a same letter indicate no significant difference between them (> 0.05).
3 討論
3.1 兩種無(wú)機(jī)鐵對(duì)長(zhǎng)莖葡萄蕨藻生長(zhǎng)和光合色素的影響
鐵是藻類生長(zhǎng)所必需的重要微量元素,同時(shí)也是限制性因素之一,在細(xì)胞呼吸、葉綠素產(chǎn)物合成以及作為輔酶參與電子和氧的轉(zhuǎn)運(yùn)等過(guò)程中發(fā)揮作用[18-21]。在培養(yǎng)液中適當(dāng)增加鐵的濃度可促進(jìn)小球藻()、銅藻()的生長(zhǎng)[15, 22]。本研究表明,無(wú)論是添加Fe2+還是Fe3+,均可促進(jìn)長(zhǎng)莖葡萄蕨藻的生長(zhǎng),說(shuō)明在天然海水中加鐵可有效促進(jìn)長(zhǎng)莖葡萄蕨藻的生長(zhǎng)。最適宜的Fe2+質(zhì)量濃度為3 mg/L,F(xiàn)e3+質(zhì)量濃度為6 mg/L。不同價(jià)態(tài)鐵的最適濃度不同,主要是因?yàn)樵孱悓?duì)兩種價(jià)態(tài)鐵離子的吸收機(jī)制不同,藻類可直接吸收利用外界環(huán)境中的Fe2+,僅小部分Fe3+可通過(guò)硫置換途徑進(jìn)入細(xì)胞,大部分需經(jīng)置換和蛋白轉(zhuǎn)運(yùn)兩個(gè)過(guò)程方可被吸收利用[23-25]。在光合色素的生成途徑中,需要以鐵卟啉環(huán)和δ-氨基-γ-酮戊二酸(ALA)為原料,鐵離子是二者生成原料之一,因此鐵可通過(guò)影響這兩種物質(zhì)的生成而影響光合色素的產(chǎn)生,從而導(dǎo)致色素含量發(fā)生變化[26]。研究表明,海帶()幼孢子體的葉綠素a和類胡蘿卜素含量隨著培養(yǎng)液中鐵濃度的增加而顯著增加[27],銅綠微囊藻()和慧氏微囊藻()葉綠素a和類胡蘿卜素的含量在富鐵培養(yǎng)液中明顯高于缺鐵的環(huán)境[28]。本研究中,添加兩種無(wú)機(jī)鐵主要對(duì)長(zhǎng)莖葡萄蕨藻葉綠素a和類胡蘿卜素含量有顯著影響,但對(duì)葉綠素b含量無(wú)顯著影響。
3.2 兩種無(wú)機(jī)鐵對(duì)長(zhǎng)莖葡萄蕨藻富鐵量的影響
一般而言,在植物耐受范圍內(nèi),植物對(duì)微量元素的富集量會(huì)隨環(huán)境中微量元素的升高而增加。目前已成功培育出多種富鐵陸生植物,在食用藻類富鐵強(qiáng)化方面亦培育出富鐵螺旋藻(1 426.40 μg/g,干基)、富鐵蛋白核小球藻(995.17 μg/g,干基)[14-15]。野生海藻鐵含量取決于沿岸海域中鐵離子含量,并呈季節(jié)性和地理性的變化[13]。關(guān)于天然長(zhǎng)莖葡萄蕨藻中鐵含量,唐賢明等[29]報(bào)道約為136.96 μg/g(干基),de Gaillande[9]報(bào)道為0.2 ~ 214 mg/kg(干基)。本研究中,測(cè)得對(duì)照組藻體鐵質(zhì)量分?jǐn)?shù)為103.62 μg/g,加入鐵后,藻體鐵含量隨培養(yǎng)液中Fe2+或Fe3+濃度的升高而增加,最高值分別可達(dá)1 522.54 μg/g和1373.93μg/g,增幅為1 369.34%和1 225.92%,表明長(zhǎng)莖葡萄蕨藻對(duì)兩種價(jià)態(tài)的鐵均有較強(qiáng)的富集能力,應(yīng)用無(wú)機(jī)鐵強(qiáng)化藻體鐵含量來(lái)生產(chǎn)富鐵功能海藻是可行的。在相同濃度下,F(xiàn)e2+處理組的富鐵量高于Fe3+處理組,是藻類對(duì)兩種價(jià)態(tài)鐵的吸收轉(zhuǎn)運(yùn)機(jī)制不同所致,F(xiàn)e2+具有更快的時(shí)效性[30]。
3.3 兩種無(wú)機(jī)鐵對(duì)長(zhǎng)莖葡萄蕨藻類抗氧化酶CAT、T-SOD活力的影響
T-SOD和CAT是兩種典型的抗氧化酶,它們通過(guò)將體內(nèi)形成的超氧化物轉(zhuǎn)換成毒性小或者無(wú)害物質(zhì)減少對(duì)植物造成的毒性損傷[31]。T-SOD在植物中主要以Mn-SOD、CuZn-SOD和Fe-SOD三種形式存在[32-33],大多數(shù)真核藻類的葉綠體基質(zhì)中,F(xiàn)e-SOD占主導(dǎo)地位。T-SOD通過(guò)控制金屬離子的得失電子對(duì)超氧化物陰離子自由基進(jìn)行歧化來(lái)維持藻類細(xì)胞的穩(wěn)定[34-35]。CAT是以鐵卟啉為輔基的結(jié)合酶,可將T-SOD酶產(chǎn)生的H2O2轉(zhuǎn)化為水。因此,在鐵元素充足狀況下,藻類體內(nèi)代謝旺盛,兩種酶活力較高,以分解代謝產(chǎn)生的毒害物質(zhì)[36-37]。本研究中,在Fe2+溶液培養(yǎng)條件下,兩種酶活力出現(xiàn)先升后降,再上升的現(xiàn)象,是因?yàn)樵弩w吸收鐵離子后會(huì)產(chǎn)生自由基,部分自由基一定程度上可抑制鐵對(duì)藻體的迫害,但當(dāng)鐵離子濃度持續(xù)增加時(shí),藻體產(chǎn)生的大量自由基又會(huì)對(duì)機(jī)體產(chǎn)生傷害,導(dǎo)致酶活力上升,以減少對(duì)藻體的迫[38];在Fe3+溶液培養(yǎng)條件下,兩種酶活力表現(xiàn)為先升高后平穩(wěn),這是因?yàn)樵弩w對(duì)不同價(jià)態(tài)鐵的利用機(jī)制不同[39]。從本研究可知,在實(shí)驗(yàn)鐵濃度范圍內(nèi)(0 ~ 12 mg/L),T-SOD和CAT保護(hù)作用基本與鐵濃度存在正相關(guān)性。當(dāng)添加的Fe2+質(zhì)量濃度為3 mg/L,藻體的兩種酶的活性最高;當(dāng)Fe3+濃度質(zhì)量達(dá)6 mg/L后,藻體T-SOD和CAT的活性隨Fe3+濃度升高相對(duì)穩(wěn)定,這時(shí)藻體的生長(zhǎng)也最佳。
3.4 兩種無(wú)機(jī)鐵對(duì)長(zhǎng)莖葡萄蕨藻總蛋白、VC和可溶性糖含量的影響
蛋白質(zhì)、VC和可溶性糖是衡量藻類營(yíng)養(yǎng)質(zhì)量的三個(gè)重要指標(biāo)。研究發(fā)現(xiàn),當(dāng)外界生長(zhǎng)環(huán)境改變時(shí),藻類的生長(zhǎng)及生理生化成分也會(huì)隨之改變。加入鐵后,藻類外界環(huán)境改變,藻體易受脅迫而導(dǎo)致蛋白質(zhì)的改變[40]。本研究中,兩種加鐵實(shí)驗(yàn)組的蛋白質(zhì)含量低于對(duì)照組,這與Xing等[41]報(bào)道的鐵使紫背浮萍()蛋白質(zhì)合成受到嚴(yán)重抑制的結(jié)果相似。這可能是因?yàn)樵弩w對(duì)鐵環(huán)境產(chǎn)生了脅迫反應(yīng),使藻體中葉綠素蛋白質(zhì)合成受抑制,從而導(dǎo)致總蛋白減少[40],或者是因?yàn)樵弩w鐵蛋白達(dá)到了一定的值后,鐵對(duì)其他種類蛋白質(zhì)合成和表達(dá)產(chǎn)生了負(fù)面影響從而降低了總蛋白含量[42]。實(shí)驗(yàn)組VC含量發(fā)生變化,是因?yàn)閂C是一種抗氧化劑,在鐵環(huán)境下細(xì)胞內(nèi)活性氧(ROS)含量改變,作為清除過(guò)量ROS的優(yōu)勢(shì)抗氧化劑,VC會(huì)隨ROS的變化而變化[43]。可溶性糖是植物體內(nèi)重要供能物質(zhì),也是一種信號(hào)傳遞物質(zhì),對(duì)植物生長(zhǎng)、增殖等過(guò)程有調(diào)控作用[44]。本研究中,加鐵組可溶性糖的含量均高于實(shí)驗(yàn)組,說(shuō)明鐵促進(jìn)植物生長(zhǎng),在生長(zhǎng)過(guò)程中植物進(jìn)行光合作用產(chǎn)生大量的碳水化合物,從而導(dǎo)致可溶性糖的含量升高[45-46]。
總的來(lái)說(shuō),在加鐵環(huán)境下,藻體除蛋白質(zhì)含量降低外,VC和可溶性糖含量均顯著增加,說(shuō)明添加鐵可較好地改善長(zhǎng)莖葡萄蕨藻的營(yíng)養(yǎng)品質(zhì),尤其是VC含量的顯著增加將有助于人體對(duì)鐵的吸收[12]。
4 結(jié)論
在培養(yǎng)海水中分別加入兩種無(wú)機(jī)鐵(Fe2+、Fe3+)后,藻體的特定生長(zhǎng)率、光合色素含量、鐵含量、兩種過(guò)氧化酶(CAT、T-SOD)活性以及總蛋白、維生素C、可溶性糖含量均受到影響。綜合考慮各指標(biāo),藻體在添加的Fe2+濃度為3 mg/L或Fe3+濃度為6 mg/L時(shí),長(zhǎng)莖葡萄蕨藻不僅獲得最佳的生長(zhǎng),而且可較好地改善其營(yíng)養(yǎng)品質(zhì)。因此,最適宜添加的Fe2+濃度為3 mg/L,最適宜的Fe3+濃度為6 mg/L。
[1] 黃建輝. 氮磷濃度及培養(yǎng)方式對(duì)長(zhǎng)莖葡萄蕨藻()生長(zhǎng)的影響[J]. 漁業(yè)研究, 2012, 34(5): 416-419.
[2] 姜芳燕, 宋文明, 楊寧, 等. 海南長(zhǎng)莖葡萄蕨藻的營(yíng)養(yǎng)成分分析及評(píng)價(jià)[J]. 食品工業(yè)科技, 2014, 35(24): 356-359.
[3] MATANJUN P, MOHAMED S, MUSTAPHA N M, et al. Nutrient content of tropical edible seaweeds,,and[J]. Journal of Applied Phycology, 2009, 21(1): 75-80.
[4] PAUL N A, NEVEUX N, MAGNUSSON M, et al. Comparative production and nutritional value of “sea grapes” — the tropical green seaweedsand[J]. Journal of Applied Phycology, 2014, 26(4): 1833-1844.
[5] MAEDA R, IDA T, IHARA H, et al. Immunostimulatory activity of polysaccharides isolated fromon macrophage cells[J]. Journal of the Agricultural Chemical Society of Japan, 2012, 76(3): 501-505.
[6] SHARMA B R, DONG Y R. Anti-diabetic effects of: stimulation of insulin secretion in pancreatic β-cells and enhancement of glucose uptake in adipocytes[J]. Asian Pacific Journal of Tropical Biomedicine, 2014, 4(7): 575-580.
[7] MAEDA R, IDA T, IHARA H, et al. Induction of Apoptosis in MCF-7 Cells by β-1,3-Xylooligosaccharides Prepared from[J]. Bioscience, Biotechnology and Biochemistry, 2012, 76(5): 1032-1034.
[8] 徐素萍. 微量元素鐵與人體健康的關(guān)系[J]. 中國(guó)食物與營(yíng)養(yǎng), 2007, 13(12): 53-56.
[9] DE GAILLANDE C, PAYRI C, REMOISSENET G, et al. Caulerpa consumption, nutritional value and farming in the Indo-Pacific region[J]. Journal of Applied Phycology, 2017, 29(5): 2249-2266.
[10] ALLEN L, de BENOIST B, DARY O, et al. Guidelines on food fortification with micronutrients[M]. Geneva: World Health Organization and Food and Agricultural Organization of the United Nations, 2006.
[11] GREGORY P J, WAHBI A, ADU-GYAMFI J, et al. Approaches to reduce zinc and iron deficits in food systems[J]. Global Food Security, 2017, 15: 1-10.
[12] GHARIBZAHEDI S M T, JAFARI S M. The importance of minerals in human nutrition: Bioavailability, food fortification, processing effects and nanoencapsulation[J]. Trends in Food Science & Technology, 2017, 62: 119-132.
[13] WELLS M L, POTIN P, CRAIGIE J S, et al. Algae as nutritional and functional food sources: revisiting our understanding[J]. Journal of Applied Phycology, 2017, 29(2): 949-982.
[14] 江涓. 螺旋藻對(duì)鐵的富集作用研究[J]. 湖北師范學(xué)院學(xué)報(bào)(自然科學(xué)版), 2002, 22(4): 25-27.
[15] 凌娜, 孫慶巖, 茅云翔, 等. 鐵對(duì)蛋白核小球藻生長(zhǎng)及營(yíng)養(yǎng)品質(zhì)的影響[J]. 中國(guó)海洋藥物, 2014, 33(4): 45-49.
[16] 郭輝. 海葡萄()切斷組織再生培養(yǎng)及發(fā)育條件研究[D]. 青島:中國(guó)科學(xué)院研究生院(海洋研究所), 2014.
[17] 蔡煜, 劉東超, 王曉梁, 等. 溫度、照度和氮磷營(yíng)養(yǎng)鹽對(duì)長(zhǎng)莖葡萄蕨藻富硒的影響[J]. 廣東海洋大學(xué)學(xué)報(bào), 2019, 39(3): 49-56.
[18] KOBAYASHI T, NOZOYE T, NISHIZAWA N K. Iron transport and its regulation in plants[J]. Free Radical Biology and Medicine, 2018, 133: 11-20.
[19] BRIAT J F O, DUBOS C, GAYMARD F. Iron nutrition, biomass production, and plant product quality[J]. Trends in Plant Science, 2015, 20(1): 33-40.
[20] CHEN B, ZOU D, YANG Y. Increased iron availability resulting from increased CO2enhances carbon and nitrogen metabolism in the economical marine red macroalga(Rhodophyta)[J]. Chemosphere, 2017, 173: 444-451.
[21] MARTIN J H, FITZWATER S E, GORDON R M. Iron deficiency limits phytoplankton growth in Antarctic waters[J]. Global Biogeochemical Cycles, 1990, 4(1): 5-12.
[22] MIKI O, NAGAI T, MARZUKI M, et al. Effects of Fe fertilizer eluate on the growth ofat the germling and immature stages[J]. Journal of Applied Phycology, 2016, 28(3): 1775-1782.
[23] SHAKED Y, KUSTKA A B, MOREL F M M. A general kinetic model for iron acquisition by eukaryotic phytoplankton[J]. Limnology and Oceanography, 2005, 50(3): 872-882.
[24] 凌娜, 仲星, 李紅秀, 等. 杜氏鹽藻對(duì)不同價(jià)態(tài)鐵的吸收機(jī)制研究[J]. 中國(guó)海洋藥物, 2015, 34(3): 29-34.
[25] 胡權(quán). 不同價(jià)態(tài)鐵對(duì)藻華暴發(fā)影響規(guī)律及其機(jī)理研究[D]. 廣州:廣東工業(yè)大學(xué), 2013.
[26] 姚波, 席北斗, 胡春明, 等. 鐵限制對(duì)浮游植物生長(zhǎng)和群落組成的影響研究綜述[J]. 生態(tài)環(huán)境學(xué)報(bào), 2010, 19(2): 459-465.
[27] SUZUKI Y, KUMA K, MATSUNAGA K. Effect of iron on oogonium formation, growth rate and pigment synthesis of(Phaeophyta)[J]. Fish Science, 1994, 60: 373-378.
[28] XING W, HUANG W M, LI D H, et al. Effects of iron on growth, pigment content, photosystem II efficiency, and siderophores production ofand[J]. Current Microbiology, 2007, 55(2): 94-98.
[29] 唐賢明, 劉小霞, 孟凡同, 等. 海馬齒和長(zhǎng)莖葡萄蕨藻的營(yíng)養(yǎng)成分分析及評(píng)價(jià)[J]. 熱帶生物學(xué)報(bào), 2018, 9(2): 129-135.
[30] BOTTGER L H, MILLER E P, ANDRESEN C, et al. Atypical iron storage in marine brown algae: a multidisciplinary study of iron transport and storage in[J]. Journal of Experimental Botany, 2012, 63(16): 5763-5772.
[31] 張容芳, 唐東山, 劉飛. 藻類抗氧化系統(tǒng)及其對(duì)逆境脅迫的響應(yīng)[J]. 環(huán)境科學(xué)與管理, 2011, 36(12): 21-25.
[32] MALLIK S, NAYAK M, SAHU B B. Response of antioxidant enzymes to high NaCl concentration in different salt-tolerant plants[J]. Biologia Plantarum, 2011, 55(1): 191-195.
[33] 黃苑媚, 劉志剛, 謝恩義, 等. 水流速率對(duì)全緣馬尾藻幼孢子體生長(zhǎng)和生理活性的影響[J]. 廣東海洋大學(xué)學(xué)報(bào), 2014, 34(6): 45-50
[34] HURTADO-GALLEGO J, MARTíN-BETANCOR K, RODEA-PALOMARES I, et al. Two novel cyanobacterial bioluminescent whole-cell bioreporters based on superoxide dismutases MnSOD and FeSOD to detect superoxide anion[J]. Chemosphere, 2018, 201: 772-779.
[35] 韓珊, 劉裕峰, 朱天輝, 等. 板栗鐵型超氧化物歧化酶基因(CmFeSOD) 的克隆及原核表達(dá)[J]. 西北農(nóng)業(yè)學(xué)報(bào), 2019, 28(6): 935-944.
[36] 劉建忠, 黃瑩瑩, 楊惠英. 金屬離子對(duì)過(guò)氧化氫酶的生物合成的影響[J]. 中山大學(xué)學(xué)報(bào)(自然科學(xué)版), 1999, 38(3): 123-124.
[37] 陳思嘉, 楊芳, 鄭文杰, 等. 硒脅迫對(duì)鈍頂螺旋藻抗氧化酶系統(tǒng)的影響[J]. 水生生物學(xué)報(bào), 2007, 31(5): 706-711.
[38] 鹿寧, 臧曉南, 張學(xué)成, 等. 逆境脅迫對(duì)藻類抗氧化酶系統(tǒng)的影響[J]. 武漢大學(xué)學(xué)報(bào)(理學(xué)版), 2012, 58(2): 119-124.
[39] 陳孝花. 除藻劑對(duì)黃絲藻藻華葉綠素a含量和抗氧化酶活性的影響[D]. 上海:上海海洋大學(xué), 2011.
[40] 魏思佳, 戴紹軍. 藻類響應(yīng)環(huán)境脅迫的蛋白質(zhì)組變化研究進(jìn)展[J]. 現(xiàn)代農(nóng)業(yè)科技, 2013, 20(3): 244-245.
[41] XING W, HUANG W M, LIU G H, et al. Effect of excess iron and copper on physiology of aquatic plant(L.) Schleid[J]. Environmental Toxicology, 2010, 25(2): 103-112.
[42] 章藝, 史鋒, 劉鵬, 等. 土壤中的鐵及植物鐵脅迫研究進(jìn)展[J]. 浙江農(nóng)業(yè)學(xué)報(bào), 2004, 16(2):110-114.
[43] KOCH K E, YING Z, WU Y, et al. Multiple paths of sugar-sensing and a sugar/oxygen overlap for genes of sucrose and ethanol metabolism[J]. Journal of Experimental Botany, 2000, 51: 417-427.
[44] 方云花. 大蒜可溶性糖分積累、分配與轉(zhuǎn)運(yùn)規(guī)律研究[D]. 西寧;青海大學(xué), 2013.
[45] 張琳, 崔紅米, 王建軍, 等. 鎘脅迫對(duì)不結(jié)球白菜Vc合成-半乳糖途徑基因表達(dá)及抗氧化系統(tǒng)的影響[J]. 植物生理學(xué)報(bào), 2015, 51(7): 1099-1108.
[46] 張進(jìn), 吳良?xì)g, 孔向軍, 等. 鐵鋅混合肥噴施對(duì)豌豆子粒鐵、鋅、可溶性糖和維生素C含量的影響[J]. 植物營(yíng)養(yǎng)與肥料學(xué)報(bào), 2006, 12(2): 245-249.
Effects of Two Types of Inorganic Iron on the Growth, Iron Content and Physiological Characteristics of
WANG Xiao-liang1,2, LIU Dong-chao1,2, XU Ri-sheng1,2, CHEN Hai-sheng1,2
(1.,524088,; 2.,524025,)
【】The effects of two types of inorganic ferrum on the growth, photosynthetic pigment, iron content, antioxidative enzymes activity and nutrient components of.【】Ferric chloride (FeCl3) and ferrous sulfate (FeSO4) were added as iron sources in diet and their effects on growth, photosynthetic pigment content, iron content, enzymatic activity (CAT, T-SOD) and total protein, Vitamin C and soluble sugar content ofwere studied in a single factor experiment.【】The specific growth rate, photosynthetic pigment concentration, iron content, antioxidative enzymes activity (CAT, T-SOD) and Vitamin C concentration of algae were significantly higher than the control group under Fe2+or Fe3+treatment (<0.05). The iron content of algae is significantly increased with the increase of Fe2+or Fe3+concentration in the culture solution with the highest values 1 522.54 μg/g and 1 373.93 μg/g respectively and the increase was by 1 369.34% and 1 225.92%respectively. The highest Vitamin C concentration of algae reached 15.41 μg/mg (for Fe2+) and 12.89 μg/mg (for Fe3+), increasing by 107.56% and 73.586%,respectively. Fe2+group had a significant effect on the soluble sugar content of algae (<0.05). The total protein content of algae was significantly lower than that of the control group (<0.05). 【】When the algae was under 3 mg/L Fe2+or 6 mg/L Fe3+treatment, the algae achieved the best growth and improvement in nutritional quality.
; growth; iron content; antioxidative enzymes; nutrient content
S968.4
A
1673-9159(2020)02-0020-08
10.3969/j.issn.1673-9159.2020.02.004
2019-12-02
廣東省科技計(jì)劃項(xiàng)目(2015A020209167)
王曉梁(1993-),男,碩士,主要從事藻類生理生態(tài)學(xué)研究。E-mail:544517970@qq.com
劉東超(1963-),男,副教授,主要從事藻類生理生態(tài)學(xué)研究。E-mail: dongchaoliu696@hotmail.com
王曉梁,劉東超,徐日升,等. Fe2+和Fe3+對(duì)長(zhǎng)莖葡萄蕨藻生長(zhǎng)、鐵含量和生理特性的影響[J]. 廣東海洋大學(xué)學(xué)報(bào),2020,40(2):20-27.
(責(zé)任編輯:劉慶穎)
