2016年福建省B型流感病毒基因特征分析
2020-04-13 08:42:52陳宏彬張炎華修文瓊翁育偉謝劍鋒
中國人獸共患病學報 2020年2期
陳宏彬,張炎華,林 琦,修文瓊,翁育偉,謝劍鋒
流行性感冒(Influenza)簡稱流感,是流感病毒引起的一種急性發熱性呼吸道傳染病。流感病毒是RNA病毒,病毒顆粒的表面有兩種形態不一、長度均為10~20 nm的糖蛋白突起,即血凝素和神經氨酸酶,均具有抗原性。HA及NA易于發生變異,而且變異是各自獨立的。B型流感病毒在某些特定的情況下所導致的疾病負擔甚至超過A型流感病毒,2013-2014年流感流行季節,在北美地區特別是美國暴發了大范圍B型流感疫情[1]。B型流感病毒與A型相比更容易引發患者發生并發癥, 對兒童、青少年、老人等特定人群易感, 且感染兒童及青少年后引起的病死率較高,給人類的公共衛生健康造成了嚴重威脅[2]。 B型耐藥相關突變株的突變位點多種多樣,且我國的B型流感病毒耐藥情況與國際情況并不十分一致[3]。而B型流感的多樣性和耐藥病毒株越來越多的涌現可能影響公共衛生,有必要對其研究探討以提供防控參考依據。
2016年至2017年北半球季節性流感病毒三價疫苗推薦毒株:A/California/7/2009 (H1N1)pdm09-like virus; A/Hong Kong/4801/2014 (H3N2)-like virus; B/Brisbane/60/2008-like virus。四價流感疫苗含兩種B型流感病毒,除上述3種病毒株外,增加的另一種B型流感病毒為B/Phuket/3073/2013-like virus疫苗株。B型流感病毒HA蛋白的抗原決定簇主要包括了4個區域分別是120環(116~137)、150環(141~150)、160環(160~172)和190螺旋(194~202),其中Victoria系的120環包括了75和77位點[4]。RBS包括190螺旋(193-202)、240環(237-242)、140環(136-143)、Phe-95、Trp-158、His-191、Tyr-202[5]。B型流感耐藥位點包括E117G/A/D/V、R150K、D197E/N/Y、I221V/I/T、H273Y、R292K、N294S、R374K、G407S[6-7]。
1 材料和方法
1.1資料及標本來源 從中國疾病預防控制信息系統采集日期2016年1月1日至2016年12月31日的我省流感監測數據。毒株來源2016年福建省流感網絡監測流感樣病例,犬腎傳代細胞或雞胚分離培養。
1.2病毒核酸的提取 天隆科技(蘇州)有限公司病毒RNA提取試劑盒。
1.3引物設計與合成引物序列 見表1,生工生物工程(上海)股份有限公司合成。(引物來源:http://gsc.jcvi.org/projects/msc/influenza/)
1.4RT-PCR擴增及測序 TaKaRa Prime Script one step RT-PCR試劑盒。反應程序:50 ℃ 30 min,94 ℃ 2 min;94 ℃ 30 s,60 ℃ 30 s,72 ℃ 1 min,30 cycles;72 ℃ 10 min。產物測序由鉑尚生物技術(上海)有限公司完成。
1.5病毒序列分析及進化樹構建 測序用SeqMan軟件拼接,并選取WHO推薦的北半球流感疫苗株和代表株多序列排列比對。Victoria系與北半球流感疫苗株B/Brisbane/60/2008,我國代表株B/Chongqing-Yuzhong/1384/2010; 2006-2007年北半球疫苗株B/Malaysia/2506/2004,2002-2004年北半球疫苗株 B/Hong Kong/330/2001及參照序列 B /Victoria /02/1987(均來自美國 GenBank 數據庫)繪制進化樹;Yamagata系與2015-2016北半球疫苗株B/Phuket/3073/2013,2013-2015北半球疫苗株B/Massachusetts/2/2012,2012-2013北半球疫苗株 B/Wisconsin /01 /2010, WHO 疫苗種子株 B/Hubei-Wujiagang /158 /2009;北半球2008-2009年疫苗株B /Florida /4 /2006;北半球2005-2006 年疫苗株B /Shanghai /361 /2002;北半球2001-2002年疫苗株 B /Sichuan /379 /99及參照序列B /Yamagata /16 /1988(均來自美國 GenBank 數據庫) 繪制進化樹。GASID下載2016年其他省份HA、NA序列。MEGA構建Neighbor-Joining進化樹, Kimura 2-parameter模型繪制核酸系統進化樹,bootstrap設為1000。Net NGlyc 1.0 Server在線分析軟件預測糖基化位點。

表1 B型流感病毒NA/HA基因擴增引物Tab.1 NA/HA gene amplification primers for influenza B virus
2 結 果
2.1福建省2016年流感流行情況分析 2016年福建省流感網絡監測流感樣病例共24 131份,原始標本PCR檢測到流感4 935例;其中Victoria系共1 988例(40.28%),Yamagata系480例(9.7%),季H3共1 368例(27.43%),新甲H1共1 083例(21.95%)。H1N1,H3N2、B型流感交替流行(圖1),上半年B型為主,H1N1其次,下半年H3N2則成為流行優勢株。B型兩個譜系同時流行,冬春季節為發病高峰。8~9月B型流感數量明顯降低,H3N2成為主流。為了解福建省流行的B型流感病毒分子特性,挑取各地市不同時間段51株流感毒株對HA、NA基因進行分析。
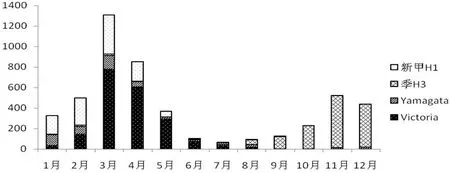
圖1 福建省2016年流感監測不同型別構成圖Fig.1 Different types of influenza surveillance in Fujian Province in 2016
2.2HA/NA同源進化分析 Victoria系的28株HA和31株NA核苷酸序列與B/Brisbane/60/2008疫苗株同源性分別在98.6%~100%和97.8%~100%。Yamagata系的23株HA和20株NA核苷酸序列與B/PHUKET /3073/2013疫苗株同源性分別在98.3%~100%和97.8%~100%。HA基因進化樹分析見圖2,Yamagata系和Victoria系分為2個分支,我國疫苗株與我省及其他省份毒株在同一分支,距離近,無重配發生。NA基因進化樹圖3也分為Yamagata系和Victoria系,與疫苗株在同一分支上,與同年其他省份距離近在同一分支上,有2株重配株(BY-NA/BV-NA)。2016年流感與早期B型毒株NA距離較遠,已經形成新的分支。
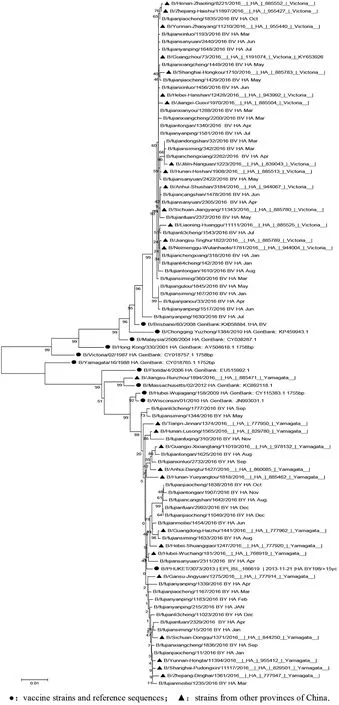
●:vaccine strains and reference sequences; ▲:strains from other provinces of China.圖2 2016年福建省B型流感病毒HA基因進化樹Fig.2 Evolution of HA gene of influenza B virus in Fujian Province in 2016
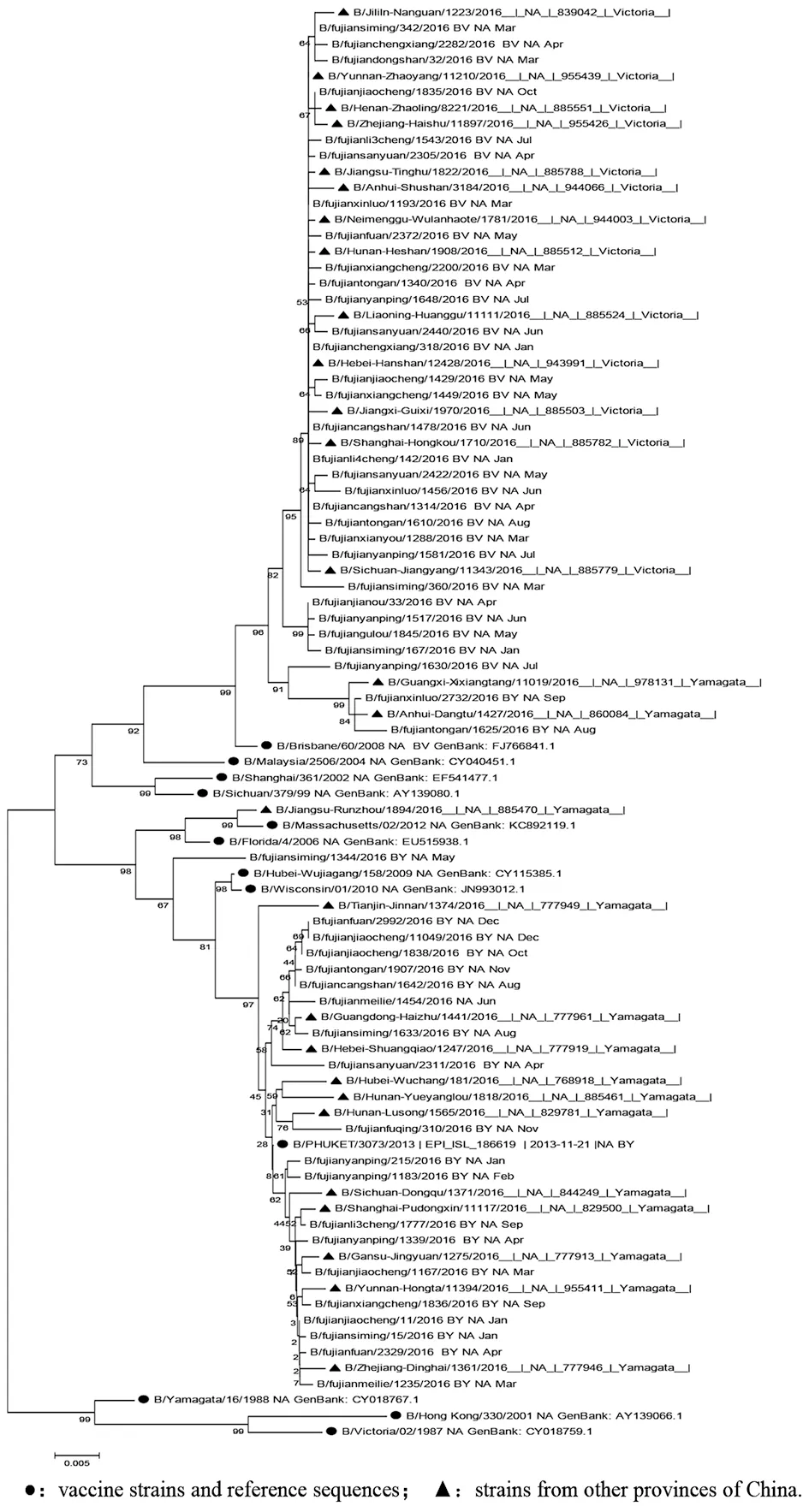
●:vaccine strains and reference sequences; ▲:strains from other provinces of China.圖3 2016年福建省B型流感病毒NA基因進化樹Fig.3 Evolution of NA gene of influenza B virus in Fujian Province in 2016
2.3HA變異分析 HA基因變異只顯示抗原決定簇上變異位點(表2、3)。Victoria系與疫苗株B/Brisbane/60/2008相比,HA共有16個變異位點,大部分毒株存在I117V,N129D,A199T變異,涉及到120環(117、129)、190螺旋(197,199)也是RBS、150環(2株)其中RBS(1株)。117位點有4株無變異,而這4株有T189A變異。只1株117,129均無變異,而這株有I146V,K209N變異。Yamagata系與疫苗株B/PHUKET/3073/2013相比,HA共有19個變異位點,均存在L172Q、M251V變異,大部分毒株存在K211R,涉及到120環(2株)、150環(1株)、RBS+190螺旋(2株); 5株有29(V-A)或275(A-V)變異,無此變異的毒株有K211R變異。
2.4NA變異分析 2016年福建省51株B型流感NA發現耐藥位點D197N(1株),余未見耐藥位點。NA酶活性區域由功能部位8個氨基酸R-118、D-151、R-152、R-224 、E-276、R-292、R-371和 Y-406 和結構部位11個氨基酸 E-119、R-156、W-178、S-179、D-198、I-222、 E-227、H-274、E-277、N-294和 E-425(以上位點基于N2編碼),在A型和B型流感病毒中均為保守氨基酸[8]。NA酶活性區域或者結構部位發生構象變異的突變會影響病毒對NAI的敏感性,51株B型流感1株D198N變異,余未發現以上活性區變異。
Victoria系與疫苗株B/Brisbane/60/2008相比,NA共37個變異位點,I120V,K220N,S295R,N340D,E358K, D384G在大部分毒株存在; Yamagata系與疫苗株B/PHUKET/3073/2013相比NA共27個變異位點,變異位點零散。除了1株其余都發現有I49T或A62T位點變異,而沒有2個位點變異的這株發生了A67V變異。P100S、I115V兩位不同時發生變異或都無變異。類似情況還見于S295R、M403I兩個位點,除了1株(思明1344)同時發現S295R、M403I兩個位點變異,而這株獨有G70E、R186K、D235N、I248V、K320D變異,變異位點(9)最多,與疫苗株不在同一分支。
2.5糖基化位點分析 28株Victoria系HA存在25、59、145(抗原決定簇150環)、166(抗原決定簇160環)、197(4株除外,抗原決定簇190螺旋和RBS)、233、304、333、492、518、531、563共11-12個糖基化位點。與疫苗株相比多1個糖基化位點197(4株除外)。23株Yamagata系HA存在25、59、145(抗原決定簇150環)、167(抗原決定簇160環)、196(1株除外,抗原決定簇190螺旋和RBS)、303、332、491、517、530、562共10-11個糖基化位點。與疫苗株一致。29株Victoria系NA存在56、64、144、284共4個糖基化位點。與疫苗株一致。22株Yamagata系NA存在56、64、144、284、463(2株重配株除外)共4-5個糖基化位點。與疫苗株一致。

表2 福建省2016年B型Victoria系流感HA基因變異情況Tab.2 Variation of HA gene in influenza B Victoria in Fujian Province in 2016

表3 福建省2016年B型Yamagata系流感HA基因變異情況Tab.3 Variation of HA gene in influenza B Yamagata in Fujian Province in 2016
3 討 論
B型流感和A型一樣受到了很強的人群免疫壓力,病毒依靠外膜蛋白,特別是血凝素重鏈區(HA1)的變異,來改變抗原特性從而逃避機體固有免疫[9]。最近流行的病毒包括Victoria和Yamagata兩系相對熵高的HA殘基最常位于120環區域[10],本文研究中Victoria系毒株HA變異大多數涉及到120環,表位區域外的一些變異位點有待進一步研究。Yamagata和Victoria的HA變異方式不同,Victoria的變異主要涉及到2-3個抗原決定簇以及RBS,而Yamagata抗原決定簇變異的少,這兩系可能以不同方式進行變異累積。
2016年福建省Yamagata系HA的糖基化位點與疫苗株相比只1株少1個糖基化位點,其他糖基化位點一樣。Victoria系HA的糖基化位點與疫苗株相比只4株沒有增加1個糖基化位點。HA糖基化位點的增減及位置變化,HA上糖鏈結構、數量的變化以及受體結合位點附近糖鏈的修飾等不僅對病毒的增殖、毒力產生影響,還會影響病毒與宿主細胞受體的結合能力等[11]。病毒在進化過程中也會獲得新的糖基化位點,這些新積累的糖基化位點可能會掩蓋抗原位點,成為病毒逃避宿主免疫壓力的一種方式[12]。Victoria系增加的糖基化位點可能是2016年Victoria系在福建省流感中占多數的原因之一。
目前,一線抗流感病毒藥物神經氨酸酶抑制劑已在臨床上廣泛上使用,甲型和乙型流感病毒對臨床上常用的耐藥率并不高(保持在2%以內[13]),但亦有研究表明耐藥株容易在兒童、65歲以上老人和免疫功能低下者體內分離出[14]。51株B型流感病毒均未發現以上活性區變異,只1株耐藥株,其他50株敏感提示我省B型流感對NAI的敏感性未發生明顯改變,可推測福建省目前流行B型流感對神經氨酸酶抑制劑類藥物敏感。國家疾控中心推斷目前我國流行的B型流感毒株對神經氨酸酶抑制劑類藥物敏感[15],與本研究結論一致。雖然B型流感的2個種系在進化上存在差異,但二者可在人群中同時流行增加發生基因重配的機會。從進化樹上看重配的病毒與疫苗株距離較遠,重配是否影響B型流感病毒的生長特性、毒力等需進一步的研究。
在抗原決定簇區外的一些位點也發生了變異,除了目前已知的抗原決定簇區域可能還存在其他的變異區[9]。變異如何影響病毒的抗原性需進一步研究,增加對流感病毒進化規律的認識能幫助我們更好的有效預防和治療。多方研究報道均提示病毒蛋白上的一些位點在進化過程中會相互聯系、相互協同;一種簡單的情形就是:同一個蛋白一對正選擇位點在插曲式的正選擇作用下,或者保持穩定,或者同時突變(如 PB1 蛋白52 位于 576 位)[16]。本研究發現的一些變異位點可為研究其功能相關性提供基礎。
有研究表明一旦H3N2病毒離開東南亞地區,病毒就無法進行長期的進化,假如流行病毒是有規律的從外地輸入,如果種子株輸入源可以確認,就可能預測出哪個變異株將在流行區域出現,而隨之優化疫苗株的選擇[17]。國家疾控中心提示BY系個別位點變異主要來自福建省,多數低反應毒株分離自南方省份,要高度關注南方省份Yamagata系病毒的抗原性和基因特性變異情況[18]。進化樹圖可看出我省與其他省份的毒株并不能區分開,可能為同一個傳染源。大部分B型流感同源性高差異不明顯,但仍有個別株差異較大。由于個體中流感免疫反應的復雜性與多次暴露于不同毒株有關,不同接觸史的宿主可能會繼續經歷不同的感染模式,這可能會影響流感病毒持續的抗原進化[19]。推測HA的單適應性突變也與近幾年疫苗效果不佳有關,而NA基因未受疫苗的影響或許可為下一代疫苗即構建一個同時含有可表達HA和NA蛋白的重組株的研究提供參考[20]。因此本文分析2016年福建省B型流感病毒HA和NA變異特征為我省疫情防控提供參考依據。
利益沖突:無
引用本文格式:陳宏彬,張炎華,林琦,等.2016年福建省B型流感病毒基因特征分析[J].中國人獸共患病學報,2020,36(2):118-123,157. DOI:10.3969/j.issn.1002-2694.2019.00.176
