抗菌肽brevilaterin與ε-聚賴氨酸對金黃色葡萄球菌的協同抑菌機理
2020-03-31 04:29:28寧亞維付浴男韓盼盼王志新賈英民
食品科學 2020年5期
寧亞維,蘇 丹,付浴男,韓盼盼,王志新,賈英民,
(1.河北科技大學生物科學與工程學院,河北 石家莊 050018;2.北京工商大學食品與健康學院,北京 100048)
金黃色葡萄球菌(Staphylococcus aureus)廣泛分布于環境中,極易造成食品污染,是食品中常見的食源性致病菌[1]。金黃色葡萄球菌能引起嚴重的感染,甚至威脅人類和動物的生命,如皮膚和軟組織感染、中毒性休克綜合征和敗血癥[2]。因此,抑制食品中金黃色葡萄球菌的生長繁殖對提高食品的安全性至關重要。
抗菌肽brevilaterin是由側孢短芽孢桿菌產生的具有廣譜抑菌活性的小分子肽,具有熱穩定性好、生產成本低等優點,作為食品防腐劑具有較好的開發潛力[3]。課題組前期研究發現抗菌肽brevilaterin對細菌、真菌均具有抑菌能力,在與天然防腐劑聯合抑菌效應的研究中發現,抗菌肽brevilaterin與ε-聚賴氨酸對金黃色葡萄球菌的最小抑菌濃度(minimal inhibit concentration,MIC)分別為10 AU/mL、0.015 6 mg/mL,分級抑菌濃度指數為0.5,兩者聯用具有協同抑菌效應[4]。因此,本研究擬考察抗菌肽brevilaterin與ε-聚賴氨酸聯用對金黃色葡萄球菌的抑菌機理,為降低其在食品防腐中的添加量從而降低使用成本提供理論參考。ε-聚賴氨酸是由白色鏈霉菌產生的陽離子多肽,具抑菌譜廣等優點[5-6]。ε-聚賴氨酸作用于菌體細胞后,可通過改變細胞膜通透性[7]、破壞細胞結構[8]、抑制蛋白質合成[9],以及與菌體DNA結合[10],從而發揮抑菌作用。Ye Ruosong等[11]對ε-聚賴氨酸的抑菌機理研究表明,ε-聚賴氨酸可以通過活性氧氧化應激以及基因調控等方面發揮抑菌作用。而抗菌肽brevilaterin及其與ε-聚賴氨酸聯用的抑菌機理尚不明確,因此本研究將對抗菌肽brevilaterin與ε-聚賴氨酸的聯合抑菌機理進行研究。
據報道大多數抗菌肽首先通過破壞細菌細胞膜進入胞內發揮抑菌作用,如抗菌肽LL-37[12]、抗菌肽polyphemusin[13],但是有些抗菌肽可以不破壞細菌細胞膜完整性而進入胞內,通過抑制DNA和RNA合成來達到殺菌效果,例如抗菌肽microcin B17[14]、抗菌肽buforin II[15]均具有抑制DNA復制的功能。大多數抗菌肽可以通過多靶位發揮抑菌作用,如抗菌肽P-1可引起粉紅單端孢細胞膜通透性增加,抑制蛋白質及核酸合成,導致菌體代謝紊亂,進而達到抑菌作用[16]。因此,本研究將從細胞膜質子動力勢、細胞膜完整性、細胞膜代謝活力、細胞超微結構、蛋白質以及核酸合成等方面研究抗菌肽brevilaterin與ε-聚賴氨酸對金黃色葡萄球菌的聯合抑菌機理,以期為抗菌肽brevilaterin與ε-聚賴氨酸在食品中的應用提供理論依據。
1 材料與方法
1.1 材料與試劑
金黃色葡萄球菌(S. aureus ATCC 25923)河北科技大學食品生物技術與安全實驗室保藏;抗菌肽brevilaterin 河北科技大學食品生物技術與安全實驗室制備[17]。
ε-聚賴氨酸 上海麥克林生化科技有限公司;DiSC3(5)、BCECF-AM 美國Sigma公司;紅四氮唑上海源葉生物科技有限公司;正丁醇 天津市永大化學試劑有限公司;LIVE/DEAD BacLight細菌活力試劑盒 美國Thermo Fisher Scientific公司;低分子質量蛋白Marker、DNA Ladder 北京索萊寶科技有限公司;細菌基因組DNA提取試劑盒 天根生化科技有限公司;Agarose G-10 法國Biowest公司。
1.2 儀器與設備
Evolution 220型紫外分光光度計 美國Thermo Fisher Scientific公司;F-7000-FL型熒光分光光度計、H-7650型透射電子顯微鏡(transmission electron microscope,TEM) 日本日立公司;Accuri C6 plus型流式細胞儀 美國Becton Dickinson公司;BX53型熒光顯微鏡 日本奧林巴斯株式會社; Gel Doc XR+型凝膠成像系統 美國Bio-Rad公司。
1.3 方法
1.3.1 實驗分組
所有實驗均按抑菌劑種類和終濃度將實驗分為6 組,即2 個抗菌肽組(MIC和1/4 MIC)、2 個ε-聚賴氨酸組(MIC和1/4 MIC)、聯用組(1/4 MIC抗菌肽brevilaterin+1/4 MIC ε-聚賴氨酸)、對照組(未添加抑菌劑)。
1.3.2 抑菌動力學的測定
將對數期金黃色葡萄球菌菌懸液(106CFU/mL)與等體積不同抑菌劑水溶液混合,使抗菌肽brevilaterin組終濃度分別為MIC、1/4 MIC,ε-聚賴氨酸組的ε-聚賴氨酸組終濃度分別為MIC、1/4 MIC,聯用組抗菌肽brevilaterin、ε-聚賴氨酸的終濃度均為1/4 MIC,以質量分數0.85%生理鹽水作為對照組。在37 ℃培養箱孵育,定時取出。取稀釋好的菌懸液100 μL進行平板涂布,37 ℃孵育24 h后進行菌落計數。
1.3.3 質子動力勢的測定
將對數期金黃色葡萄球菌用5 mmol/L Hepes緩沖液(含10 mmol/L葡萄糖、100 mmol/L氯化鉀,pH 7.2)清洗、重懸,加入DiSC3(5)熒光探針(終濃度為1 μmol/L),并于30 ℃避光孵育15 min。在比色皿中加入菌懸液(終濃度為108CFU/mL)與等體積的抑菌劑,以纈氨霉素(Valinomycin,Val)作為陽性對照、尼日利亞菌素(Nigericin,Nig)為陰性對照,終濃度均為1 μmol/L。使用熒光分光光度計進行時間掃描,其中激發波長為650 nm,發射波長為672 nm。
將對數期金黃色葡萄球菌用10 mmol/L葡萄糖溶液(pH 6)清洗、重懸為3×108CFU/mL,加入BCECF-AM熒光探針(終濃度1 μmol/L),于30 ℃避光孵育20 min。菌懸液與抑菌劑按體積比1∶1混合,以Nig作為陽性對照,Val作為陰性對照(終濃度均為5 μmol/L),使用熒光分光光度計在激發波長502 nm、發射波長525 nm處進行時間掃描。
1.3.4 呼吸鏈脫氫酶活力的測定
參考馮建嶺[18]的方法,通過測定抑菌劑作用后細胞膜呼吸鏈脫氫酶的變化研究金黃色葡萄球菌細胞代謝活力。向具塞試管中分別加入1 mL對數期菌液,Tris-HCl緩沖液(pH 8.6、0.05 mol/L)、葡萄糖溶液(0.1 mol/L)以及紅四氮唑(1 mg/mL)各2 mL,充分搖晃均勻。然后加入1 mL抑菌劑,以無菌水作為對照組,37 ℃孵育5 h后滴加兩滴濃硫酸終止反應。最后向試管中加入5 mL正丁醇振蕩萃取,然后靜置8 min,測量上層液體于490 nm波長處的吸光度。
1.3.5 細胞膜完整性的測定
將對數期金黃色葡萄球菌用磷酸鹽緩沖液(phosphate buffered saline,PBS)(0.02 mol/L、pH 7.2)清洗、重懸。菌懸液(終濃度為108CFU/mL)與不同濃度抑菌劑等體積混合。在37 ℃條件下培養1 h后用PBS清洗、重懸。加入熒光染料SYTO 9和碘化丙啶(propidiumiodide,PI),使其終濃度均為2 μmol/L,30 ℃條件下避光孵育15 min,用PBS清洗、重懸后,分別采用流式細胞儀和熒光顯微鏡對染色細胞進行計數分析與觀察拍照。
1.3.6 菌體超微結構的觀察
將對數期金黃色葡萄球菌用PBS(0.01 mol/L、pH 7.2)清洗、重懸后,分別向菌懸液(終濃度為2×108CFU/mL)中加入各抑菌劑。37 ℃下培養1 h后,采用PBS清洗兩遍,然后用體積分數4%戊二醛4 ℃固定過夜。之后,再用體積分數1%鋨酸在4 ℃固定3 h,用不同體積分數(30%、50%、70%、80%、90%)的丙酮溶液逐級脫水后,再用無水丙酮置換兩次。用包埋劑包埋、聚合后進行超薄切片,采用醋酸雙氧鈾進行染色后,使用TEM觀察拍照。
1.3.7 菌體蛋白質合成的測定
采用十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳測定蛋白,具體參考Ning Houqi等[19]的方法,并進行適當修改。將培養至對數生長期的金黃色葡萄球菌用生理鹽水清洗、重懸后與抑菌劑混合(菌懸液終濃度為3×108CFU/mL)。在37 ℃條件下孵育4 h后,用生理鹽水清洗、重懸至75 μL加入4×蛋白上樣緩沖液,在100 ℃沸水中加熱5 min后,取出放至室溫25 ℃。安裝好電泳裝置后,于上樣孔中加入10 μL樣品,調電壓至80 V開始電泳,待溴酚藍進入分離膠后120 V跑至膠底。將膠塊放置于染色液(包括1 g/L考馬斯亮藍G250和45%(體積分數,下同)甲醇、10%冰醋酸、45%蒸餾水)中搖床染色20 min后,放入脫色液(包括10%甲醇、10%冰醋酸、80%蒸餾水)中進行搖床脫色,直至條帶清晰后使用凝膠成像儀拍照記錄結果。
1.3.8 菌體DNA的測定
采用DNA試劑盒提取金黃色葡萄球菌基因組DNA,使用超微量紫外分光光度計測定所提DNA的質量濃度(48 μg/mL)和純度(OD260nm/OD280nm=1.89)。制備質量分數0.8%的瓊脂糖凝膠,其中加入4S Red Plus 核酸染色劑(終濃度為0.1 μL/mL)。將DNA(終質量濃度為12 μg/mL)與各濃度抑菌劑混合,在37 ℃下水浴30 min后,進行瓊脂糖凝膠阻滯電泳并使用凝膠成像儀拍照觀察。
1.4 數據處理與統計分析
所有實驗均重復3 次取其平均值。采用Origin 8.0軟件通過單因素方差分析法對實驗結果進行統計分析并作圖,P<0.05為差異顯著。
2 結果與分析
2.1 抗菌肽brevilaterin與ε-聚賴氨酸聯用對金黃色葡萄球菌的抑菌動力學結果
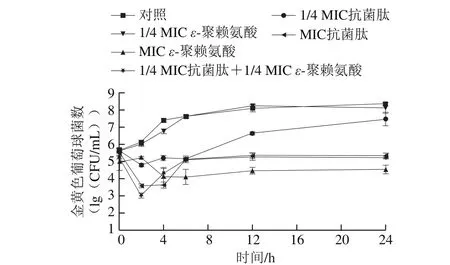
圖 1 抗菌肽brevilaterin與ε-聚賴氨酸聯用對金黃色葡萄球菌數的影響Fig. 1 Time-kill curves of brevilaterin and/or ε-polylysine against S. aureus
基于前期抗菌肽brevilaterin和ε-聚賴氨酸協同抑菌效應的評價結果[4],選取1/4 MIC抗菌肽brevilaterin和1/4 MIC ε-聚賴氨酸進行復配并考察兩者的協同抑菌動力學。由圖1可以看出,MIC抗菌肽brevilaterin、MIC ε-聚賴氨酸以及聯用組(1/4 MIC抗菌肽brevilaterin和1/4 MIC ε-聚賴氨酸)均能夠抑制金黃色葡萄球菌的生長,且聯用組的活菌數明顯低于1/4 MIC抗菌肽brevilaterin和1/4 MIC ε-聚賴氨酸單獨使用組的活菌數,結果進一步證實了抗菌肽brevilaterin與ε-聚賴氨酸的協同抑菌效果。
2.2 抗菌肽brevilaterin與ε-聚賴氨酸對金黃色葡萄球菌細胞膜質子動力勢的影響
質子動力勢由跨膜電勢和跨膜pH值梯度構成,參與ATP合成、活性離子轉運、蛋白磷酸化等多種生物學過程[20-21]。DiSC3(5)是一種膜電位敏感探針,在正常細胞中處于自猝滅狀態;當膜電位被改變時,DiSC3(5)會釋放到介質中,熒光強度增加[22],因此采用熒光探針DiSC3(5)考察了抗菌肽brevilaterin與ε-聚賴氨酸對細胞跨膜電位的影響。結果表明,MIC抗菌肽brevilaterin處理后DiSC3(5)熒光強度迅速增加(圖2A),說明其對細胞跨膜電位有較大影響,且作用效果與陽性對照Val類似。MIC ε-聚賴氨酸、1/4 MIC ε-聚賴氨酸、1/4 MIC抗菌肽brevilaterin對膜電位沒有明顯影響,聯用組的熒光強度稍高于單個組分處理組。結果表明,抗菌肽brevilaterin會破壞金黃色葡萄球菌跨膜電勢,ε-聚賴氨酸對金黃色葡萄球菌跨膜電勢無明顯影響。
BCECF-AM進入細胞后被細胞內的酯酶水解為BCECF留在細胞內,在適當pH值下可被激發產生熒光,且細胞內pH值越高,BCECF探針的熒光強度越強[23]。因此,采用熒光探針BCECF-AM研究了抗菌肽brevilaterin與ε-聚賴氨酸對金黃色葡萄球菌跨膜pH值梯度的影響。Nig可破壞細胞膜的跨膜pH值梯度,以其作為陽性對照,Val對細胞膜跨膜pH值梯度無影響,以其作為陰性對照[24]。如圖2B所示,1/4 MIC ε-聚賴氨酸、MIC ε-聚賴氨酸以及聯用組的熒光強度變化較大,1/4 MIC抗菌肽brevilaterin、MIC抗菌肽brevilaterin組的BCECF熒光強度沒有明顯變化。結果表明,ε-聚賴氨酸會破壞金黃色葡萄球菌的跨膜pH值梯度,而抗菌肽brevilaterin對金黃色葡萄球菌的跨膜pH值梯度無明顯影響。
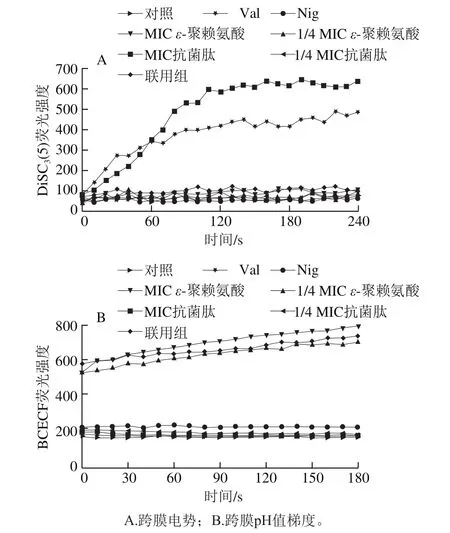
圖 2 抗菌肽brevilaterin與ε-聚賴氨酸對金黃色葡萄球菌質子動力勢的影響Fig. 2 Effect of brevilaterin combined with ε-polylysine on transmembrane proton potential of S. aureus
2.3 抗菌肽brevilaterin與ε-聚賴氨酸聯用對細胞膜完整性的影響
利用PI/SYTO 9熒光探針檢測抗菌肽brevilaterin與ε-聚賴氨酸對細胞膜完整性的影響[25]。SYTO 9探針通常會標記群體中的所有細菌,而PI只能標記細胞膜受損的細菌。當兩種探針同時存在時,PI會與SYTO 9發生競爭性結合,導致SYTO 9染色熒光減少。流式細胞儀(圖3、4)結合熒光顯微鏡圖(圖5)結果顯示,對照組(膜破損率為9.4%)與1/4 MIC ε-聚賴氨酸(膜破損率為10.4%)幾乎全部發出綠色熒光,而經過MIC抗菌肽brevilaterin處理后的細胞因為膜破損較嚴重(膜破損率高達97.3%),PI與SYTO 9發生競爭性結合顏色呈橙紅色。1/4 MIC抗菌肽brevilaterin、MIC ε-聚賴氨酸以及聯用組處理后,檢測到部分紅色熒光,說明部分細胞膜受損(破損率分別為36.3%、22.6%、51.3%),MIC抗菌肽brevilaterin組檢測到大量紅色熒光,幾乎沒有綠色熒光,說明細胞膜的破損較嚴重。結果表明,MIC抗菌肽brevilaterin可以完全破壞細胞膜完整性,而MIC ε-聚賴氨酸僅造成部分膜完整性損傷,聯用組對膜完整性的破壞程度約等于抗菌肽brevilaterin與ε-聚賴氨酸二者的加和,說明1/4 MIC抗菌肽brevilaterin與1/4 MIC ε-聚賴氨酸對金黃色葡萄球菌膜完整性的破壞作用具有疊加性。
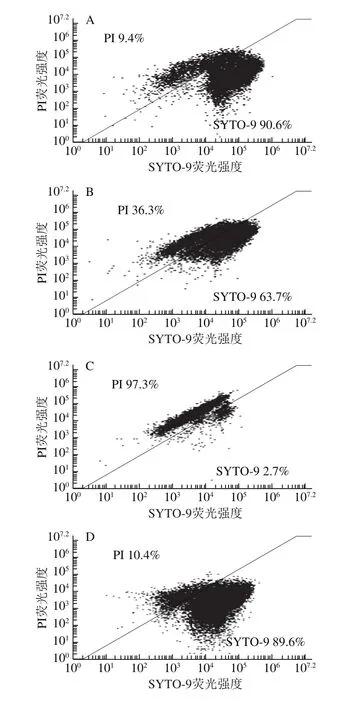
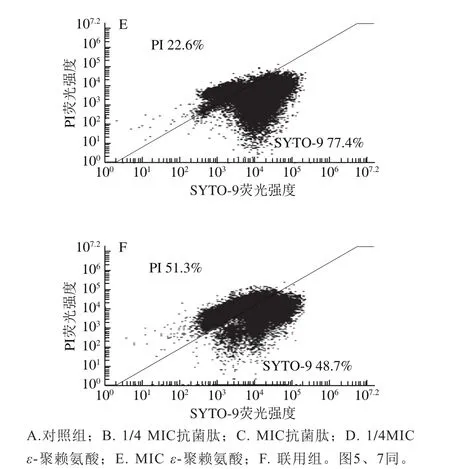
圖 3 抗菌肽brevilaterin與ε-聚賴氨酸作用于金黃色葡萄球菌的流式細胞圖Fig. 3 Effect of brevilaterin combined with ε-polylysine on flow cytometry analysis of S. aureus
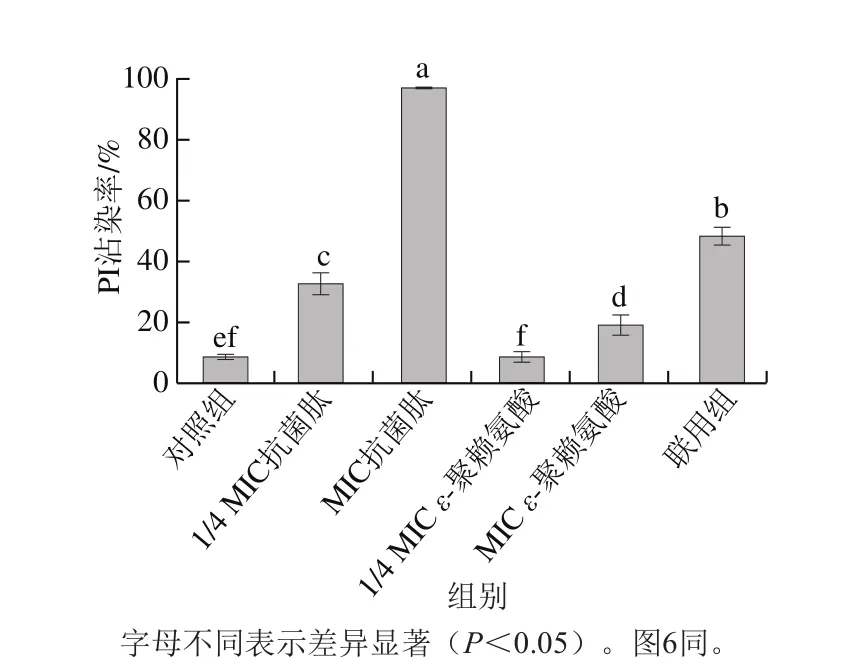
圖 4 抗菌肽brevilaterin與ε-聚賴氨酸對金黃色葡萄球菌細胞膜完整性的影響Fig. 4 Effect of brevilaterin combined with ε-polylysine on membrane integrity of S. aureus
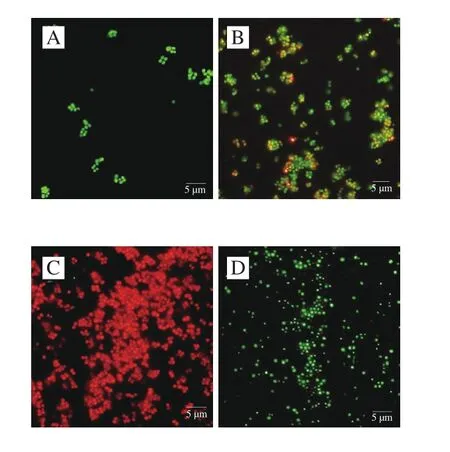
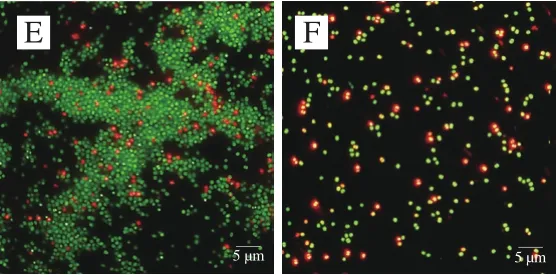
圖 5 抗菌肽brevilaterin與ε-聚賴氨酸作用于金黃色葡萄球菌的熒光顯微鏡圖Fig. 5 Effect of brevilaterin combined with ε-polylysine on membrane integrity of S. aureus evaluated by fluorescence microscopy
2.4 抗菌肽brevilaterin與ε-聚賴氨酸對細胞膜代謝活力的影響

圖 6 抗菌肽brevilaterin與ε-聚賴氨酸對金黃色葡萄球菌呼吸鏈脫氫酶的影響Fig. 6 Effects of brevilaterin and ε-polylysine on respiratory chain dehydrogenase activity of S. aureus
通過測定呼吸鏈脫氫酶活性來研究抑菌劑對細胞膜代謝活力的影響[26]。紅四氮唑進入細胞后會被呼吸鏈脫氫酶還原為紅色的三苯甲臢,根據其含量確定呼吸鏈脫氫酶的活力。如圖6所示,1/4 MIC抗菌肽組的吸光度與對照組相比大幅降低,MIC抗菌肽組的吸光度最低,1/4 MIC ε-聚賴氨酸與MIC ε-聚賴氨酸組的吸光度均顯著高于對照組,聯用組的吸光度均低于抗菌肽brevilaterin與ε-聚賴氨酸二者單獨使用。結果表明抗菌肽brevilaterin對呼吸鏈脫氫酶有明顯抑制作用,而ε-聚賴氨酸對呼吸鏈脫氫酶無抑制作用。
2.5 抗菌肽brevilaterin與ε-聚賴氨酸聯用對菌體超微結構的影響
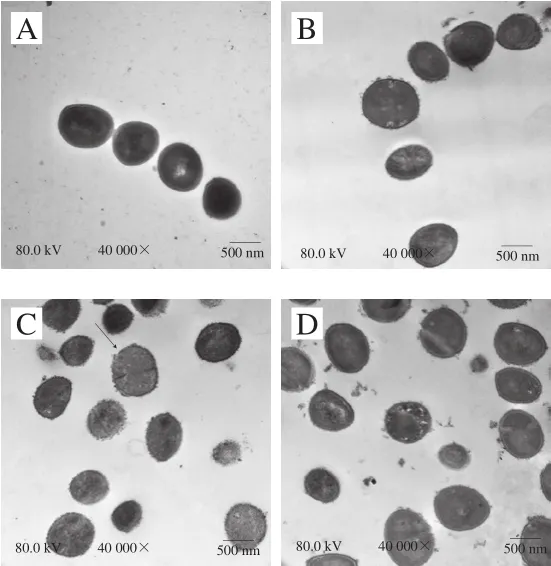
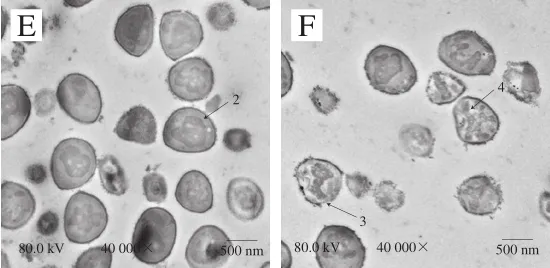
圖 7 抗菌肽brevilaterin與ε-聚賴氨酸對金黃色葡萄球菌超微結構影響的透射電鏡圖Fig. 7 Changes in ultrastructure of S. aureus treated with brevilaterin and ε-polylysine
從圖7可以看出,對照組菌體細胞壁和細胞膜無破損,內容物完整;1/4 MIC抗菌肽brevilaterin、MIC抗菌肽brevilaterin處理后細胞表面均出現破損(箭頭1),MIC組破損較嚴重;1/4 MIC ε-聚賴氨酸處理后小部分細胞出現輕微破損;MIC ε-聚賴氨酸處理后細胞邊緣模糊,部分細胞表面出現破損,細胞質密度均有所降低(箭頭2);聯用組的細胞發生變形破裂(箭頭3),且細胞內容物發生泄漏導致細胞質密度降低(箭頭4)。上述結果表明抗菌肽brevilaterin與ε-聚賴氨酸聯用可協同破壞壁和細胞膜,導致內容物泄漏。結果中ε-聚賴氨酸對細胞超微結構的影響與Hyldgaard等[27]研究的經ε-聚賴氨酸處理后的大腸桿菌細胞變化情況相似,均能使細胞發生形變、細胞內容物發生泄露。
2.6 抗菌肽brevilaterin與ε-聚賴氨酸聯用對細胞蛋白合成的影響
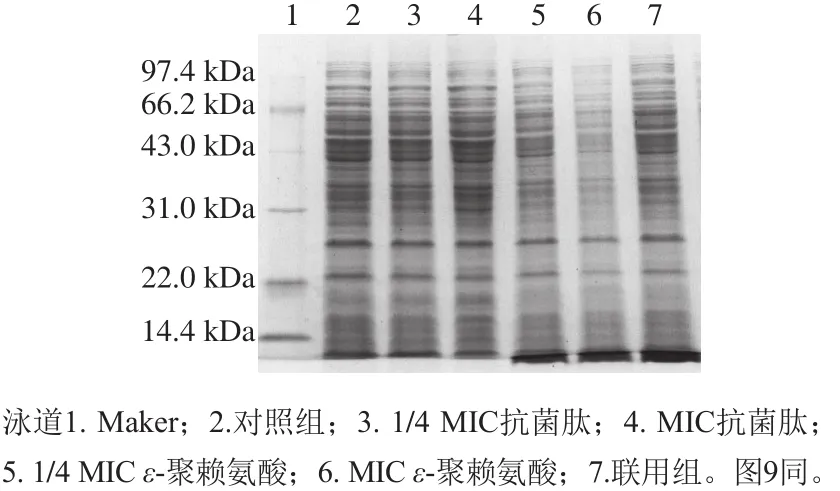
圖 8 抗菌肽brevilaterin與ε-聚賴氨酸對金黃色葡萄球菌蛋白質合成的影響Fig. 8 Effect of brevilaterin combined with ε-polylysine on protein synthesis in S. aureus
從圖8可以看出,抗菌肽brevilaterin處理后的蛋白質條帶與對照組相比無明顯變化;MIC ε-聚賴氨酸處理后蛋白質條帶整體灰度變淺,說明蛋白質濃度降低,且在分子質量小于14.4 kDa處,1/4 MIC ε-聚賴氨酸、MIC ε-聚賴氨酸、聯用組處理后蛋白質條帶灰度加深。上述結果表明,ε-聚賴氨酸對大分子蛋白質的合成有抑制作用或者是能將大分子蛋白質降解為小分子蛋白質。韓晴[28]通過十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳研究了ε-聚賴氨酸對金黃色葡萄球菌蛋白質合成的影響,也發現ε-聚賴氨酸能夠造成小于14.4 kDa處蛋白質條帶灰度加深。
2.7 抗菌肽brevilaterin與ε-聚賴氨酸聯用對菌體DNA的影響
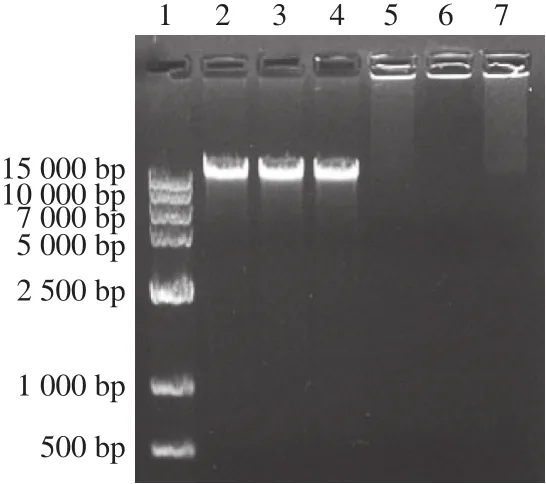
圖 9 抗菌肽brevilaterin與ε-聚賴氨酸聯用對金黃色葡萄球菌DNA的影響Fig. 9 Effect of brevilaterin combined with ε-polylysine on DNA of S. aureus
如圖9所示,與對照組相比,抗菌肽brevilaterin作用后的菌體DNA條帶無明顯變化;1/4 MIC、MIC ε-聚賴氨酸以及聯用處理后,DNA均滯留于上樣孔,可能是因為電荷的電性發生改變,也可能是因為DNA雙螺旋結構發生改變,從而影響DNA遷移率[29]。周祺等[7]研究ε-聚賴氨酸與腸球菌DNA的結合能力時,發現ε-聚賴氨酸與腸球菌DNA有較強的結合能力,可通過與DNA結合進而破壞DNA的功能;Liu Hongxia等[30]也證實了ε-聚賴氨酸與DNA之間具有結合作用。
3 結 論
抗菌肽brevilaterin與ε-聚賴氨酸對金黃色葡萄球菌具有協同抑菌效應。抗菌肽brevilaterin可通過消散細胞跨膜電勢,增加細胞膜通透性,破壞細胞膜完整性,進入細胞內從而抑制呼吸鏈代謝活力最終抑制菌體生長。ε-聚賴氨酸可通過破壞跨膜pH值梯度,增加細胞膜通透性,進入胞內抑制細胞蛋白質合成(或者降解蛋白質),與DNA結合從而抑制細菌生長。抗菌肽brevilaterin與ε-聚賴氨酸聯用兼具兩者單獨作用的抑菌機理,即可通過影響跨膜pH值梯度,增加細胞膜通透性,破壞細胞膜完整性,抑制呼吸鏈代謝活力,與DNA結合影響蛋白質合成從而發揮協同抑菌作用。
