17種錦雞兒屬植物葉片解剖結構及抗旱性分析
2020-04-27 08:06:22馬紅英呂小旭計雅男李小偉
水土保持研究 2020年1期
馬紅英, 呂小旭, 計雅男, 李小偉
(1.寧夏大學 農學院, 銀川 750021; 2.甘肅醫學院, 甘肅 平涼 744000)
錦雞兒屬(CaraganaFabr.)是豆科蝶形花亞科灌木或小喬木。全世界約有100種,主要分布于亞洲和歐洲東部溫帶干旱和半干旱地區,約有66種分布于我國東北、華北、西北、西南各省區,是溫帶荒漠、半荒漠地區及亞熱帶高寒地區的主要建群種[1],因此我國錦雞兒屬植物資源豐富。該屬中許多植物在我國被用于防風固沙、水土保持、牲畜飼料以及生產燃料和纖維的生物資源,其中10余種被用在傳統中藥、蒙藥和藏藥中已有悠久的歷史,被認為具有“滋陰,健脾,活血和促血”的功效[2];錦雞兒屬植物根系發達,具根瘤,抗旱耐瘠,是水土保持和防風固沙的優良樹種[3];在黃土高原地區進行了大面積的推廣應用,已證實在退化嚴重的半干旱和干旱地區,該屬植物對植被恢復具有重要的意義[4]。鑒于此,近年來,錦雞兒屬植物備受學者關注,國內外學者主要對錦雞兒屬植物的系統分類與區系[5]、形態解剖[6-7]、遺傳與育種[8-9]和細胞與分子生物學[10]等方面進行了報道。
干旱是影響植物正常生長的主要因素之一[11-12],植物對干旱環境的適應是形態結構、生理和生化等多方面綜合作用的結果,且形態結構是植物生理生化反應的基礎。而植物葉片直接暴露在外界環境中,對外界環境反應最為敏感,其結構特征能夠反映植物對水分的利用狀況,因此植物葉片、上下表皮、柵欄組織、海綿組織和主脈厚度能夠反映植物的抗旱特征。有研究表明:葉片厚度、柵欄組織厚度、主脈厚度和緊密度等與抗旱性呈正相關關系;海綿組織厚度、疏松度等與抗旱性呈負相關關系[13-14]。
本研究選取民勤沙生植物園和賀蘭山南寺17種錦雞兒屬植物葉片為試驗材料,借助石蠟切片技術觀察葉片解剖結構,綜合評價其抗旱特性,其研究結果對干旱、半干旱地區植被恢復樹種選擇具有指導意義,同時為錦雞兒屬植物資源的合理開發利用提供理論依據。
1 材料與方法
1.1 材料區概況
本研究所用材料取自甘肅民勤沙生植物園和內蒙古賀蘭山南寺。民勤沙生植物園位于民勤縣騰格里沙漠西緣,氣候屬溫帶大陸性極干旱氣候,具有明顯的沙漠氣候特征,風大沙多,降雨稀少,氣候干燥,日照強烈,年平均氣溫7.4℃,年平均降水量110 mm,年蒸發量2 500~2 700 mm,無霜期180 d左右;賀蘭山南寺位于巴彥浩特東南30 km處,氣候屬中溫帶干旱地區,該地區干旱少雨,風大沙多,日照強烈,蒸發量大,年平均降雨量200 mm,年蒸發量1 500~3 000 mm,無霜期145 d左右。
1.2 研究材料
采集的材料包括南口錦雞兒(C.zahlbruckneri)、狹葉錦雞兒(C.stenophylla)、甘蒙錦雞兒(C.opulens)、短腳錦雞兒(C.brachypoda)、荒漠錦雞兒(C.roborovskyii)、邊塞錦雞兒(C.bongardiana)、樹錦雞兒(C.sibirica)、紅花錦雞兒(C.rosea)、黃刺條錦雞兒(C.frutex)、多刺錦雞兒(C.spinosa)、刺葉錦雞兒(C.acanthophylla)、灰毛小葉錦雞兒(C.microphylla)、檸條錦雞兒(C.korshinskii)、粉刺錦雞兒(C.pruinosa)、中間錦雞兒(C.intermedia)、鬼箭錦雞兒(C.jubata)和川西錦雞兒(C.erinacea)共17種(表1)。每種植物選取發育良好、有代表性的植株,選取其葉,每種錦雞兒采6株,共6次重復。于2018年6月采樣,經FAA固定,采用石蠟切片法切片(取材、固定、脫水、透明、透蠟、包埋、切片、貼片、染色、封藏等步驟)制成永久制片,將制片置于Olympus顯微鏡下觀察、并用Motic Image Advanced 3.0軟件測量并進行顯微照相,2018年10月完成測定。
1.3 數據處理
選擇葉片厚度、上表皮細胞厚度、下表皮細胞厚度、柵欄組織厚度、海綿組織厚度、主脈厚度、葉片結構緊密度和葉片結構疏松度共8項葉片抗旱性結構指標進行測定。柵/海=柵欄組織厚度/海綿組織厚度,組織結構緊密度=柵欄組織厚度/葉片厚度,組織結構疏松度=海綿組織厚度/葉片厚度。Microsoft Excel 2010軟件整理試驗數據,并使用SPSS 21.0軟件對葉片樣本進行層次聚類分析。
1.4 抗旱能力綜合評價
通過模糊數學隸屬函數值法綜合評價17種錦雞兒屬植物的抗旱性[15]。
R(Xi)=(Xi-Ximin)/(Ximax-Ximin)
(i=1,2,3,…,n)
(1)
式中:Xi為指標測定值;Ximax和Ximin分別為所有材料中同一指標的最大值和最小值。若測定指標與抗旱性為負相關關系,則計算方法為:
R(Xi)=1-(Xi-Ximin)/(Ximax-Ximin)
(i=1,2,3,…,n)
(2)
將抗旱隸屬函數值進行累加,求其平均數:
X=∑R(Xi)/n(i=1,2,3,…,n)
式中:X為平均抗旱隸屬值。X大表示抗旱性強;X小表示抗旱性弱。
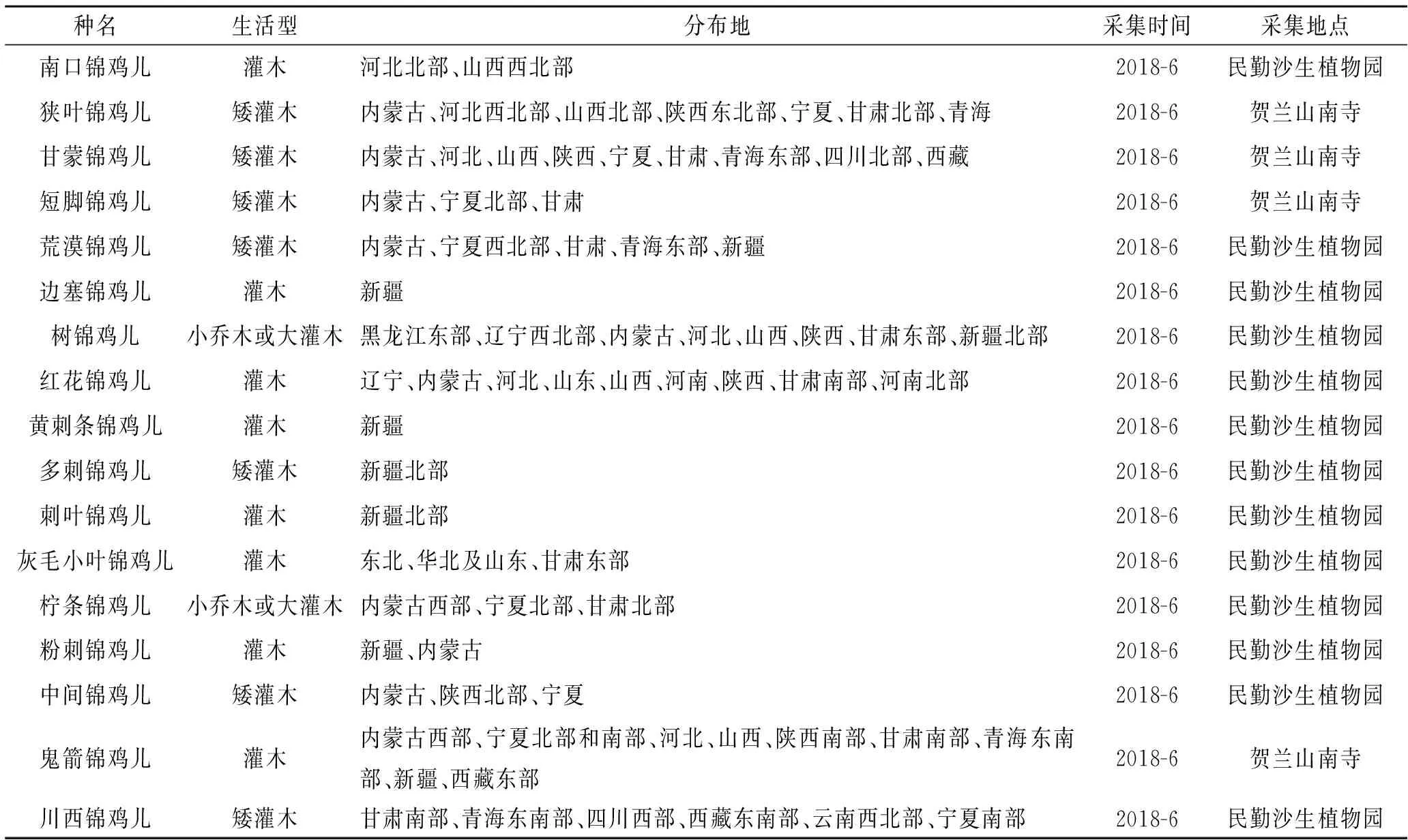
表1 材料名稱及來源
2 結果與分析
2.1 17種錦雞兒屬植物葉片解剖結構特征
通過對17種錦雞兒屬植物葉片橫切面結構(圖1)觀察,葉表皮細胞均為單層;但葉肉結構組成有顯著差異,其解剖結構特點是:葉肉組織類型出現普通型、過渡型和環柵型,其中普通型柵欄組織和海綿組織并存,過渡型柵欄組織趨于發達,海綿組織逐漸退化,環柵型海綿組織基本消失。因此可分為以下3種類型:(1) 普通型,即葉肉組織在近軸面分化為柵欄組織,在遠軸面分化為海綿組織,分別為南口錦雞兒、甘蒙錦雞兒、樹錦雞兒、紅花錦雞兒、黃刺條錦雞兒、粉刺錦雞兒和中間錦雞兒;(2) 環柵型,即葉肉組織在近軸面和遠軸面都分化為柵欄組織,這種類型是狹葉錦雞兒、短腳錦雞兒、荒漠錦雞兒、多刺錦雞兒和灰毛小葉錦雞兒;(3) 過渡型,其海綿組織細胞形狀趨于柵欄細胞狀,且有些部位呈柵欄狀,分別是邊塞錦雞兒、刺葉錦雞兒、檸條錦雞兒、鬼箭錦雞兒和川西錦雞兒。
與抗旱有關的葉肉結構特征數據(表3)顯示,多刺錦雞兒葉片厚度最大,為302.91 μm,樹錦雞兒的葉片厚度、柵欄組織厚度、海綿組織厚度和主脈厚度最小,分別為101.44 μm,31.18 μm,23.82 μm,53.96 μm。短腳錦雞兒上表皮細胞厚度最大,為21.02 μm。川西錦雞兒上表皮細胞厚度和下表皮細胞厚度最小,但組織結構緊密度最大,分別為10.34 μm,8.632 μm和0.50。狹葉錦雞兒下表皮細胞厚度和主脈厚度最大,但組織結構緊密度最小,分別為19.06 μm,125.16 μm和0.27。邊塞錦雞兒柵欄厚度和海綿厚度最大,分別為126.46 μm和67.88 μm。中間錦雞兒組織結構疏松度最大,為0.31;粉刺錦雞兒組織結構疏松度最小,為0.18。
2.2 解剖結構的聚類分析
聚類分析結果表明(圖2):17種錦雞兒屬植物分為2類;第1類包括荒漠、短腳、邊塞和多刺錦雞兒4種,葉片厚度、柵欄組織厚度、海綿組織厚度、葉片結構緊密度大,表皮細胞層厚度較第2類偏小(除短腳錦雞兒),葉片結構疏松度小;第2類包括灰毛小葉、鬼箭、中間、紅花、黃刺條、甘蒙、川西、檸條、粉刺、南口、刺葉、樹和狹葉錦雞兒13種,葉片結構疏松度大,葉片厚度、柵欄組織厚度、葉片結構緊密度小,表皮細胞厚度小。
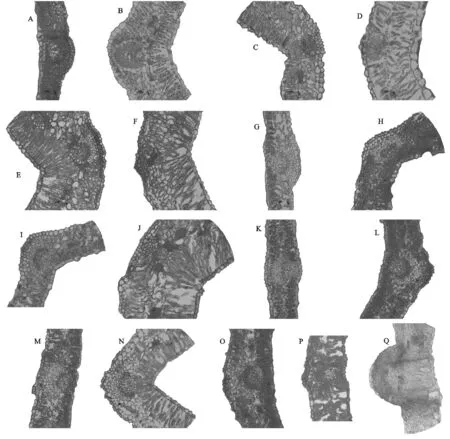
注:A南口錦雞兒,B狹葉錦雞兒,C甘蒙錦雞兒,D短腳錦雞兒,E荒漠錦雞兒,F邊塞錦雞兒,G樹錦雞兒,H紅花錦雞兒,I黃刺條錦雞兒,J多刺錦雞兒,K刺葉錦雞兒,L灰毛小葉錦雞兒,M檸條錦雞兒,N粉刺錦雞兒,O中間錦雞兒,P鬼箭錦雞兒,Q川西錦雞兒。
2.3 基于隸屬函數對錦雞兒屬植物抗旱性分析
不同錦雞兒屬植物具有不同的抗旱性,本研究選取了8項葉片抗旱性結構指標,利用隸屬函數來評價錦雞兒屬植物的抗旱能力,其數值越大表明抗旱性越強。結果顯示,17種錦雞兒屬植物抗旱能力由大到小排序為:短腳錦雞兒>荒漠錦雞兒>多刺錦雞兒>狹葉錦雞兒>川西錦雞兒>粉刺錦雞兒>邊塞錦雞兒>南口錦雞兒>紅花錦雞兒>甘蒙錦雞兒>鬼箭錦雞兒>檸條錦雞兒>黃刺條錦雞兒>刺葉錦雞兒>灰毛小葉錦雞兒>中間錦雞兒>樹錦雞兒(表4)。
3 討 論
在整個生長發育過程中,環境對植物的形態建成起著重要作用,形態特征是植物適應環境的直接表現形式[16],干旱是影響植物生長發育的重要因素之一,不同種植物受自身遺傳特性與外界環境的共同影響,對干旱形成了不同的生態適應對策。植物葉片與外界進行水分和氣體交換,承擔著光合和蒸騰作用,是植物對逆境脅迫最敏感的部位,在復雜的外界環境中形成了特殊的結構與功能[17],可較好地反映植物對干旱等逆境條件的適應性。根據葉的生長狀況和形態結構等特征可判斷植物對環境的適應能力[18],因此植物葉片常被植物生理學家用來作為主要研究對象[19]。
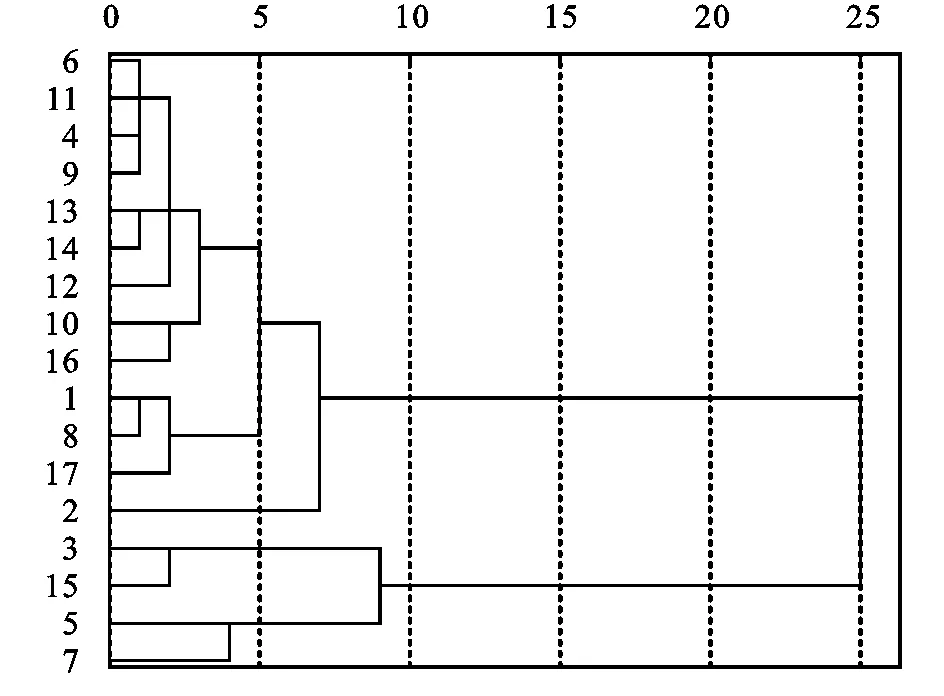
注:圖中數字表示:1南口錦雞兒,2狹葉錦雞兒,3荒漠錦雞兒,4中間錦雞兒,5邊塞錦雞兒,6灰毛小葉錦雞兒,7多刺錦雞兒,8刺葉錦雞兒,9紅花錦雞兒,10檸條錦雞兒,11鬼箭錦雞兒,12川西錦雞兒,13黃刺條錦雞兒,14甘蒙錦雞兒,15短腳錦雞兒,16粉刺錦雞兒,17樹錦雞兒。
環境對葉片結構的影響主要體現在葉片厚度、上表皮細胞厚度、下表皮細胞厚度、柵欄組織厚度、海綿組織厚度、主脈厚度、葉片結構緊密度、葉片結構疏松度等方面。其中葉片厚度是植物的抗旱特征之一,吳林等[20]認為植物葉片越厚,保水能力越強,植株則越抗旱;徐揚等[21]認為葉片柵欄組織越發達,植株的抗旱性越強;任媛媛等[22]和劉紅茹等[13]認為主脈中發達的維管束和強化的機械組織可以使植株更加抗旱。本研究結果發現:17種錦雞兒屬植物抗旱機制各異,其中短腳錦雞兒葉肉組織較疏松,但其表皮比較厚,葉片的儲水、保水及抗旱能力則越強[23];多刺錦雞兒葉片厚度大且柵欄組織厚度也較大,抗旱性也較強,但表皮細胞厚度卻小于短腳錦雞兒,可見表皮細胞越厚可以更大程度的限制蒸騰的發生,增強抗旱能力;狹葉錦雞兒主脈較發達、表皮厚度較大,以此來提高植物的抗旱性;川西錦雞兒與粉刺錦雞兒通過增加柵海比提高植物的抗旱性,但川西錦雞兒發達的柵欄組織使它的組織結構緊密度增加,因此它的抗旱性更強;邊塞錦雞兒通過增加柵欄組織厚度提高植物的抗旱性。在葉肉結構從普通型過渡到環柵型的過程中,葉片厚度、柵欄組織、海綿組織等發生一系列的變化,使得不同錦雞兒對干旱有不同的應對策略,這也是錦雞兒屬植物從東向西抗旱性增強,出現替代分布的重要原因之一[24]。
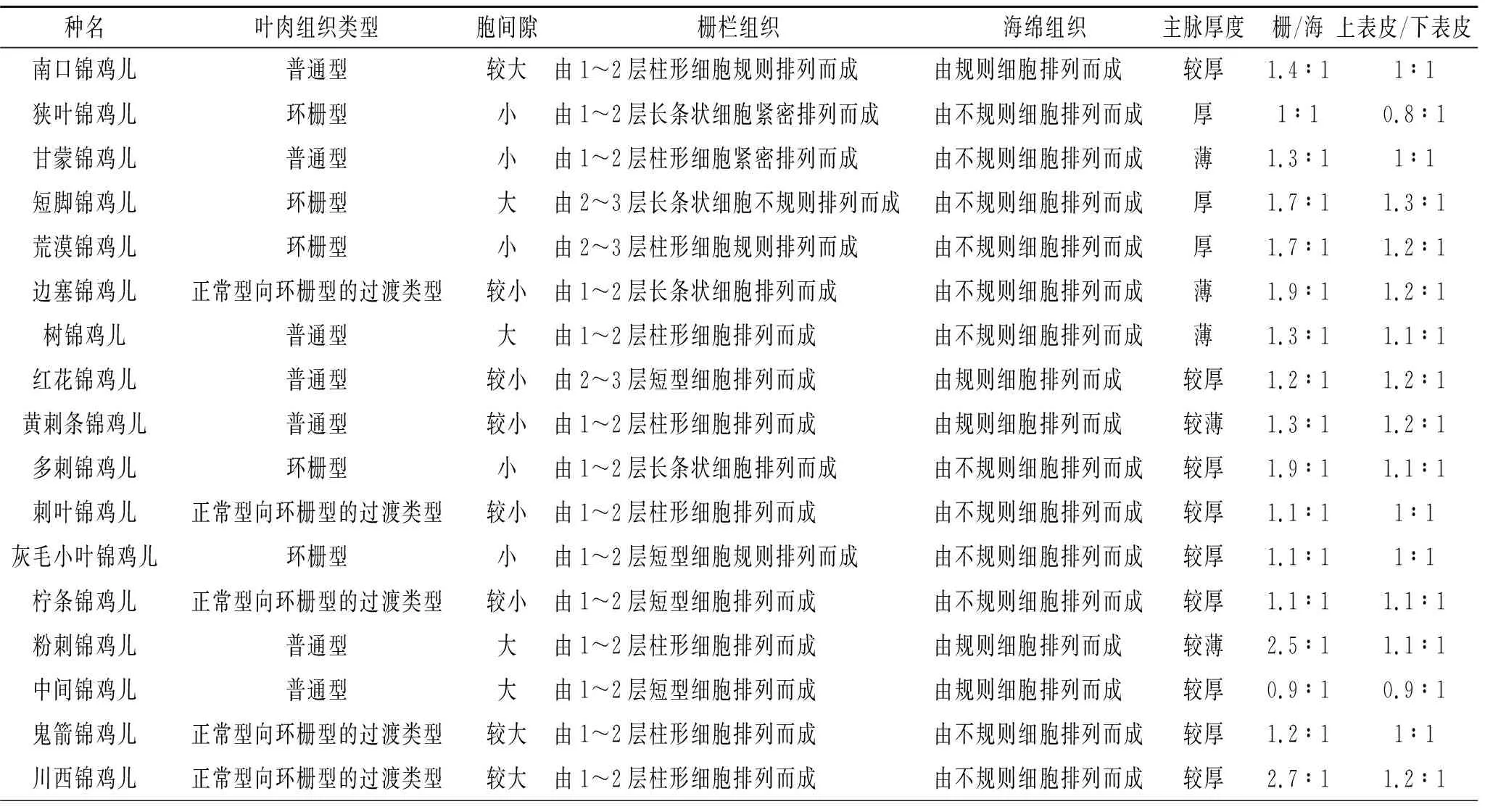
表2 17種錦雞兒屬植物葉片解剖結構及主要特征
通過對17種錦雞兒屬植物進行聚類分析,可將不同種錦雞兒屬植物分為2類。層次聚類分析的第1類錦雞兒中短腳錦雞兒葉片厚度較小,但其依靠表皮細胞厚度以及柵欄組織厚度防止水分過分蒸發,使其適應干旱的環境[25]。多刺錦雞兒表皮細胞厚度較小,葉片厚度、柵欄組織厚度以及組織結構緊密度大,這些特征可以增強光合能力,提高光合效率,達到高效利用水分的能力[26]。邊塞錦雞兒雖主脈厚度較小,但其具有發達的柵欄組織,使得抗旱性增強。第2類錦雞兒中狹葉錦雞兒雖然柵欄組織厚度較小,但其表皮細胞厚度大、主脈發達,主脈密度越高,抑制水分散失的能力越強,能夠忍耐的最低水勢也越低,則其抗旱能力越強[27]。川西錦雞兒葉片厚度以及表皮細胞厚度較小,其柵欄組織厚度以及組織結構緊密度較大,可提高水分利用效率,減少水分散失,使其抗旱性增強。
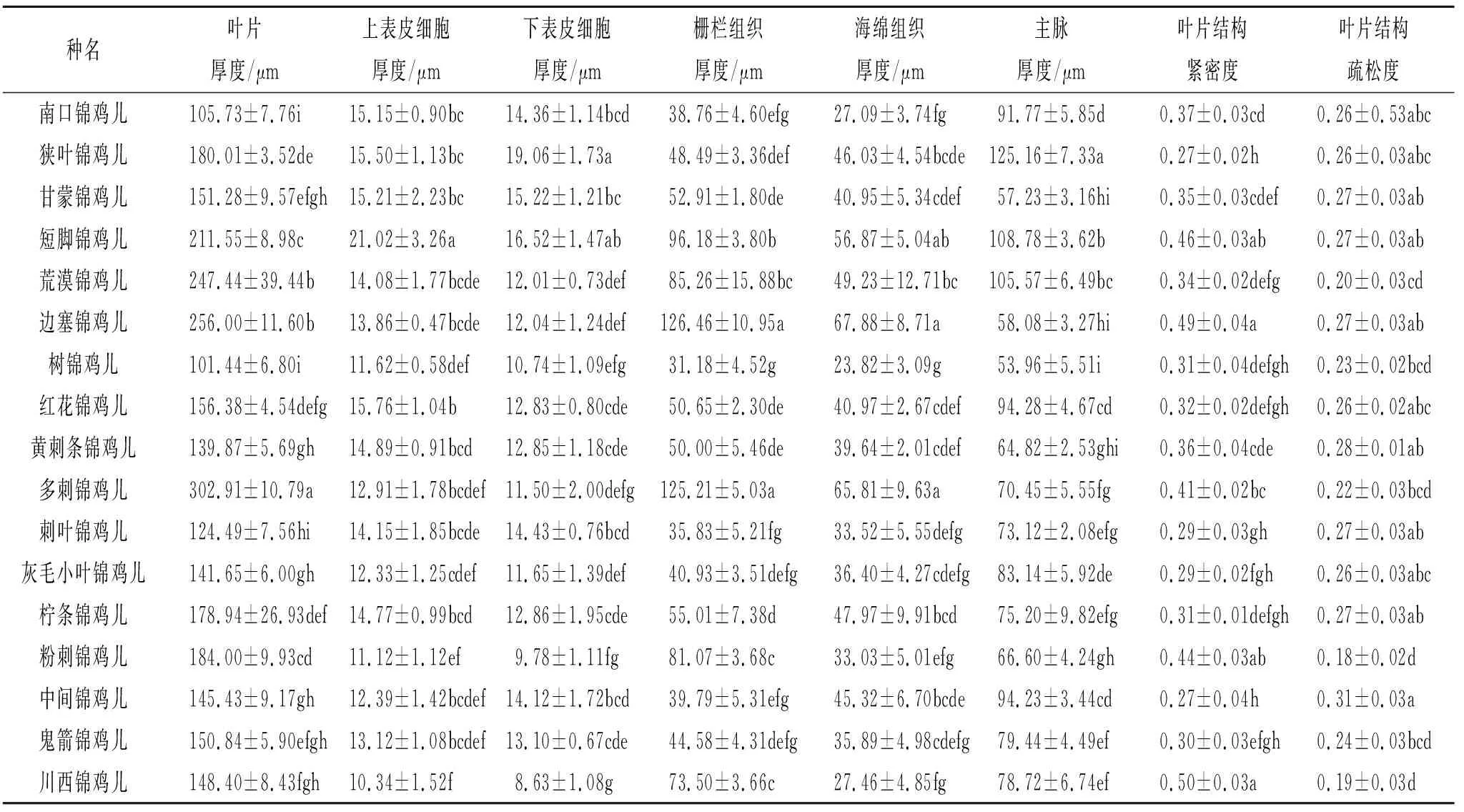
表3 17種錦雞兒屬植物葉片解剖結構特征數據(n=6)
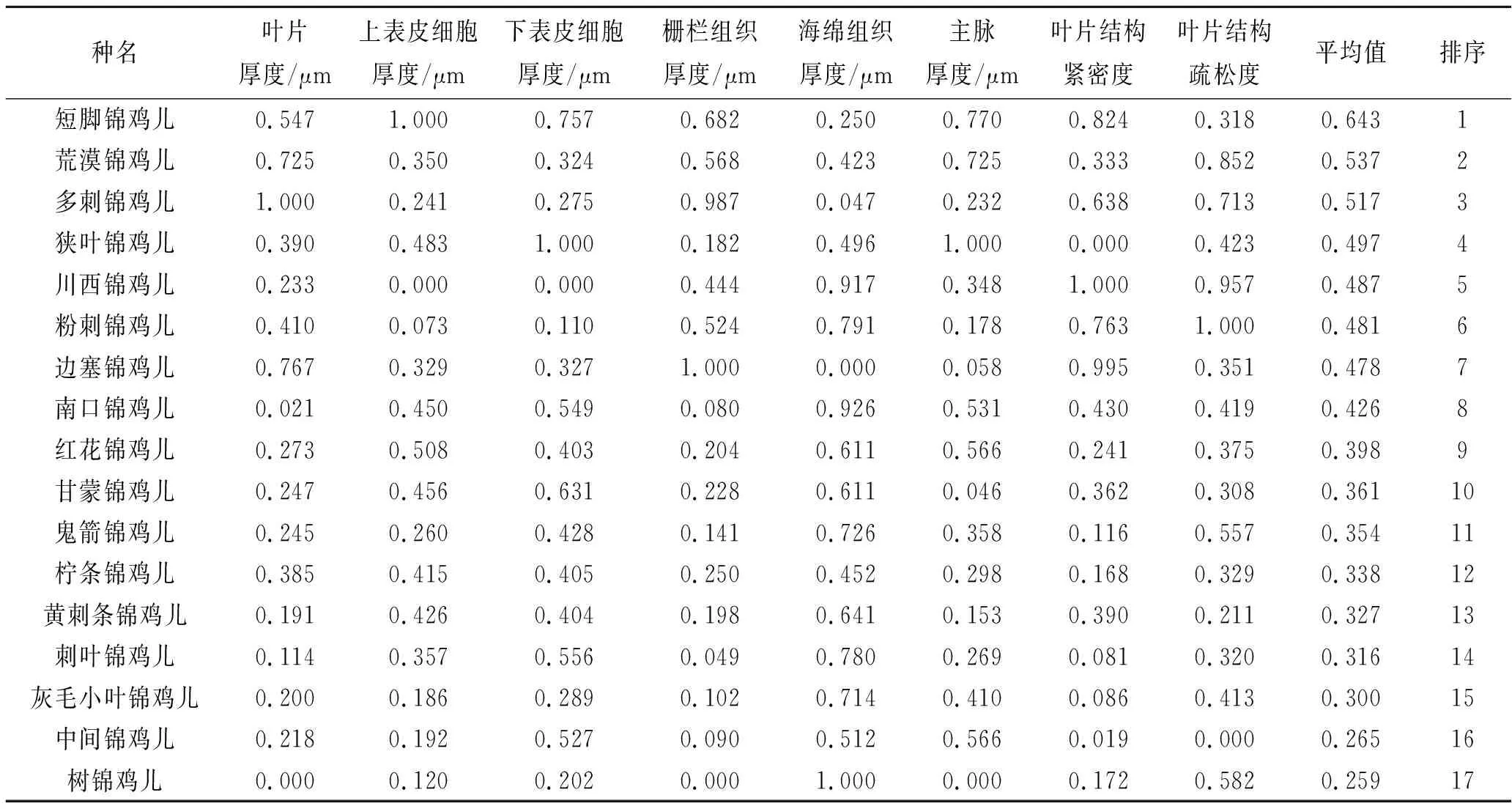
表4 17種錦雞兒屬植物隸屬函數值分析及排序
植物的抗旱性是一個復雜的、受多重因素影響的綜合特性[28],因此,錦雞兒屬不同植物的抗旱機制也不盡相同,每種植物僅憑某一項指標進行評價抗旱性具有片面性[22]。因此使用基于模糊數學原理的隸屬函數法進行抗旱性研究分析,其結果更加準確客觀。本研究結果顯示:短腳、荒漠和狹葉錦雞兒表現出較強的抗旱性,這與李蒙蒙等[1]基于葉片微形態結構評價錦雞兒屬植物的抗旱性得到的研究結果一致,且葉片厚度、表皮細胞厚度與主脈厚度隨著抗旱性的增強逐漸變厚;檸條錦雞兒比中間錦雞兒與灰毛小葉錦雞兒抗旱性強,這是由于檸條錦雞兒的葉片厚度與表皮細胞厚度更大,這與李晶[29]研究輸水結構與抗旱性所得出的結果一致;荒漠、中間、檸條和樹錦雞兒的抗旱性,與周萌[30]以形態適應特征和生理機制對錦雞兒研究所得出的結果不完全一致。這是因為植物在生長發育的過程中受到多種因素的影響,且植物適應干旱環境是一個復雜的形態特征和生理生化的協調過程。目前,檸條錦雞兒和中間錦雞兒已在典型荒漠帶沙地、荒漠化草原中大量種植,且取得了較好的生態效益[31];而本試驗結果表明:短腳錦雞兒、荒漠錦雞兒、狹葉錦雞兒和刺葉錦雞兒抗旱性強,可以在年降雨量少,蒸發量大的荒漠地區種植;川西錦雞兒、粉刺錦雞兒與南口錦雞兒也表現出了較強的抗旱性,在以后的干旱區植被恢復建設中種植,來增加群落物種的多樣性。
該研究對西北干旱地區植被建設與生態環境具有重要的作用。因此不論是依據形態特征還是生理特征來判斷植物的抗旱性,都有一定的局限性。不同錦雞兒植物的抗旱機制不同,因此在以后的研究過程中應綜合主要指標且從各方面來衡量每種植物的抗旱性。
4 結 論
17種錦雞兒屬植物葉肉解剖結構表現出3種不同類型,在抗旱性增強的過程中,葉肉結構從普通型、過渡型、環柵型依次過渡,在此過程中葉片逐漸變厚,柵欄組織、主脈越來越發達、葉片緊密度增強;聚類分析將17種錦雞兒依據各自不同特征聚為2類;根據隸屬函數法對17種錦雞兒抗旱能力大小進行排序,結果為:短腳錦雞兒>荒漠錦雞兒>多刺錦雞兒>狹葉錦雞兒>川西錦雞兒>粉刺錦雞兒>邊塞錦雞兒>南口錦雞兒>紅花錦雞兒>甘蒙錦雞兒>鬼箭錦雞兒>檸條錦雞兒>黃刺條錦雞兒>刺葉錦雞兒>灰毛小葉錦雞兒>中間錦雞兒>樹錦雞兒。
猜你喜歡
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
