白酒雜醇油酯化酶的高效表達(dá)
2020-03-28 01:25:54宮春杰李安軍劉源才沈永祥汪江波薛棟升
中國(guó)釀造 2020年2期
宮春杰,李安軍,劉源才,張 嚴(yán),沈永祥,汪江波,薛棟升*
(1.湖北工業(yè)大學(xué) 生物工程與食品學(xué)院,湖北 武漢 430068;2.安徽古井貢酒股份有限公司,安徽 亳州 236800;3.勁牌有限公司,湖北 黃石 435000)
在白酒釀造行業(yè)中,雜醇油是影響酒質(zhì)及口感的主要因素之一[1]。通過原料控制及工藝優(yōu)化等手段不能從根本上解決這一問題。將能夠酯化雜醇油的酶類或菌株加入原料或酒曲參與發(fā)酵,不僅能夠降低雜醇油的含量,而且能夠增加酯類種類,賦予酒體更加飽滿的口感。小曲白酒和液態(tài)釀造的白酒最大的缺陷是較高的雜醇油含量造成飲用后容易頭痛和飲用時(shí)的苦澀味[2-4]。因此,酯化酶的篩選和表達(dá)對(duì)小曲白酒品質(zhì)的提升具有重要的意義。
酯化酶來源極為廣泛,根據(jù)其來源可大致分為動(dòng)物來源酯化酶、植物來源酯化酶及微生物來源酯化酶。其中動(dòng)植物來源的酯化酶提取工藝繁瑣,成本相對(duì)較高[5-7],因此用于實(shí)驗(yàn)或工業(yè)的酯化酶基本來源于微生物[8-10]。而且從酒曲、酒醅或其他極端環(huán)境中分離的微生物酯化酶,具有一些更加優(yōu)異的功能及催化活性[11-12]。微生物酯化酶具有催化條件溫和、能耗低、副產(chǎn)物少,具有高效、高選擇性、環(huán)境友好的優(yōu)點(diǎn),更適合于工業(yè)化大生產(chǎn),在基礎(chǔ)理論研究和實(shí)際應(yīng)用中都具有重大價(jià)值[12-16]。
傳統(tǒng)酒曲中一些產(chǎn)香酵母及細(xì)菌也能產(chǎn)生催化作用的酯化酶,但只能催化乙酸乙酯、己酸乙酯等的合成,對(duì)雜醇油無明顯的催化作用[17-20]。雜醇油酯化酶和構(gòu)建高效表達(dá)雜醇油的酵母菌株鮮有報(bào)道。
本實(shí)驗(yàn)以從深海沉積物中篩選出產(chǎn)雜醇油酯化酶菌株的總核糖核酸(ribonucleic acid,RNA)為模板,采用逆轉(zhuǎn)錄聚合酶鏈?zhǔn)椒磻?yīng)(reverse transcription-polymerase chain reaction,RT-PCR)方法擴(kuò)增近平滑假絲酵母雜醇油酯化酶基因,并于畢赤酵母中高效表達(dá)。酯化雜醇油的酯化酶不僅對(duì)提升小曲白酒和液態(tài)釀造的白酒的品質(zhì)具有重要的意義,而且對(duì)整個(gè)釀酒行業(yè)提升白酒的品質(zhì)具有重要的實(shí)際應(yīng)用意義。
1 材料與方法
1.1 材料與試劑
菌株:從深海沉積物中篩選出的產(chǎn)雜醇油酯化酶的近平滑假絲酵母(Candida parapsilosis);畢赤酵母(Pichia pastoris)GS115:真核表達(dá)宿主;質(zhì)粒pPICZαA:外分泌型表達(dá)載體,亞克隆于大腸桿菌(Esherichia coli)DH5α。
試劑:乙酸正戊酯、乙酸正丙酯、乙酸異丁酯、乙酸異戊酯及乙醇等(均為色譜級(jí)):上海阿拉丁生化科技股份有限公司;其他試劑盒:天根生化科技(北京)有限公司;限制性內(nèi)切酶:寶生物工程(大連)有限公司。
種子培養(yǎng)基:蛋白胨10 g,牛肉膏3 g,葡萄糖5 g,陳海水補(bǔ)至1 000 mL,pH 7.6,115 ℃滅菌30 min。
產(chǎn)酶培養(yǎng)基:胰蛋白胨10 g,麩皮25 g,K2HPO41.0 g,MgSO4·7H2O 0.5 g,NaCl 10.0 g,自來水補(bǔ)至1 000 mL,pH 7.5,121 ℃滅菌20 min。
1.2 儀器與設(shè)備
SC005675聚合酶鏈?zhǔn)椒磻?yīng)(polymerase chain reaction,PCR)分析儀、Tanon-1600Bio-Rad凝膠成像系統(tǒng)儀:美國(guó)Bio-Rad公司;TG16-W高速冷凍離心機(jī):美國(guó)Thermo Fisher Scientific公司;JY92-II氣相色譜儀:寧波新芝生物科技有限公司;HP-101毛細(xì)管氣相色譜柱(50 m×0.53 mm×0.50 μm):德國(guó)Merck公司。
1.3 方法
1.3.1 雜醇油酯化酶的提取及含量測(cè)定
產(chǎn)酯化酶的近平滑假絲酵母或構(gòu)建的酵母菌先于種子培養(yǎng)基上28 ℃、200 r/min活化18 h,以2%接種量接種于產(chǎn)酶培養(yǎng)基,產(chǎn)酶培養(yǎng)基裝液量70 mL/250 mL,30 ℃、200 r/min培養(yǎng)3d。發(fā)酵醪液3層干凈紗布過濾,濾液于臺(tái)式高速冷凍離心機(jī)5000r/min離心5min,除去細(xì)小麩皮殘?jiān)熬昃w。上清液以0.22μm尼龍濾膜過濾,除去菌體,得到酯化酶粗酶液。
酯化體系:雜醇油含量(正丙醇、異丁醇、異戊醇)各1%;乙酸含量2%;粗酶液含量50%;水含量45%。先加水、雜醇油及乙酸,再加入酶液。且在加入粗酶液前,以200 g/L NaOH溶液將體系pH調(diào)至3.5。粗酶液均做2組酯化體系,其中1組為實(shí)驗(yàn)組,另1組為對(duì)照組。對(duì)照組為酶液以等量的去離子水代替。酯化體系30 ℃、200 r/min酯化24 h。
以頂空氣相色譜法測(cè)量酯化產(chǎn)物中雜醇油酯的含量[21-22]。氣相色譜條件為色譜柱:HP-101毛細(xì)管氣相色譜柱(50 m×0.53 mm×0.50 μm);柱溫:二階段升溫:初始溫度35 ℃,保持7 min,以4 ℃/min升至60 ℃,再以6 ℃/min升至105℃,最后以20℃/min升至200℃,保持10min;進(jìn)樣口溫度200℃,檢測(cè)器溫度220℃,載氣為高純氮?dú)猓瑲錃猓?0.0mL/min;空氣流量:200 mL/min;柱前壓為30 kPa,柱流量為1.12~1.36 mL/min;尾吹30.0 mL/min;分流比40∶1;進(jìn)樣量1 mL;內(nèi)標(biāo)為2%乙酸正戊酯。
1.3.2 酯化酶基因PCR擴(kuò)增
根據(jù)酯化酶序列,利用引物設(shè)計(jì)軟件Primer 5.0設(shè)計(jì)特異引物,引物序列如下:
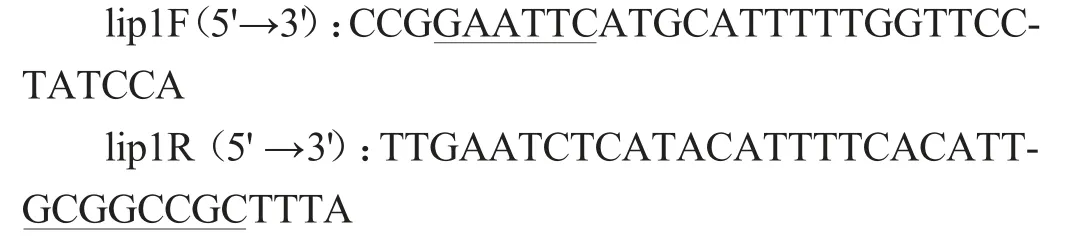
上游引物加入保護(hù)堿基CCG 和EcoRI 酶切位點(diǎn)GAATTC,下游引物加入保護(hù)堿基TTTA和NotI酶切位點(diǎn)GCGGCCGC。
1.3.3 總RNA提取
將活化的近平滑假絲酵母(Candida parapsilosis)接種至種子培養(yǎng)基中,30 ℃培養(yǎng)24 h,按賽默飛公司RNA Mini Kit說明書步驟提取總核糖核酸(RNA)。提取的酵母菌總RNA用1%瓊脂糖電泳,電泳電壓為150 V,電泳25 min。
1.3.4 逆轉(zhuǎn)錄聚合酶鏈反應(yīng)擴(kuò)增雜醇油酯化酶編碼序列
通過以提取的酵母菌Lip-8總RNA為模板,反轉(zhuǎn)錄合成cDNA第一鏈,并進(jìn)行目的基因的擴(kuò)增。本步驟采用天根生化科技(北京)有限公司的Quant One Step(reverse transcription-polymerase chain reaction,RT-PCR)kit(KR113)進(jìn)行操作,其反應(yīng)體系及PCR條件均按試劑盒說明書進(jìn)行。PCR擴(kuò)增產(chǎn)物采用0.8%瓊脂糖電泳,80 V、35 min,用凝膠成像儀檢測(cè)目的條帶的大小,PCR產(chǎn)物回收純化測(cè)序。
1.3.5 表達(dá)載體構(gòu)建
將反轉(zhuǎn)錄獲取的PCR產(chǎn)物及提取的空載質(zhì)粒pPICZαA分別進(jìn)行雙酶切,再以T4連接酶16 ℃過夜連接。化學(xué)轉(zhuǎn)化法導(dǎo)入空載Esherichia coliDH5α[23],得到亞克隆菌株。再提取質(zhì)粒,以BamH I酶酶切線性化,按照BTX電穿孔儀ECM 830自帶畢赤酵母電轉(zhuǎn)程序,進(jìn)行電轉(zhuǎn)化。
1.3.6 重組子的誘導(dǎo)表達(dá)及蛋白電泳
獲得的酵母轉(zhuǎn)化子活化后,每24 h補(bǔ)加甲醇至終濃度1%,誘導(dǎo)畢赤酵母表達(dá)酯化酶[24-25]。誘導(dǎo)結(jié)束后,離心取上清液進(jìn)行十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳(sodium dodecyl sulphate-polyacrylamide gel electrophoresis,SDSPAGE)分析。樣品與5倍上樣緩沖液混合均勻后,沸水浴5 min,再10 000 r/min常溫離心2 min,儲(chǔ)存于4 ℃?zhèn)溆谩悠飞蠘恿繛?5 μL,濃縮膠電泳電壓為80 V,分離膠電泳電壓為130 V。電泳結(jié)束后以快速染色脫色法進(jìn)行處理[26]。
2 結(jié)果與分析
2.1 近平滑假絲酵母酯化酶酯化雜醇油的性能
菌株所產(chǎn)的酯化酶粗酶液加入酯化體系酯化,測(cè)量酯化液中乙酸正丙酯、乙酸異丁酯、乙酸異戊酯的質(zhì)量濃度。將所測(cè)得的實(shí)驗(yàn)組質(zhì)量濃度減去空白組質(zhì)量濃度后,得到實(shí)際酯化合成的質(zhì)量濃度結(jié)果見表1。由表1可知,近平滑假絲酵母產(chǎn)生的酯化酶具有一定的酯化正丙醇、異丁醇和異戊醇的能力。

表1 近平滑假絲酵母酯化酶酯化雜醇油的性質(zhì)Table 1 Properties of esterase from Candida parapsilosis for fusel oil esterification
2.2 酯化酶基因擴(kuò)增
2.2.1 近平滑假絲酵母總RNA提取
采用總RNA提取試劑盒法從近平滑假絲酵母中提取總RNA,提取的總RNA電泳分析見圖1。由圖1可知,提取的RNA條帶較清晰,完整性較好,純度也較高,可以用于反轉(zhuǎn)錄合成互補(bǔ)脫氧核糖核酸(complementary deoxyribonucleic acid,cDNA)。
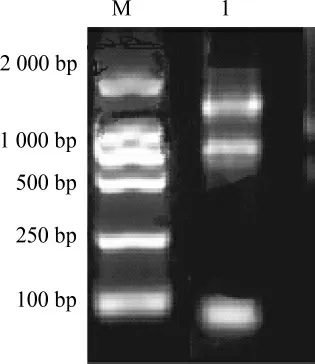
圖1 近平滑假絲酵母總RNA瓊脂糖凝膠電泳結(jié)果Fig.1 Agarose gel electrophoresis results of total RNA of Candida parapsilosis
2.2.2 近平滑假絲酵母酯化酶基因RT-PCR
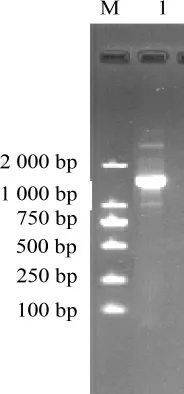
圖2 近平滑假絲酵母酯化酶反轉(zhuǎn)錄PCR產(chǎn)物瓊脂糖凝膠電泳結(jié)果Fig.2 Agarose gel electrophoresis results of reverse transcriptase-PCR product of esterase from Candida parapsilosis
以總RNA為模板,以QuantOneStepRT-PCRkit進(jìn)行PCR,近平滑假絲酵母酯化酶反轉(zhuǎn)錄PCR產(chǎn)物電泳圖見圖2。由圖2可知,PCR產(chǎn)物大小接近于1 500 bp,與預(yù)期片段大小一致。純化后測(cè)序,推出酯化酶的氨基酸序列,結(jié)果見圖3。

圖3 酯化酶氨基酸序列Fig.3 Amino acid sequence of esterase
2.3 畢赤酵母中表達(dá)雜醇油酯化酶基因
2.3.1 重組子的誘導(dǎo)表達(dá)及SDS-PAGE
誘導(dǎo)結(jié)束后取上清進(jìn)行SDS-PAGE分析,結(jié)果見圖4。孔道1為誘導(dǎo),孔道2為未誘導(dǎo),根據(jù)蛋白質(zhì)電泳圖分析可知,孔道1于44.3~66.4 kDa間多出一個(gè)條帶。根據(jù)蛋白質(zhì)數(shù)據(jù)庫(kù)(protein data bank,PDB)及美國(guó)國(guó)家生物技術(shù)信息中心(national center for biotechnology information,NCBI)數(shù)據(jù)庫(kù)中l(wèi)ipase 1編碼氨基酸計(jì)算,lipase 1分子質(zhì)量為50.86 kDa,與目的條帶接近。結(jié)果表明構(gòu)建的載體能轉(zhuǎn)化入畢赤酵母細(xì)胞,并能啟動(dòng)雜醇油酯化酶基因的表達(dá)。
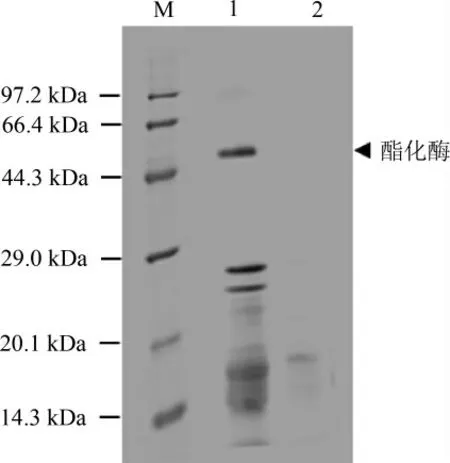
圖4 畢赤酵母表達(dá)的酯化酶的SDS-PAGE結(jié)果Fig.4 SDS-PAGE of esterase expressed by Pichia pastoris
2.3.2 構(gòu)建的畢赤酵母表達(dá)雜醇油酯化酶
重組子誘導(dǎo)表達(dá)結(jié)束后,以5 000×g、4 ℃離心5 min,上清即為酯化酶粗酶液。按1.3方法進(jìn)行酶活的檢測(cè),結(jié)果見表2。由表2可知,根據(jù)頂空氣相色譜的檢測(cè)結(jié)果,該酯化酶活性比原始菌株表達(dá)酶活高。畢赤酵母表達(dá)的酯化酶酯化合成乙酸正丙酯、乙酸異丁酯、乙酸異戊酯的合成量比近平滑假絲酵母的酯化酶合成的量分別增加1.63倍、1.15倍、0.99倍,結(jié)果表明,酯化酶基因在畢赤酵母中高效表達(dá)。

表2 畢赤酵母表達(dá)的酯化酶酶活Table 2 Esterase activity expressed by Pichia pastoris
2.3.3 雜醇油酯化酶分子模型分析
根據(jù)已知的lipase的蛋白質(zhì)序列,通過Discovery Studio軟件及“Swiss-model Workspace”[27]進(jìn)行l(wèi)ipase的模型構(gòu)建。根據(jù)已報(bào)道的Candida antarctica酯化酶Lipase A(PDB:3guu.1)[28]空間結(jié)構(gòu)為模板,構(gòu)建的空間結(jié)構(gòu)圖見圖5。構(gòu)建的空間結(jié)構(gòu)模型以PyMoL軟件分析[29]。由圖5可知,lipase與傳統(tǒng)的酯化酶空間構(gòu)型相差無幾,均為8個(gè)α螺旋圍成的“8套環(huán)”的環(huán)域結(jié)構(gòu),底物固定于環(huán)中進(jìn)行催化合成。
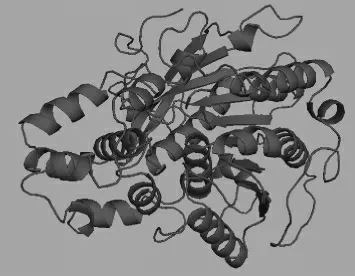
圖5 酯化酶的分子模型Fig.5 Molecular model of esterase
3 結(jié)論
本研究解析出雜醇油酯化酶的氨基酸序列,并對(duì)雜醇油酯化酶基因進(jìn)行了異源表達(dá),構(gòu)建了高效表達(dá)雜醇油酯化酶的酵母工程菌株。構(gòu)建的酵母菌株能酯化正丙醇、異丁醇和異戊醇。雜醇油酯化酶不僅能豐富了酒體酯類含量,還能大幅度降低蒸餾的酒體中的雜醇油含量從而極大的提升白酒品質(zhì)。本研究構(gòu)建的高效表達(dá)雜醇油酯化酶的酵母菌株對(duì)提升釀造白酒的品質(zhì)具有重要的實(shí)際意義。
