新型冠狀病毒肺炎的免疫失衡及干預策略
2020-04-26 07:04:22楊晨茜劉怡彤孟思偉王冰琳馮美卿
中國藥理學通報 2020年4期
楊晨茜,瞿 嬌,劉怡彤,孟思偉,王冰琳,馮美卿,孫 洋
(1. 南京大學生命科學學院生物技術與藥學系,醫藥生物技術國家重點實驗室,江蘇 南京 210023;2. 復旦大學藥學院,上海 201203)
1 引言
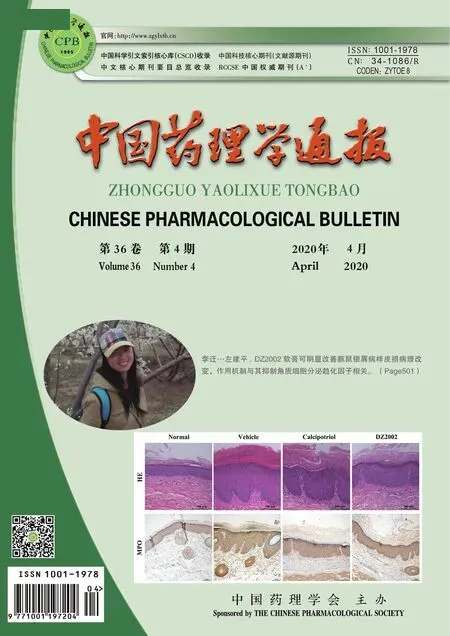
在過去的幾十年里,冠狀病毒(coronaviruses,CoVs)已經上升為全球公共衛生的重大威脅[1]。自2019年12月起,由新型冠狀病毒SARS-CoV-2引發的新型冠狀病毒肺炎(以下簡稱新冠肺炎)的迅速傳播加劇了人們對疫情的恐慌[2]。與先前爆發的SARS冠狀病毒(SARS-CoV)及中東呼吸綜合征冠狀病毒(MERS-CoV)相比,新型冠狀病毒除了具有相似的基因組、體內復制動力學和生物學性質外,還表現出一些獨特性[3-4]。因此,全面認識新型冠狀病毒對于疾病防控和藥物研發十分關鍵。
同時,臨床上觀測到新冠肺炎患者,尤其是重癥患者的淋巴細胞計數顯著降低,中性粒細胞異常增多,同時伴隨著細胞因子大量累積[5]。在SARS和MERS等病毒感染的后期,患者同樣出現免疫力低下和系統性炎癥[6]。這些現象提示新冠肺炎發病過程中存在免疫應答失調,通過對患者免疫狀態進行動態評估,有望為新冠肺炎患者診斷、治療效應監測及預后評估提供輔助信息。然而新冠肺炎免疫應答機制十分復雜,臨床用藥時機、方法及劑量亟待進一步探究。
該文通過對SARS冠狀病毒、MERS冠狀病毒以及新近流行的新型冠狀病毒進行全面比較,以更好地深入了解宿主-病原體相互作用,希望有助于從這幾次冠狀病毒引起的傳染病爆發中吸取教訓,積累經驗。通過對新冠肺炎免疫應答機制的深入研究,可為提高臨床療效及開發藥物奠定堅實的理論基礎。
2 冠狀病毒的介紹及分類
冠狀病毒(coronaviruses, CoVs)是屬于巢病毒目(nidovirales)的一類在自然界中廣泛存在的單鏈RNA病毒。冠狀病毒直徑約為80~120 nm,基因組全長約為27~32 kb,是目前已知的基因組最大的RNA病毒,可感染人類和多種哺乳動物,引起呼吸道、消化道以及神經系統等多種疾病,其中在人類中主要引起呼吸道的感染[3]。
冠狀病毒由刺突糖蛋白(spike protein,S蛋白)、核衣殼蛋白(nucleocapsid protein, N蛋白)、小包膜糖蛋白(envelope protein,E蛋白)、膜糖蛋白(membrane protein, M蛋白)以及核糖核酸組成[7]。S蛋白(180 ku)是一種插入到病毒包膜中的I型膜蛋白,賦予冠狀病毒特有的冠狀形態,可利用病毒和宿主細胞特異性受體的結合介導病毒的吸附和侵入過程。S蛋白由受體結合亞基S1和膜融合亞基S2組成。在病毒入侵時,S1亞基與宿主細胞表面受體結合使病毒附著于細胞表面,然后S2亞基促使病毒與宿主細胞的膜融合使病毒基因組進入細胞中[8]。N蛋白是核衣殼中唯一的蛋白質,也是病毒感染的細胞中含量最多的蛋白,它由兩個與RNA結合的N端結合域和C端結合域組成,負責與病毒基因組結合,識別基因組包裝信號將包膜基因組打包成病毒顆粒[9-11]。E蛋白(8~12 ku)是僅含有76~109個氨基酸殘基的完整跨膜蛋白,包括一個N端胞外域和一個C端胞內域,主要參與病毒粒子的組裝與釋放。M蛋白(25~30 ku)是冠狀病毒中含量最多的結構蛋白,含有3個跨膜結構域,它與E蛋白相互作用共同介導病毒包膜的組裝[12]。在一些β冠狀病毒中還存在血凝素糖蛋白(HE蛋白),該蛋白作為血凝素,與細胞表面的唾液酸結合,具有乙酰酯酶活性,促進S蛋白介導的病毒侵入過程和病毒通過粘膜的傳播[13]。
冠狀病毒具有高度保守的基因組結構,含有5′帽端結構和3′poly(A)尾,基因組中編碼非結構蛋白的復制酶基因約占基因組的三分之二,約20 kb;結構基因和輔助基因只占10 kb左右。冠狀病毒基因組順序如下:5′-復制酶(replicase)-S蛋白-E蛋白-M蛋白-N蛋白-3′。冠狀病毒整個感染過程包括吸附入侵、基因合成、病毒包裝和釋放4個步驟[14]。
根據遺傳學差異,冠狀病毒可分為α、β、γ、δ 四個屬,目前已發現七種人類冠狀病毒(HCoVs):HCoV-NL63、HCoV-229E、 HCoV-OC43、HCoV-HKU1、SARS-CoV、MERS-CoV以及2019新型冠狀病毒(SARS-CoV-2)[3]。
2.1 SARS冠狀病毒 (SARS-CoV)2002年11月,始于中國廣東省的以非典型性肺炎(非典)為主要臨床表現的嚴重急性呼吸系統綜合癥(SARS)短時間內迅速在世界范圍內傳播,共造成全球多個國家8 437例患者感染,超過900例患者死亡[15]。幾乎所有感染患者以發熱、干咳、呼吸短促等呼吸道癥狀開始,并在隨后幾天迅速發展為病毒性肺炎,胸部X射線檢查顯示肺部炎癥浸潤。部分患者出現淋巴細胞減少癥、白細胞減少癥、血小板減少癥[16-19]。通過對患者標本中分離出的病毒鑒定,發現造成此次疾病的病原體為一種冠狀病毒SARS-CoV[20-22],研究人員在中國馬蹄蝠中亦分離到了活性的SARS-CoV,證實蝙蝠是其自然界中的天然宿主[23-24]。SARS-CoV感染人類細胞的受體為一種金屬肽酶——血管緊張素轉換酶2(ACE2),它可有效地與病毒S蛋白的S1區域結合導致病毒的感染[25]。
2.2 中東呼吸綜合征冠狀病毒 (MERS-CoV)2012年9月,在沙特阿拉伯地區發現了一種來源于駱駝的高致病性的冠狀病毒,引起人類中東呼吸綜合征(MERS),這種冠狀病毒被命名為MERS-CoV[26-27]。據世界衛生組織的統計數據,MERS-CoV在全球范圍內造成2 066例患者感染,至少720例患者死亡,致死率遠高于SARS,致病性極強[28]。與SARS感染癥狀相似,MERS患者首先表現為發熱、寒顫和肌肉酸痛,隨后急性呼吸衰竭成為主要癥狀,部分患者伴有消化道癥狀和肝腎衰竭等并發癥[29]。所有患者在胸片上均有輕微到廣泛的單側和雙側異常。實驗室分析顯示,患者伴有乳酸脫氫酶和天冬氨酸氨基轉移酶濃度升高、血小板減少癥和淋巴細胞減少癥的表現[30]。不同于SARS-CoV和HCoV-NL63結合ACE2,MERS-CoV是以廣泛表達于肺泡上皮細胞的二肽基肽酶4(DDP4)作為感染人類細胞的表面受體[31]。這種受體分布的差異可能是引起SARS-CoV感染I型肺泡上皮細胞,而MERS-CoV感染II型肺泡上皮細胞和非纖毛支氣管細胞的主要原因[32-33]。
2.3 2019新型冠狀病毒 (SARS-CoV-2/2019-nCoV)2019年12月,中國湖北省武漢市爆發了一系列原因不明的與病毒性肺炎癥狀極為相似的肺炎病例。患者下呼吸道樣本深度測序發現這是一種新型冠狀病毒感染導致的肺炎,全基因組測序和系統發育分析表明這種新型冠狀病毒與SARS-CoV和MERS-CoV不同,被歸類為β冠狀病毒的2b譜系,世界衛生組織將其命名為2019-nCoV,而后國際病毒分類委員會將其命名為SARS-CoV-2[34]。截止目前全球47個國家共造成82 294例感染,2 804例死亡[35]。當前的證據表明,蝙蝠可能是該病毒的原始宿主,而中間宿主目前尚未有定論[36]。新型冠狀病毒感染導致臨床最為常見的癥狀為發熱、乏力和干咳,大多數患者出現呼吸困難和肺部雙側磨玻璃渾濁,常見并發癥有急性呼吸窘迫綜合征(ARDS)、急性心肌損傷、休克和繼發性感染,這些癥狀與SARS-CoV和MERS-CoV感染存在一定相似性,但新型冠狀病毒感染患者就診時發熱的比例明顯低于SARS-CoV和MERS-CoV感染,且較少的出現上呼吸道和消化道癥狀[5,37-38]。Tab 1從多個方面對3種冠狀病毒進行了比較,可以更加直觀地體現3種冠狀病毒的異同。
對新型冠狀病毒進行結構分析發現,其與SARS冠狀病毒的受體結合域具有高度相似性[36]。目前研究已證實新型冠狀病毒也利用ACE2受體作為病毒感染和入侵宿主細胞的特異性受體[39-40]。2020年2月16日美國德克薩斯大學奧斯汀分校的研究團隊利用冷凍電鏡技術,首次解析了新型冠狀病毒的S蛋白結構,他們發現新型冠狀病毒是利用高度糖基化的同源三聚體S蛋白進入宿主細胞的,且ACE2蛋白與新型冠狀病毒S蛋白的親和力是SARS冠狀病毒的10到20倍,這也在一定程度上解釋了新冠病毒具備更強傳染力的原因[41]。2月20日,西湖大學研究團隊利用冷凍電鏡在2.9埃全分辨率條件下全球范圍內首次成功解析ACE2的完整結構,揭示了人類全長的ACE2受體與新型冠狀病毒S蛋白受體結合結構域(RBD)復合體的精細化結構[42]。
Tab 1 Biological characteristics of SARS-CoV, MERS-CoV and SARS-CoV-2
在基因型-組織表達數據庫(genotype-tissue expression,GTEx)中,ACE2 mRNA可廣泛表達于全身各組織器官(Fig 1),其中在睪丸組織表達最高,其次是小腸末端回腸,而肺、腎臟、心臟等組織ACE2表達量相似,提示其均為新型冠狀病毒的易感器官。值得注意的是,ACE2組織表達分布提示睪丸表達量很高,提示病毒很可能作用于該器官,引起病毒性睪丸炎[43],因此治愈出院的新冠肺炎年輕男性患者,更需要定期隨訪。
3 新冠肺炎的免疫失衡及干預策略
雖然目前臨床上大部分新冠肺炎患者癥狀較輕,但仍存在部分患者早期病情較輕,后期突然加重的現象,最終死于多器官功能衰竭,而加重的原因可能是細胞因子風暴[5,37]。此外,新冠肺炎患者的中性粒細胞異常增多,同時伴隨著淋巴細胞計數的顯著降低[5,37-38]。這些現象提示新冠肺炎發病過程中存在免疫應答失調,下面將對這些現象進行逐一剖析,以進一步了解新冠肺炎發病的免疫應答機制,為制定相應的干預策略提供依據。
3.1 細胞因子風暴——新冠肺炎患者的“奪命殺手”細胞因子風暴(cytokine storm)一詞首次出現在1993年一篇關于移植物抗宿主疾病的文章中[44]。從2000年開始它被廣泛使用在多種感染性疾病如乙型溶血性鏈球菌感染、流感以及SARS中。近年來,細胞因子風暴也成為CART細胞療法帶來的最主要的副作用。那么什么是細胞因子風暴,細胞因子風暴,又稱炎癥風暴,是指機體在感染微生物或者其他劇烈刺激的情況下,免疫系統被過度激活,導致體液中多種細胞因子迅速大量產生的現象。細胞因子風暴引發的炎癥從局部開始通過系統循環擴散到全身,從而導致多種疾病如急性呼吸窘迫綜合征(ARDS)、敗血癥、急性胰腺炎等發生,嚴重威脅患者生命[45]。
在SARS冠狀病毒相關的細胞因子風暴中,重癥SARS患者血清中的促炎細胞因子(如IFN-γ, IL-1β, IL-6, IL-12, TGF-β)和趨化因子(如CXCL10,CXCL9, CCL2, IL-8)呈現高表達[46-49],但抗炎因子IL-10則低表達[46]。類似地,在MERS冠狀病毒引起的細胞因子風暴中,重癥患者血清中的促炎因子(如IFN-γ,TNF-α,IL-15,IL-17)和趨化因子(如CXCL10,CCL5,IL-8)高表達[50-52]。近來爆發的新冠肺炎重癥患者血漿中的IL-2, IL-7, IL-10, G-CSF, IP10, MCP-1, MIP-1A和TNF-α顯著升高[37]。值得注意的是,抗炎因子IL-10在SARS和新冠肺炎重癥患者中的表達趨勢完全相反。IL-10會抑制許多促炎細胞因子、趨化因子和趨化因子受體的表達,對細胞因子風暴起到抑制作用。新冠肺炎尤其是重癥患者外周血抗炎因子IL-10上升的原因有待進一步研究。
對于新冠肺炎尤其是重癥患者的治療,除了一些抗病毒藥物外,基于細胞因子風暴進行干預已成為減輕患者過度炎癥反應的有效策略(Fig 2)。IL-6是造成細胞因子風暴的典型促炎因子,IL-6阻斷策略已成功用于多種慢性炎癥疾病的治療。根據美國國家癌癥研究所提出的CTCAE v4.0制定了與抗體治療相關的細胞因子風暴的分級系統,在患者出現危及生命的癥狀,需要呼吸機支持或達到四級器官毒性時,可采用抗IL-6聯合糖皮質激素治療的手段[53]。第一個IL-6抑制劑托珠單抗(Tocilizumab)是一種人源化的抗IL-6受體單克隆抗體,目前廣泛用于類風濕關節炎的治療[54]。通過分析123例新冠肺炎患者外周血和細胞因子分析狀況,臨床研究人員發現重癥患者中CD4 T和CD8 T細胞比例明顯降低,而促炎因子IL-6水平明顯升高,表明T細胞亞群和細胞因子可作為預測病情由輕到重轉變的依據之一[55]。目前以中國科學技術大學附屬第一醫院牽頭的托珠單抗在新型冠狀病毒肺炎(COVID-19)中的有效性及安全性的多中心、隨機對照臨床研究(注冊號:ChiCTR2000029765)正在開展中。TNF-α是急性病毒性疾病中十分關鍵的細胞因子,可通過過度的炎癥反應加重冠狀病毒的致病性。已有研究報道,使用TNF-α中和抗體或可溶性TNF-α受體融合蛋白治療可減少流感病毒感染的小鼠肺部細胞因子產生和炎性細胞浸潤[56-57]。在小鼠模型中敲除TNF-α受體可明顯緩解SARS冠狀病毒感染所導致的嚴重組織損傷[58]。臨床報道新冠肺炎重癥患者血漿中TNF-α水平顯著上升[37],提示在重癥患者上使用TNF-α中和抗體亦是阻斷細胞因子風暴的有效手段。阿達木單抗為抗人腫瘤壞死因子(TNF)的人源化單克隆抗體。事實上,上海長征醫院已在中國臨床試驗注冊中心申請格樂立(阿達木單抗)治療新型冠狀病毒肺炎(COVID-19)重型和危重型患者的安全性和有效性的臨床研究(注冊號:ChiCTR2000030089 )。
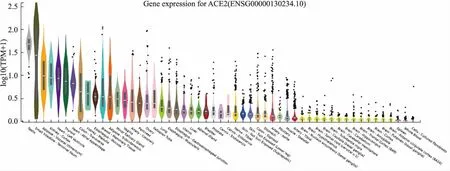
Fig 1 ACE2 mRNA expression distribution in human tissues based on GTEx database
Fig 2 Cytokine storm, also termed cytokine-release syndrome, in SARS-CoV-2-induced acute respiratory distress syndrome (ARDS)
另一方面,鐘南山院士團隊分析了1 099新冠肺炎病例發現,只有43.1%的患者在入院時出現發熱癥狀,即使是重癥患者發熱比例也只有47.4%,而在這些重癥發熱患者中,入院時體溫大于38.0 ℃的高熱只占26.3%[59]。對比1 425 例SARS患者的臨床數據發現,其發熱比率高達94%[60],可見新冠肺炎患者就診時發熱的比例明顯低于SARS患者。南京市第二醫院聯合南京大學的研究團隊最近發現,相比于22例非新冠病毒感染的肺炎患者,71例普通型新冠肺炎患者T細胞和B細胞更加顯著地下降,而單核細胞、中性粒細胞和NK細胞數量卻無顯著差異[38],而這3種細胞恰恰是細胞因子風暴的主要參與者。華中科技大學同濟醫學院的研究團隊通過對外周血細胞因子動態檢測發現,與普通型新冠肺炎患者(27例)相比,重癥患者(13例)的多種細胞因子如IL-6、IL-2、TNF-α、IFN-γ以及IL-10僅在某一時間段有顯著性上升,而其他時間段兩組無明顯差異,且這些炎癥因子的血清水平相對較低[61],從這13例新冠肺炎重癥患者來看,細胞因子風暴的現象并不明顯,可能與目前的樣本數偏少有關。以上結果提示,對于新冠肺炎患者細胞因子風暴是否為病情進展的主要因素還需要進一步確證。但不可否認,失控的細胞因子風暴會進一步造成炎癥彌漫,導致肺泡結構受損,進而機體肺血氧不足,最終進展為急性呼吸窘迫綜合征(ARDS),危及患者生命。
3.2 中性粒細胞捕獲網——新冠肺炎的免疫干預新靶點中性粒細胞是外周血白細胞中最豐富的細胞類型,是機體先天免疫的重要組成部分,在介導炎癥反應和免疫應答方面發揮著十分重要的作用[62]。一方面,中性粒細胞可經過沿血管內壁運動、結合內皮細胞選擇素/黏附素、遷移穿過內皮-上皮細胞層等步驟在病毒感染24小時內進入肺泡腔中[63]。在肺泡腔內,中性粒細胞被腔內炎癥因子和病原體迅速激活,通過吞噬作用清除病原微生物[64]。另一方面,中性粒細胞可在炎癥環境下,釋放一種以DNA為骨架內含多種殺菌蛋白的網狀結構,即中性粒細胞捕獲網(neutrophil extracellular traps, NETs),可有效清除病原微生物,抑制其擴散[65]。
然而,越來越多的研究表明:中性粒細胞及其胞外捕獲網既能殺滅入侵的病原微生物,又能加重組織器官的損傷。這主要歸因于在清除病毒的過程中,中性粒細胞產生的活性氧、細胞因子、NETs均對上皮-內皮屏障具有破壞作用[66]。有研究表明NETs參與了多種病毒包括流感病毒在內感染導致的肺部炎癥進程[67-72]。重癥流感患者的血漿中NETs水平明顯升高,同時中性粒細胞對IL-8或脂多糖刺激會釋放更多的MPO-DNA復合物;另外,H7N9和H1N1感染患者體內NETs可以增加肺泡上皮細胞的通透性[67]。在甲型H1N1流感病毒PR8株侵染的小鼠模型中也觀察到過多NETs的形成,進而損傷肺泡毛細血管,引發肺功能障礙[68]。補體C5a介導的NETs會加劇急性肺損傷病人對高致病性病毒的感染。在感染H1N1的小鼠模型中發現阻斷C5a會抑制中性粒細胞和巨噬細胞在氣道中的浸潤,減少NETs的形成以及肺上皮的損傷[69]。臨床研究表明[70],急性呼吸窘迫綜合征患者支氣管肺泡灌洗液內中性粒細胞的濃度和NETs的數量與疾病嚴重程度呈正相關。值得一提的是,中性粒細胞暴露于急性呼吸窘迫綜合征患者支氣管肺泡灌洗液后可加速NETs的產生[71],這也許歸因于肺泡灌洗液中細胞因子風暴的作用。MERS患者和近來爆發的新冠肺炎患者均表現出中性粒細胞大量增多的現象[61,72]。全國首例新冠肺炎逝者的遺體解剖報告顯示,新冠肺炎主要引起深部氣道和肺泡損傷為特征的炎性反應,肺部纖維化及實變沒有SARS導致的病變嚴重,但滲出性反應較SARS明顯。尸體肺部切面可見大量粘稠的分泌物從肺泡內溢出[73],這或許與NETs在肺部異常聚集有關(Fig 3),但分泌物形成的具體原因仍需研究。
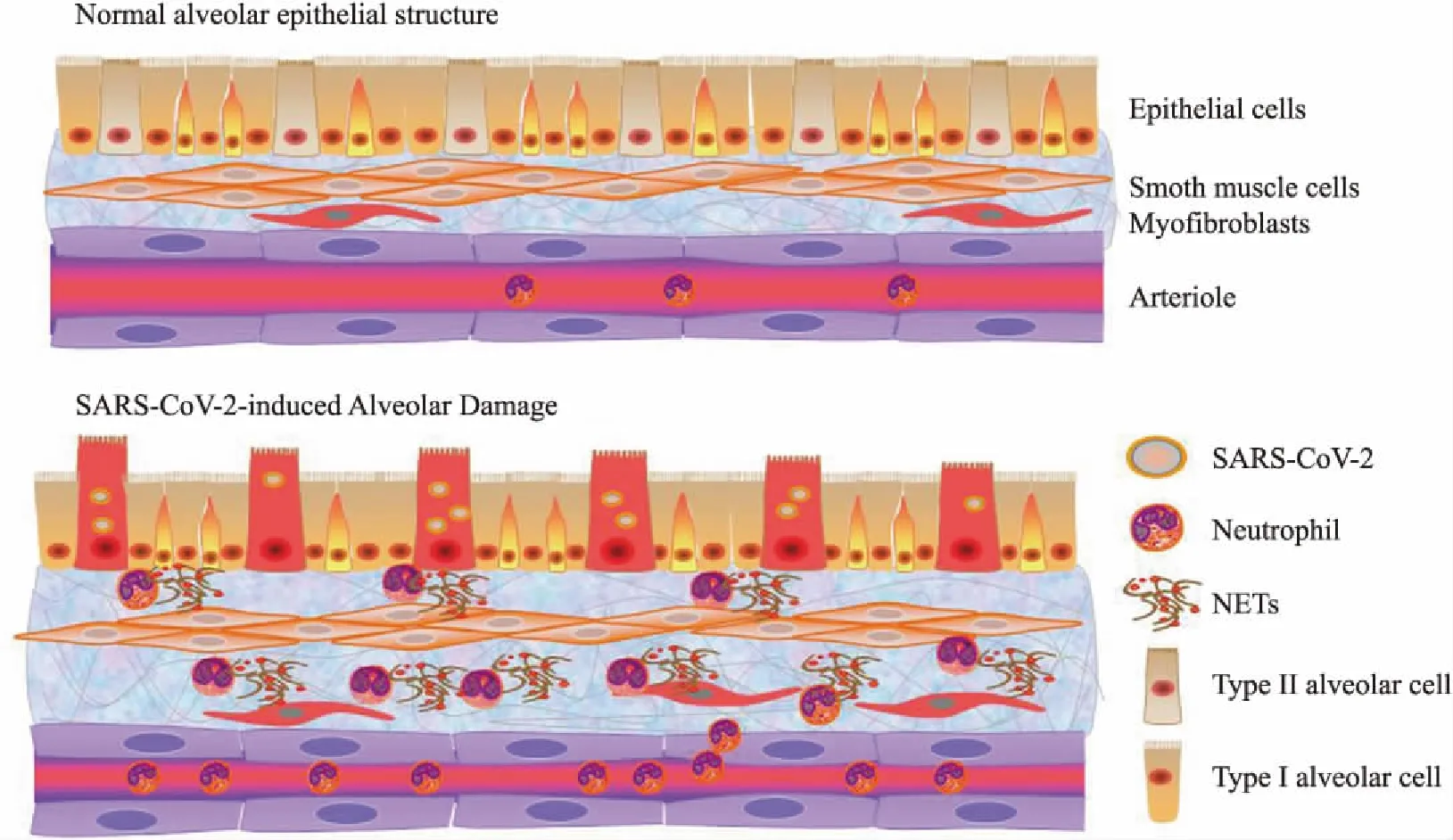
Fig 3 Neutrophil extracellular traps (NETs) is possibly invovled in SARS-CoV-2-induced lung injury
目前,進入臨床試驗階段的藥物也有與NETs潛在的關聯。例如,利用抗IL-6受體的單克隆抗體托珠單抗(Tocilizumab)治療新冠肺炎的臨床試驗(注冊號:ChiCTR2000029765)已在開展之中,而IL-6是誘導NETs的重要炎癥因子。此外,新冠肺炎的治療方案中推薦了氯喹[74],而已有研究顯示自噬抑制劑氯喹能抑制NETs的形成,提高胰腺炎小鼠的生存率[75]。相信隨著研究的深入,針對NETs為靶點的治療策略將為臨床病毒性肺炎提供新的選擇。
3.3 淋巴細胞亞群失衡——新冠肺炎臨床現象帶來的思考淋巴細胞(lymphocyte)可分為T淋巴細胞(T細胞)、B淋巴細胞(B細胞)和自然殺傷(NK)細胞,是免疫系統功能調節的主要執行者。在免疫應答過程中,當各淋巴細胞亞群的數量及功能出現異常時,機體可發生一系列的病理改變。因此,淋巴細胞亞群失衡是反映機體免疫應答異常的重要指標[76]。
南京市第二醫院聯合南京大學的團隊近期研究顯示,相比于非新冠病毒引起的肺炎(non-COVID-19),新冠肺炎(COVID-19)患者出現明顯的T細胞和B細胞計數減少,且數量與器官損傷相關生化指標呈負相關[38]。此外,鐘南山院士團隊對1 099例新冠肺炎患者臨床特征的回顧性研究顯示,82.1%的患者出現淋巴細胞減少癥狀,其中重癥患者占95.5%(147/154),顯著高于非重癥患者[59]。北京地壇醫院研究團隊報道,中性粒細胞與淋巴細胞比值(neutrophil-to-Lymphocyte Ratio, NLR)可作為新冠肺炎患者發展為重癥的獨立危險因素[72]。深圳市第三人民醫院的研究團隊,通過收集新冠肺炎患者支氣管肺泡灌洗液中的免疫細胞,利用單細胞TCR免疫組庫測序技術,發現與輕癥患者相比,重癥患者體內可殺傷新冠病毒的細胞毒性T淋巴細胞的克隆數目顯著下降[77]。這一系列的數據提示新冠肺炎患者出現明顯的淋巴細胞亞群失衡,從而造成免疫功能受損,這種情況在重癥患者上更為嚴重[37]。此外,陸軍軍醫大學的研究團隊發現新冠肺炎患者T細胞計數明顯減少以及細胞因子的累積,而存活的T細胞也呈現功能耗竭狀態。隨著患者病情進展,T細胞上PD-1和Tim-3表達增加,提示T細胞逐漸失能[78]。無獨有偶,四川大學華西醫院已經在中國臨床試驗注冊中心申請PD-1單抗用于治療新型冠狀病毒肺炎(COVID-19)重型或危重型患者的臨床對照研究(注冊號:ChiCTR2000030028),利用PD-1單抗阻斷PD-1/PD-L1通路,恢復T細胞功能改善免疫失衡。此外,減少的T細胞中,CD4及CD8 T細胞下降幅度更為顯著,且與病情嚴重程度正相關[55,61,79]。另一方面,解放軍總醫院王福生院士團隊對一例新冠肺炎死亡患者進行病理檢測[80],外周血的流式分析發現CD4及CD8 T細胞數量雖然大幅降低,但T細胞的狀態卻被顯著激活。CD4 T細胞中具有高促炎效應的Th17細胞數量增加,CD8 T細胞則呈現高顆粒酶細胞毒性,這一現象似乎可以解釋患者出現的嚴重免疫損傷。Th17細胞作為T細胞獨特的一類分支,其表達失調使患者病情更加錯綜復雜,同時提示淋巴細胞在新冠肺炎中的重要作用需要進一步探究。總而言之,臨床治療中密切關注T細胞,尤其是CD4及CD8 T細胞數量及功能的變化,或將直接影響后期治療策略的選擇。
針對新冠肺炎患者出現淋巴細胞數量減少的現象,臨床上會使用諸如胸腺肽、丙種球蛋白等改善機體低下的免疫功能;而由細胞因子引發的免疫系統正反饋而造成的細胞因子風暴,臨床上最常用的藥物為糖皮質激素,然而糖皮質激素在抑制細胞因子風暴的同時會引發免疫抑制并延緩冠狀病毒的清除,有繼發性感染等風險[81]。最近來自Lancet的述評探討了糖皮質激素治療新冠肺炎的可行性,文章指出將糖皮質激素用于新冠肺炎的治療需要符合相應的臨床指征[82]。糖皮質激素使用的時機和劑量對于合理有效的免疫調節至關重要,需要更多有價值的研究為疫情診療提供證據,并推動相關指南的制定。
綜上可知,對于新冠肺炎患者尤其是重癥患者,病情的轉折可能與淋巴細胞亞群失衡造成的免疫功能紊亂有關。通過結合患者基本臨床特征與淋巴細胞的數量、功能以及細胞因子的種類、水平,有望挖掘出新冠肺炎診斷及預后的潛在標志物。
4 總結與展望
新冠肺炎發病是病毒與機體免疫系統“互相博弈”的過程,面對病毒的感染,機體需要啟動免疫系統及時識別、清除病毒,但是失調的、過度的免疫反應又會引起患者體內炎癥因子過度表達而出現細胞因子風暴。此外,臨床上觀察到新冠肺炎患者往往出現中性粒細胞的大量累積以及淋巴細胞數量的下降,這些都有望成為新冠肺炎免疫干預的切入環節。目前,多種已上市藥物或臨床前階段的候選藥物正在積極開展臨床試驗用于新冠肺炎的防治[83-84]。這其中的一些藥物發揮療效離不開對免疫系統的調節。未來研究人員將進一步深入探索新冠肺炎發病過程中的免疫應答機制,闡明相關的靶點及信號通路,以推動治療藥物和疫苗的研發,為臨床上新冠肺炎的診療及預后提供可靠的科學依據。