水稻輪回選擇群體XTBG-HP1表型遺傳多樣性分析
2020-04-17 08:58:20唐如玉徐鵬余迪求
廣西植物 2020年2期
關鍵詞:水稻
唐如玉 徐鵬 余迪求


摘要:? 該研究基于4個陸稻群體及172個水稻品種或雜交組合,構建了水稻多親本隱性核不育輪回選擇群體XTBG-HP1,并經過4次輪回重組,采用16個表型性狀對其進行了遺傳多樣性分析。結果表明:(1)該群體14個數量性狀符合正態分布,各表型均存在極端性狀個體。(2)數量性狀變異系數范圍為0.08~0.41,均值為0.20;Shannon-Wiener多樣性指數范圍為0.72~1.92,均值為1.50。(3)群體在株型與產量構成因子性狀方面有顯著的相關性,對株型的選擇可以實現產量性狀的改良。(4)劍葉長、每穗粒總數、千粒重、穗長、粒長、一次枝梗數、有效穗數、劍葉寬、二次枝梗數、抽穗期10個性狀可作為群體綜合評價指標。(5)劍葉長、二次枝梗數、每穗粒總數3個表型性狀具有較高的遺傳變異、豐富的遺傳多樣性及與綜合得分F值相關系數較高。綜合以上結果發現,后期群體進行基因挖掘、品種改良以及優良育種材料的選育可以基于劍葉長、二次枝梗數及每穗粒總數3個表型性狀,同時要充分利用群體株型與產量構成因子性狀間的顯著相關性。此外,該研究群體中極端單性狀或綜合得分F值較高的個體,可進一步用于品種選育。
關鍵詞: 水稻, 輪回選擇, 表型性狀, 群體遺傳多樣性, 群體改良
中圖分類號:? Q943文獻標識碼:? A
文章編號:? 1000-3142(2020)02-0159-14
Abstract:? Genetic diversity is the basis of gene mining, cultivars improvement and breeding, the genetic diversity analysis of recurrent selection population could provide reference and guidance for further population improvement and elite cultivars screening. In this study, we constructed the rice recurrent selection population XTBG-HP1 based on four upland rice populations and 172 rice cultivars or combination, and we evaluated the genetic diversity of the rice population, by using sixteen phenotypic traits after four recurrent combinations. The results were as follows: (1) The fourteen quantitative traits of study population were normal distribution, and there were extreme trait individuals in each phenotype. (2) The coefficient variation of fourteen quantitative traits ranged from 0.08 to 0.41 with a mean of 0.20. Shannon-Wiener diversity index of each trait ranged from 0.72 to 1.92 with a mean of 1.50. (3) There was significant correlation between plant type traits and yield components traits in population, indicating the selection of population plant type could improve population yield traits. (4) The ten traits of flag leaf length, total number of per panicle, 1 000-Grain weight, panicle length, grain length, primary branch number, effective panicle, flag leaf width, the secondary branch number and heading date could be used as indicators for population comprehensive evaluation. (5) The flag leaf length, the secondary branch number and total number of per panicle with high genetic variation, better genetic diversity and high correlation coefficient with composite scores F value. The results of comprehensive experiment analysis showed that the research population has abundant genetic diversities. The late population gene mining, selection of good breeding materials and varieties improvement can be place emphasis on flag leaf length, the secondary branch number, and total number of per panicle, and make the best of the correlation between plant type traits and yield components traits. In addition, individuals with extreme single trait or higher F value in the study population can also be further used for cultivars improvement.
Key words: rice, recurrent selection, phenotypic traits, population genetic diversity, group improvement
水稻是世界上主要的糧食作物,全球有50%以上的人以水稻為主食,在全世界廣泛種植(Ansari et al., 2017)。在水稻育種史上,水稻產量經歷了兩次飛躍:一次是從20世紀50年代末至70年代初,以半矮化品種應用為特征的水稻綠色革命使世界水稻增產2倍(Jain,2012);另一次是20世紀70年代至80年代,中國率先采用雜交水稻育種技術,使水稻產量潛力大約提高了25%(Peng et al., 2008)。此后,新型超高產水稻育種,取得了顯著進展(Cheng et al., 2007;Peng et al., 2008)。然而,近十年來,水稻產量徘徊不前,似乎達到上限(Ansari et al., 2017)。世界人口數量的持續增長、可用耕地面積和灌溉水資源的減少以及環境變化等都對水稻增產穩產提出了更高的要求和挑戰(Ansari et al.,2017)。傳統育種由于雜交方式和雜交手段的限制,所以育成品種存在綜合親本少、遺傳基礎狹窄等問題,而以傳統育種方法育成的不育系和恢復系本身存在上述問題,也反過來影響育種進程(Li & Zhang,2013;Müller et al., 2017)。此外,由于水稻育種對相同優異骨干親本持續利用及品種推廣的單一化,加之集中對雜交后代的關注性狀進行選擇,選育過程中必然會造成大量優良基因損失及遺傳基礎狹窄,進而造成水稻單產徘徊不前和抗性的遺傳脆弱性(胡標林等,2012)。水稻大部分重要農藝性狀均屬于數量性狀,受到多個微效基因的共同調控。此外,水稻眾多基因間存在連鎖現象,傳統雜交育種方式難以對這些基因同時進行改良。隨著水稻眾多重要功能基因的克隆和分子解析以及分子生物學技術的應用(Jiang et al., 2012),水稻育種必然會朝著高產、多抗、優質、廣泛適應性的目標發展(萬建民,2007;Li & Zhang,2013)。因此,對多基因、多性狀同時改良的育種方式成為水稻育種發展的必然選擇。
輪回選擇基于遺傳基礎豐富的群體,經過多次選擇、重組、選擇,可打破基因間的連鎖,增加有利基因重組機會,從而有效整合分布于不同親本的優異性狀,在改良群體的同時保持廣泛的遺傳多樣性(漆映雪和鄒小云,2008),這是一種理想的育種方式。輪回選擇育種尤其適用于數量性狀的改良,如產量、抗性等重要農藝性狀(Peng et al., 2008)。早在1919年,Hayes等就提出了輪回選擇的概念(夏軍紅和鄭用鏈,2002)。由于雜交方式的限制,輪回選擇最早用于異花授粉作物玉米(Bolaos et al., 1993)。隨著雄性不育株的發現,輪回選擇得以在自花授粉作物中應用。目前,輪回選擇已經在自花授粉作物大豆(Brim & Stuber,1973;Posadas et al., 2014)、小麥(Marais et al., 2010;Ramya et al., 2016)及水稻(Fujimaki,1979;Frouin et al., 2014;Grenier et al.,? 2015)上廣泛應用。
水稻雄性不育株的發現,促進了輪回選擇在水稻上的應用(Grenier et al., 2015;Morais et al., 2017;Pang et al., 2017)。Morais et al.(2017)選取株高和產量兩個性狀,從基因和表型兩方面對旱稻群體CNA6輪回選擇群體4個周期的遺傳變異情況進行分析,結果表明在4個輪回選擇周期中,株高和產量性狀都保持較好的遺傳多樣性。Morais et al.(2017)選取產量、株高、開花天數3個性狀,通過對水稻群體進行3次輪回選擇,從遺傳進程、遺傳變異、優選品系潛力三方面對輪回選擇效率的研究表明,輪回選擇具有促進遺傳進程,維持群體遺傳多樣性,提高群體對單性狀、雙性狀及三性狀提取優勢系的遺傳潛力,是一種有效的育種方式。
遺傳多樣性研究的方法較為完善,主要有基于形態學的標記、細胞學的標記、生理生化標記以及分子標記等。表型性狀是水稻育種和進行復雜性狀機理解釋的重要依據,能通過最直接的外觀性狀反映遺傳,與微衛星標記具有很高的相似性(齊永文,2004)。當研究群體較大時,表型性狀研究最為簡單、經濟,應用十分廣泛,常用于遺傳育種工作中(胡標林等,2012;王海崗等,2016)。李自超等(2001)和陳越等(2019)通過表型性狀,對云南水稻種質資源進行了分析,結果發現云南省稻種資源具有豐富的遺傳多樣性和表型多樣性。胡標林等(2012)選用14個表型性狀,對美國農業部水稻核心種質表型性狀遺傳多樣性進行了分析和綜合評價。
輪回選擇是改良群體、維持群體遺傳多樣性及篩選優良個體的有效育種方法。目前,水稻輪回選擇育種方法及表型遺傳多樣性分析方法均較為成熟,為水稻輪回選擇群體的構建及其遺傳多樣性分析提供了較好的基礎。本研究通過16個表型性狀對重組4次的水稻輪回選擇群體XTBG-HP1的表型遺傳多樣性進行分析,從而明確群體的遺傳多樣性,為研究群體的進一步選育及優良個體的篩選提供方向和指導。
1材料與方法
1.1 材料
利用4個引自哥倫比亞國際農業研究中心(CTAT)含有隱性核不育基因的陸稻群體SPACIR14、SPACIR16、SPACIR17、SPACIR18中的不育株作為母本,4個陸稻群體的親本信息及來源如表1所示。172個水稻品種或雜交組合作為重組親本,親本信息如表2所示,用于構建水稻多親本隱性核不育輪回選擇群體XTBG-HP1。4個陸稻群體不育株不育性來源于IR36的EMS突變體中的隱性核不育基因TDR(Frouin,2014)。
1.2 研究群體構建
水稻輪回選擇群體XTBG-HP1的構建流程與方法如圖1所示。大量種植陸稻群體,172個重組親本分別與陸稻群體的每5株不育株進行雜交(親本信息見表2),共獲得2 827株F1(部分親本只獲得1~4個不育單株雜交),種植F1并等量混合收取F1種子,獲得S0群體,命名為XTBG-HP1。隔離條件下單株種植S0群體(面積大于2 000 m2),等量收取分離不育株上的重組種子,種植重組種子獲得S1重組群體,相同方法經過4次群體重組,得到S4輪回重組群體。在S4輪回重組群體中隨機收取2 000株可育株種子,單粒傳種植得到F2群體,最終群體的有效數量為1 395株,作為本研究材料。
1.3 試驗方法
2018年8月,在中國科學院西雙版納熱帶植物園水稻育種試驗基地單苗種植F2群體,催芽育苗至三葉一心期移栽,株行距20 cm × 30 cm,常規種植管理。參考GB/T19557.7-2004《植物新品種特異性、一致性和穩定性測試指南-水稻》標準,對試驗群體表型性狀進行調查,并記錄各指標。除了抽穗期于抽穗時調查以外,其他性狀均于成熟期調查。總共調查16個表型性狀,包括抽穗期、有效穗數、莖粗、株高、劍葉角度、劍葉長、劍葉寬、一次枝梗數、二次枝梗數、每穗粒總數、穗型、穗長、粒長、粒寬、粒長寬比、千粒重。對劍葉角度、穗型進行定級量化,劍葉角度分為直立、中間、平展、披垂,穗型分為密集、中間型、散開,它們都分別用數字1、2、3、4來表示。
1.4 數據分析
采用Microsoft Excel 2013進行數據的基本整理和分析。對所有表型性狀數據進行標準化處理,即將各性狀定義到 [0,1]閉區間。
主成分分析。先基于主成分分析計算出各主成分得分,再利用逐步回歸分析篩選指標性狀。
2結果與分析
2.1 表型性狀分布及變異分析
2.1.1 質量性狀研究群體劍葉角度包括4種類型(圖2:A),即59.21%為直立類型、40.00%為中間型、0.65%為平展型、0.14%為披垂型,以直立和中間類型為主。穗型包括3種類型(圖2:B),即5.58%為密集型、29.15%為中間型、65.00%為散開型,以散開型為主。
2.1.2 數量性狀由研究群體14個數量性狀分布情況(圖3:A-N)可知,14個數量性狀均符合正態分布。各表型均存在極端差異性狀,表明該群體內的個體可能進行了較充分的重組。
表型性狀變異系數,可反應不同群體的均度。通過計算該群體14個數量性狀的均值、標準差計算出相應變異系數,由表3可知群體的變異情況。群體抽穗期變幅為42~109 d,均值為71.54±8.22,變異系數為0.12。有效穗數的變幅為1~22個,均值為6.98±2.85,變異系數為0.41。莖粗變幅為0.80~16 mm,均值為5.51±1.18,變異系數為0.22。株高變幅為34~187 cm,均值為113.25±20.99,變異系數為0.19。劍葉長變幅為11.20~76.40 cm,均值為34.97±7.92,變異系數為0.23。劍葉寬變幅為0.60~2.80 cm,均值為1.56±0.30,變異系數為0.19。穗長變幅為8.50~41 cm,均值為25.52±3.76,變異系數為0.15。一次枝梗數的變幅為1~33個,均值為11.77±2.56,變異系數為0.22。二次枝梗數的變幅為5~110個,均值為44.74±15.16,變異系數為0.34。每穗粒總數的變幅為34~462粒,均值為214.92±71.34,變異系數為0.33。粒長變幅為7.60~15.22 mm,均值為9.83±0.91,變異系數為0.090。粒寬變幅為2.28~4.87 mm,均值為2.95±0.24,變異系數為0.080。粒長寬比變幅為2.19~4.87,均值為3.35±0.39,變異系數為0.12。千粒重的變幅為13.61~39.40 g,均值為25.27±3.59,變異系數為0.14。群體14個數量性狀的變異系數范圍為0.080~0.41,均值為0.20,說明群體具有豐富的遺傳變異。其中:有效穗數(0.41)的變異系數最大;其次分別為二次枝梗數(0.34)、每穗粒總數(0.33);變異系數最小的分別為粒寬(0.080)、粒長(0.090)。
2.2 表型性狀多樣性指數
通過對16個性狀進行標準化處理,將各表型性狀分為10個等級(圖4)。由圖4可知,除了劍葉角度、穗型2個定級描述的性狀,以及莖粗第9級、粒寬第8級以外,其他各性狀在各等級上均有分布。抽穗期主要集中分布于4~6級,占群體84.95%。有效穗數主要集中分布于2~4級,占群體75.05%。莖粗主要集中分布于3~4級,占群體83.23%。株高主要集中分布于4~7級,占群體86.45%。劍葉角度主要集中分布于1~2級,占群體99.21%。劍葉長主要集中分布于3~5級,占群體78.06%。劍葉寬主要集中分布于4~5級,占群體62.01%。穗長主要集中分布于5~7級,占群體81.65%。穗型主要集中分布于2~3級,占群體94.15%。一次枝梗數主要集中分布于3~5級,占群體97.66%。二次枝梗數主要集中分布于3~5級,占群體72.45%。每穗粒總數主要集中分布于3~6級,占群體83.40%。粒長主要集中分布于2~4級,占群體87.26%。粒寬主要集中分布于2~4級,占群體91.84%。粒長寬比主要集中分布于3~6級,占群體70.28%。千粒重主要集中分布于4~6級,占群體74.19%。為了研究各表型的遺
傳多樣性和分布頻率的平衡性,采用Shannon-Wiener多樣性指數計算群體各表型性狀的多樣性指數。多樣性分析結果(圖5) 表明, 各表型性狀的多樣性指數分布范圍為0.72~1.92,均值為1.50。其中:每穗粒總數(1.92)的多樣性指數最高;其次為二次枝梗數(1.85);最低的分別為劍葉角度(0.72)、穗型(0.81)兩個質量性狀。數量性狀中,以莖粗(1.19)、一次枝梗數(1.31)的多樣指數最低。除了莖粗(1.19)、一次枝梗數(1.31)、粒寬(1.34)幾個性狀的遺傳多樣性指數低于均值以外,其他性狀的遺傳多樣性指數都接近或高于均值,這更進一步說明了該群體的表型多樣性豐富。
2.3 表型相關性分析
運用SPSS軟件,對14個數量性狀進行相關性分析結果(表4)表明,14個表型性狀間存在不同程度的相關性,且大部分為顯著或極顯著相關。其中,株高與有效穗數、莖粗、劍葉長、劍葉寬、穗長呈極顯著正相關,且與穗長(0.58)、莖粗(0.43)兩者的相關系數較高,說明株高選育要兼顧穗長與莖粗兩種性狀。劍葉長、劍葉寬均與莖粗、穗長呈極顯著正相關,說明對劍葉長寬性狀的選擇可以同時兼顧莖粗、穗長性狀的選擇。穗長與有效穗數、莖粗呈極顯著正相關,說明穗長性狀的改良要兼顧有效穗數和莖粗兩種性狀。二次枝梗數僅與一次枝梗數呈極顯著正相關, 且相關系數較高(0.55),說明二次枝梗數的改良,只能從一次枝梗數著手。粒長寬比與粒長、粒寬分別呈極顯著正相關、負相關,與粒長的相關性系數高于粒寬,且與千粒重呈負相關。此外,千粒重與粒長(0.27)、粒寬(0.52)均呈極顯著正相關,且與粒寬的相關系數大于粒長,說明選育高產品種應更加注重粒寬。以上相關性狀中,株高、莖粗、劍葉長、劍葉寬屬于株型性狀,有效穗數、千粒重等屬于產量相關性狀,這表明株型性狀、產量性狀兩類性狀因子內及相互之間具有相關性。
2.4 表型主成分分析
主成分分析的KMO值為0.73,在設置特征值為0.8時,主要包括7個主成分。由主成分得分系數矩陣(表5)可知,前7個成分的累積貢獻率達79.34%,第1主成分的貢獻率為24.47%。劍葉長、二次枝梗數、每穗粒總數得分較高,表明第1主成分由劍葉長、二次枝梗數、每穗粒總數組成。第2主成分的貢獻率為13.38%,株高和莖粗的得分較高,說明第2主成分主要是由莖粗、株高組成。第3主成分的貢獻率為10.26%,粒寬、千粒重得分較高,說明第3主成分主要由粒寬和千粒重組成。第4主成分的貢獻率為8.73%,粒長得分較高,說明第4主成分主要由粒長組成。第5主成分的貢獻率為7.54%,一次枝梗數的得分較高,第5主成分主要由一次枝梗數組成。第6主成分的貢獻率為6.80%,抽穗期的得分較高,說明第六主成分由抽穗期組成。第7主成分的貢獻率為6.17%,有效穗數的得分較高,說明第7主成分由有效穗數組成。主成分分析結果表明,該群體7個主成分包含了劍葉長、二次枝梗數、每穗粒總數、株高、莖粗、粒寬、千粒重、粒長、一次枝梗數、抽穗期、有效穗數共11個性狀因子。
2.5 群體表型性狀的綜合評價
將研究群體13個表型性狀的標準化值,代入上述7個主成分,求得研究群體的7個主成分得分。其中,第1主成分線性方程如下:
先利用模糊隸屬函數將7個主成分得分歸一化處理,再計算各主成分權重系數(0.33、0.17、0.13、0.11、0.10、0.090、0.080),最后求得群體的綜合得分F值,進而對群體材料綜合評價。群體平均F值為-0.000 15,其中綜合得分F值大于均值的占群體總量的48.30%, 群體中含有較高的優良個體比例。
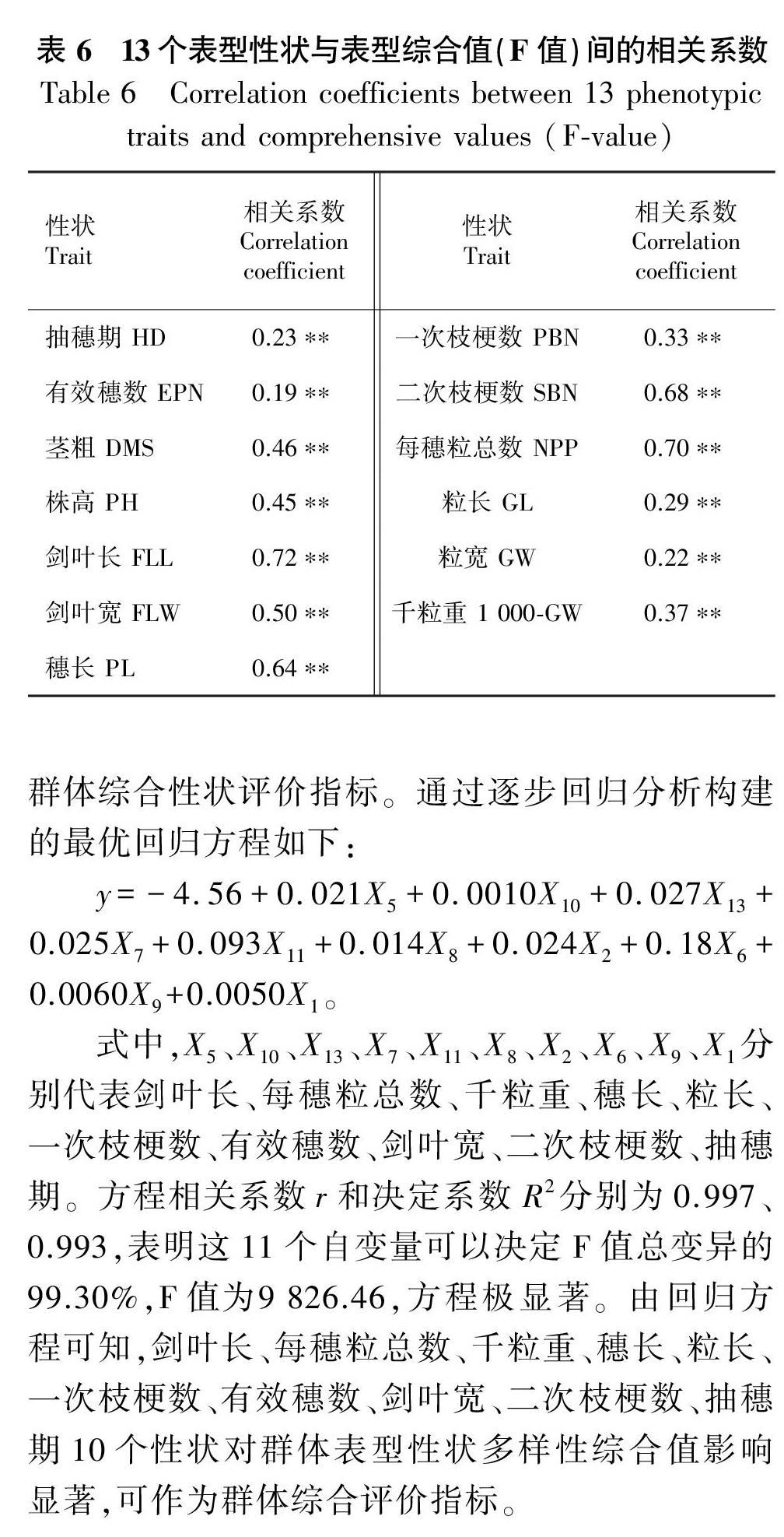
相關性分析結果(表6)表明,F值和各表型性狀均為極顯著正相關。利用綜合得分F值和13個表型性狀構建最優回歸方程。利用已求得的綜合得分F值和表型性狀構建最優回歸方程, 篩選群體綜合性狀評價指標。通過逐步回歸分析構建的最優回歸方程如下:
式中,X5、X10、X13、X7、X11、X8、X2、X6、X9、X1分別代表劍葉長、每穗粒總數、千粒重、穗長、粒長、一次枝梗數、有效穗數、劍葉寬、二次枝梗數、抽穗期。方程相關系數r和決定系數R2分別為0.997、0.993,表明這11個自變量可以決定F值總變異的99.30%,F值為9 826.46,方程極顯著。由回歸方程可知,劍葉長、每穗粒總數、千粒重、穗長、粒長、一次枝梗數、有效穗數、劍葉寬、二次枝梗數、抽穗期10個性狀對群體表型性狀多樣性綜合值影響顯著,可作為群體綜合評價指標。
3討論
遺傳多樣性越豐富,物種對環境變化的適應能力越強,就作物育種而言用于改良栽培品種和選育新品種的潛力就越大。變異系數的大小可以反映群體性狀的遺傳變異程度,Shannon-Wiener多樣性指數能直觀表示生物群落的多樣性。變異系數及多樣性指數常用于作物種質資源(李自超等,2001;胡標林等,2012;王海崗等,2016)及輪回選擇群體遺傳多樣性分析(Morais? JOP et al., 2017;Morais OP et al., 2017)。李自超等(2001)對云南不同地區稻種資源的多樣性研究結果表明,平均Shannon-Wiener指數為1.203 6~1.273 5。賀治州等(2014)通過15個表型性狀對熱帶優異種質資源進行遺傳多樣性分析,數量性狀變異系數范圍為0.070~0.24,均值為0.12,Shannon-Wiener多樣性指數范圍為0.603~2.066,均值為1.579。本研究中,研究群體具有較大的遺傳變異(變異系數范圍為0.080~0.41、均值為0.20),且遺傳多樣性豐富(Shannon-Wiener指數分布范圍為0.72~1.92、均值為1.50)。這與本研究群體在構建之時,引入了大量國內外的種質材料,親本間遺傳多樣性豐富以及輪回選擇育種方式在保持群體遺傳多樣性的同時,能通過基因間的重組,產生重組后代有關。
本研究中,群體劍葉角度與穗型的Shannon-Wiener指數最低,且明顯低于其他數量性狀,這可能與其屬于分級描述的質量性狀,存在較小的變異范圍有關。群體有效穗數、劍葉長、二次枝梗數、每穗粒總數的變異系數、多樣性指數均高于其對應均值,表明研究群體的這四個性狀具有較大的遺傳變異及豐富的遺傳多樣性。抽穗期、穗長、粒寬變異系數、多樣性指數均低于其對應均值,表明研究群體的這幾個性狀遺傳多樣性較低。
水稻表型繁多,且彼此間存在著各種關聯。輪回選擇能打破不利基因間的連鎖,利于群體優良性狀的重組(漆映雪和鄒小云,2008)。這幾組表型性狀的相關性,可能為之后群體性狀的改良提供依據。本研究綜合各性狀相關性分析,莖粗、株高、劍葉長、劍葉寬為水稻株型決定因素,有效穗數、千粒重為產量決定因素,綜合株型及產量性狀相關性分析表明,對株型選擇可以兼顧產量相關性狀。
本研究中,表型主成分分析及綜合評價結果均表明,劍葉長、每穗粒總數、千粒重、穗長、粒長、一次枝梗數、有效穗數、劍葉寬、二次枝梗數、抽穗期10個性狀可作為群體綜合評價的指標,這些性狀可以大致分為株型及產量構成因子兩類。株型及產量構成因子是水稻產量的重要決定因素,對群體遺傳變異、遺傳多樣性及綜合評價主要基于這兩類性狀,也與實驗群體側重于產量性狀分析,以及實驗性狀的選取上相一致。
本研究中,群體各表型性狀均存在極端個體,其中極端性狀個體可能是經輪回重組選擇后多基因累加產生的超親本個體,多種優良性狀的聚合體將是育種選擇的重要基礎。綜合得分F值,能反映相應個體10個指標性狀的綜合表現,F值越高,綜合表現越好,本研究群體中鑒定出的一些高F值個體,可直接用于后期品種選育。與F值相關極顯著的13個表型性狀中,劍葉長(0.72)、每穗粒總數(0.70)、二次枝梗數(0.68)3個性狀相關系數相對較高,說明這幾個性狀對群體評價較為重要,在群體進一步選育及優良個體的篩選中應注重這些性狀的選擇。群體這幾個性狀同樣具有較高的變異系數和Shannon-Wiener多樣性指數,這進一步表明本研究結果具有較好的可靠性。
本研究表型性狀分析表明,水稻輪回選擇群體XTBG-HP1具有較好的遺傳多樣性。后期群體的改良可以側重劍葉長、二次枝梗數、有效穗數等3個具有較高遺傳變異、較好遺傳多樣性及與綜合得分F值相關系數較高的3個表型性狀。此外,在群體的改良上,還要注意株型與產量構成因子性狀間的相關性,并對這一表型相關性加以利用。各表型極端性狀或綜合得分F值較高的個體,可用于后期基因挖掘、品種培育及篩選。陸稻群體及親本,包括產量、稻瘟病抗性、抗旱、陸生適應性、米質等多種優異農藝性狀。本研究主要側重于產量方面對群體表型性狀遺傳多樣性進行了相關分析,之后可以針對其他性狀進行相關分析。由于輪回選擇每次構建的群體均為一個動態群體,本研究說明了該輪回選擇群體的遺傳多樣性,之后可以對輪回選擇群體重組的過程進行一個動態分析,以便對輪回選擇育種過程中群體遺傳多樣性變化有更好的了解。此外,水稻大部分農藝性狀均屬于數量性狀,難免受環境條件的影響。結合SSR分子標記的遺傳多樣性分析,可以更加準確、全面地反應群體的遺傳多樣性。
參考文獻:
ANSARI A, WANG CL, WANG J, et al., 2017. Engineered dwarf male-sterile rice: A promising genetic tool for facilitating recurrent selection in rice? [J]. Front Plant Sci, 8(13): 1-11.
BOLAOS J, EDMEADES GO, MARTINEZ L, 1993. Eight cycles of selection for drought tolerance in lowland tropical maize Ⅲ responses in drought-adaptive physiological and morphological traits? [J]. Field Crop Res, 31(3): 269-286.
BRIM CA, STUBER CW, 1973. Application of genetic male sterility to recurrent selection schemes in soybeans? [J]. Crop Sci, 13(5): 528-530.
CHENG SH, ZHUANG JY, FAN YY, et al., 2007. Progress in research and development on hybrid rice: A super-domesticate in China? [J]. Ann Bot, 100(5): 959-966.
CHEN Y, DING ML, ZHANG DY, et al., 2019. Phenotypic diversity analysis and comprehensive evaluation of agronomic characters of rice germplasm resources in Yunnan Province [J]. J S Agric, 50(9): 1922-1930. [陳越, 丁明亮, 張敦宇, 等, 2019. 云南水稻種質資源農藝性狀表型多樣性分析及綜合評價格[J]. 南方農業學報, 50(9): 1922-1930.]
FROUIN J, FILLOUX D, TAILLEBOIS J, et al., 2014. Positional cloning of the rice male sterility gene ms-IR36, widely used in the inter-crossing phase of recurrent selection schemes? [J]. Mol Breed, 33(3): 555-567.
FUJIMAKI H, 1979. Recurrent selection by using genetic male sterility for rice improvement [J]. Jarq, 13(3): 153-156.
GRENIER C, TUONG VC, YOLIMA O, et al., 2015. Accuracy of genomic selection in a rice synthetic population developed for recurrent selection breeding? [J]. PLoS ONE, 11(5): 1-25.
JAIN HK, 2012. Transition to twenty-first century agriculture: Change of direction? [J]. Agric Res, 1(1): 12-17.
JIANG Y, CAI Z, XIE W, et al., 2012. Rice functional geno-mics research: Progress and implications for crop genetic improvement? [J]. Biotechnol Adv, 30(5): 1059-1070.
LI ZK, ZHANG F, 2013. Rice breeding in the post-genomics era: From concept to practice? [J]. Curr Opin Plant Biol, 16(2): 261-269.
MARAIS GF, BOTES WC, LOUW JH, 2010. Recurrent selection using male sterility and hydroponic tiller culture in pedigree breeding of wheat? [J]. Plant Breed, 119(5): 440-442.
MORAIS JOP, BRESEGHELLO F, BATISTA DJ, et al., 2017. Effectiveness of recurrent selection in irrigated rice breeding? [J]. Crop Sci, 57(6): 3043-3058.
MORAIS OP, MELO PGS, MORAIS OP, et al., 2017. Genetic variability during four cycles of recurrent selection in rice? [J]. Pesqui Agrop Bra, 52(11): 1033-1041.
MLLER D, SCHOPP P, MELCHINGER AE, 2017. Persistency of prediction accuracy and genetic gain in synthetic populations under recurrent genomic selection? [J]. Bethesda, 7(3): 801-811.
PANG YL, CHEN K, WANG XQ, et al., 2017. Recurrent selection breeding by dominant male sterility for multiple abiotic stresses tolerant rice cultivars? [J]. Euphytica, 12(1): 213-268.
PENG SB, KHUSH GS, VIRK P, et al., 2008. Progress in ideotype breeding to increase rice yield potential? [J]. Field Crop Res, 108(1): 1-38.
POSADAS LG, ESKRIDGE KM, SPECHT JE, et al., 2014. Elite performance for grain yield from unadapted exotic soybean germplasm in three cycles of a recurrent selection experiment [J]. Crop Sci, 54(6): 25-36.
RAMYA P, SINGH GP, JAIN N, et al., 2016. Effect of recurrent selection on drought tolerance and related morpho-physiological traits in bread wheat? [J]. PLoS ONE, 11(6): 1-17.
HU BL, WAN Y, LI X, et al., 2012. Analysis on genetic diversity of phenotypic traits in rice(Oryze sative) core collection and its comprehensive assessment? [J]. Acta Agr Sin, 38(5): 829-839.? [胡標林, 萬勇, 李霞, 等, 2012. 水稻核心種質表型性狀遺傳多樣性分析及綜合評價? [J]. 作物學報, 38(5): 829-839.]
HE ZZ, YIN M, XIE ZY, et al., 2014. Genetic diversity of tropical rice germplasms measured by phenotypic traits? [J]. Chin J Trop Agric, 34(9): 37-42.? [賀治洲, 尹明, 謝振宇, 等, 2014. 熱帶水稻優異種質資源的表型遺傳多樣性分析? [J]. 熱帶農業科學, 34(9): 37-42.]
LI ZC, ZHANG HL, ZENG YW, et al., 2001. Studies on phenotypic diversity of rice germplasm in Yunnan, China? [J]. Acta Agr Sin, 27(6): 832-837.? [李自超, 張洪亮, 曾亞文, 等, 2001. 云南稻種資源表型遺傳多樣性的研究? [J]. 作物學報, 27(6): 832-837.]
QI YW, 2004. Analysis of diversity and establishment of the core collection of improved rice varieties in China [D]. Beijing: China Agricultural University: 72-74. [齊永文, 2004. 中國水稻選育品種的遺傳多樣性分析及核心種質構建? [D]. 北京:中國農業大學: 72-74.]
QI YX, ZOU XY, 2008. Situation of recurrent selection applied in corp breeding? [J]. Acta Agric Jiangxi, 20(6): 14-17.? [漆映雪, 鄒小云, 2008. 輪回選擇在作物育種中的應用研究進展? [J]. 江西農業學報, 20(6): 14-17.]
WAN JM, 2007. Present status and prospect of molecular bree-ding in rice? [J]. Rev Chin Agric Sci Technol, 9(2): 1-9.? [萬建民, 2007. 中國水稻分子育種現狀與展望? [J]. 中國農業科技導報, 9(2): 1-9.]
WANG HG, JIA GQ, ZHI H, et al., 2016. Phenotypic diversity evaluations of foxtail millet core? [J]. Acta Agr Sin, 42(1): 19-30.? [王海崗, 賈冠清, 智慧, 等, 2016. 谷子核心種質表型遺傳多樣性分析及綜合評價? [J]. 作物學報, 42(1): 19-30.]
XIA JH, ZHENG YL, 2002. Molecular marker-assisted backcross breeding of maize Rf_3 NIL and its efficient analysis? [J]. Acta Agr Sin, 28(3):339-344.? [夏軍紅, 鄭用璉, 2002. 玉米Rf_3近等基因系的分子標記輔助回交選育與效益分析? [J]. 作物學報, 28(3): 339-334. ]
ZHU SS, ZHANG YS, 2004. Situation of recurrent selection applied in rice breeding? [J]. Guizhou Agric Sci, 32(2): 62-64.? [朱速松, 張玉珊, 2004. 輪回選擇在水稻育種中的應用研究進展 [J]. 貴州農業科學, 32(2): 62-64.]
(責任編輯 蔣巧媛)
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
