狂犬病病毒HEP-Flury M基因重排在小鼠致病性和保護性研究
2020-05-07 13:22:46梅明珠楊先鋒彭嬌嬌林穎儀林志雄郭霄峰
中國人獸共患病學報 2020年3期
梅明珠,楊先鋒,龍 騰,張 瓊,趙 靜,田 欽,彭嬌嬌,羅 均,姜 賀,林穎儀,林志雄,郭霄峰
狂犬病(Rabies)是一種動物源性病毒疾病,由狂犬病病毒引起,感染家畜和野生動物,通過接觸感染性的材料傳播給人,通常是唾液、咬傷或者抓傷。最近也有報道指出母親在分娩時可將病毒傳播給嬰兒[1]。狂犬病存在于除南極洲外的所有大陸上的150個國家和地區,每年大約有60 000人死于狂犬病,其中95%的死亡發生在非洲和亞洲。我國也是狂犬病流行最嚴重的國家之一,發病率僅次于印度,居世界第2位。根據中國衛健委最新統計2018年我國發病人數為422例,死亡人數為410例,目前尚無可靠有效的治療方法,主要依賴疫苗的免疫接種進行預防。
狂犬病病毒作為單股負鏈RNA病毒,其基因順序高度保守即3′-N-P-M-G-L-5′,基因轉錄mRNA的轉錄數量取決于其在基因順序中的位置,離3′端啟動子的距離越遠,轉錄效率越低即leader RNA>N>P>M>G>L[2]。通過基因順序的重排能夠改變病毒的基因表達,從而改變病毒的表型。單股負鏈RNA病毒目中的另一個典型病毒水泡性口炎病毒(vesicular stomatitis virus,VSV),嚴格遵守這種轉錄模式,Wertz等通過重排VSV的基因順序,能夠改變病毒表型,獲得低致病性高免疫原性的病毒毒株[3-5]。
狂犬病病毒HEP-Flury(high-egg-passage Flury),作為一種高減毒狂犬病病毒固定株,已在日本作為狂犬病疫苗株[6],其轉錄具有自己的獨特模式。前期,我們已對其N和P基因進行了重排,并對基因重排株的表型變化進行了分析[7-8]。在本研究中,我們將對M基因重排株的致病性和免疫原性進行評估,從而找出更適合的狂犬病疫苗候選株。
1 材料與方法
1.1病毒和實驗動物 狂犬病病毒親本株rHEP-Flury、M基因位于基因順序2位的病毒M2(N-M-P-G-L)、M基因位于基因順序4位的病毒M4(N-P-G-M-L)由華南農業大學獸醫學院郭霄峰課題組通過反向遺傳操作系統拯救獲得[9]。
孕17~18 d的SPF級雌性昆明系小白鼠、6~8周齡的SPF級雌性昆明系成鼠均購自南方醫科大學實驗動物中心,許可證號SCXk (粵)2011-0015,其狂犬病病毒血清抗體檢測均為陰性,健康狀況良好,在華南農業大學實驗動物中心SPF條件下進行飼養管理和試驗。所有實驗程序均遵守NIH條例,并獲得華南農業大學動物福利委員會支持。
1.2乳鼠上的毒力試驗 由于狂犬病病毒HEP-Flury只對乳鼠有致死性而對成鼠沒有,因此選取1~3日齡的乳鼠來進行病毒的毒力測定[10]。首先將病毒rHEP-Flury、M2和M4的滴度都調整為1.0×105.5FFU/ mL,然后再進行10-1~10-6的10倍倍比稀釋。每種病毒取10-1~10-5這5個稀釋倍數,顱內接種1~3日齡的SPF級昆明系乳鼠,每只乳鼠接種20 μL,每種病毒接種12只乳鼠,同時設置RPMI 1640對照組。接種后每天觀察小鼠的狀態和死亡數量并記錄,持續觀察28 d,以Reed-Muench法計算病毒在乳鼠上的半數致死量(LD50)。
1.3活病毒對成鼠的攻毒保護試驗 在免疫前對6周齡SPF級昆明系小鼠進行眼角靜脈叢采血,收集于高壓滅菌的潔凈1.5 mL離心管中。然后將離心管置于37 ℃溫箱,1 h后再放入4 ℃冰箱過夜。取出離心管于4 ℃離心機3 000 r/min離心10 min,小心將上層血清轉入另一個潔凈的離心管后放入-80 ℃冰箱待檢。此血清既能作為檢測抗體時的陰性血清,又能進一步確認所購小鼠是否含有狂犬病病毒抗體。
小鼠采血后24 h,分別將50 μL的rHEP-Flury、M2和M4于后腿腓腸肌免疫接種小鼠,每種病毒接種3個劑量,分別是103FFU、104FFU和105FFU,每個劑量接種10只小鼠。同時以RPMI 1640作為對照。免疫后21 d采集血清,測定抗體,采血后第2 d,用乙醚將小鼠麻醉后顱內注射狂犬病病毒標準攻毒株CVS-24,每只小鼠注射30 μL,包含50個顱內注射LD50(50ICLD50/30 μL)的病毒液,接種后每天觀察小鼠的狀態和死亡數量并記錄,持續觀察28 d。
1.4成鼠血清抗體監測 6~8周齡的昆明系小鼠分成4組,每組5只。后腿腓腸肌肌肉接種rHEP-Flury、M2或M4,50 μL/只(105FFU/50 μL),同時設置RPMI 1640對照組。接種后,每周從小鼠眼角靜脈叢采血分離血清,56 ℃ 30 min滅活后利用試劑盒SerelisaRabies Ab Mono Indirect kit (Synbiotics, Lyon, France)進行抗體檢測,連續測定10周。
1.5統計分析 利用軟件GraphPad Prism 6.0進行作圖及統計分析。小鼠血清抗體水平差異顯著性分析利用2way ANOVA方法;小鼠存活率利用Kaplan-Meier曲線作圖表示,差異顯著性分析利用log-rank test 方法。 星號代表統計學差異:*P<0.05; **P<0.01; ***P<0.001; ****P<0.0001;檢驗水準α=0.05,表示差異具有統計學意義。
2 結 果
2.1病毒在乳鼠上的致死性 顱內注射接種乳鼠結果顯示:M2在乳鼠上的LD50約是rHEP-Flury的1.5倍,M4的3倍;rHEP-Flury在乳鼠上的LD50約是M4的2倍(圖1A)。說明當M基因從病毒基因的2位移至4位時,致死乳鼠的病毒量逐漸降低,揭示病毒毒力隨著M基因從2位移至4位逐漸增強。同時我們記錄了乳鼠死亡開始的時間,當接種劑量為104.5FFU/mL時,rHEP-Flury、M2和M4引起小鼠死亡開始的時間都是接種后第5 d;當接種劑量為103.5FFU/mL~101.5FFU/mL時,rHEP-Flury和M4引起小鼠死亡開始的時間基本一致,而M2引起小鼠死亡開始的時間推遲,特別是接種劑量為102.5FFU/mL時,推遲更明顯(圖1B)。說明當M基因從3位移至2位時,病毒引起乳鼠發病時間推遲。這與病毒在NA細胞上的擴散規律一致,因此我們推測病毒在NA細胞上的擴散主要影響了小鼠死亡時間。
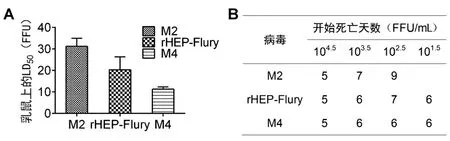
A:病毒在乳鼠上的LD50;B:病毒顱內接種后乳鼠死亡開始的時間圖1 rHEP-Flury和M基因重排病毒在乳鼠上的致死性Fig.1 Lethality of rHEP-Flury or rearranged viruses for suckling mice
2.2活病毒對成鼠的攻毒保護試驗 病毒rHEP-Flury、M2和M4在成鼠上的攻毒保護試驗結果顯示:當接種劑量為105FFU 和104FFU 時,M2、M4和rHEP-Flury免疫組小鼠的存活率差異沒有統計學意義,都顯著高于對照組RPMI 1640(圖2A、圖2B);當接種劑量為103FFU時,rHEP-Flury免疫組小鼠的存活率明顯低于M2和M4免疫組,與對照組RPMI 1640差異沒有統計學意義,此時M2和M4免疫組小鼠的存活率仍顯著高于對照組RPMI 1640(圖2C)。說明隨著接種劑量的降低,病毒對小鼠的保護率降低,其中rHEP-Flury組降低最明顯,M基因重排組降低不明顯。
接種后21 d血清抗體水平檢測顯示,小鼠血清抗體水平與病毒接種劑量呈正相關。當接種劑量由105FFU變為104FFU時,M2和M4組小鼠的血清抗體水平顯著降低;當接種劑量由104FFU變為103FFU時,rHEP-Flury組小鼠的血清抗體水平顯著降低(圖2D)。與此相符的是,我們發現在攻毒保護試驗中,當接種劑量由104FFU變為103FFU時,rHEP-Flury小鼠的存活率明顯降低。值得我們注意的是,當接種劑量由105FFU變為103FFU時,病毒M2和M4對小鼠的保護率沒有明顯降低,但是血清抗體水平顯著降低。

A-C: 小鼠存活率曲線;D:小鼠血清抗體水平圖2 攻毒保護試驗小鼠存活率及血清抗體水平Fig.2 Survial rates and antibody level in the protection of mice
2.3成鼠血清抗體水平 成鼠血清抗體水平監測結果顯示,M2、rHEP-Flury和M4免疫組小鼠的血清抗體水平在接種后1~3周快速升高,于第4周達到最高峰,然后快速降低,最后維持在一個高于保護水平0.6 EU/mL的水平(圖3)。在此我們推測抗體中和病毒的過程應起始于接種后第3周,主要發生于第4周,因為在接種后第3周至第4周,小鼠抗體水平的增速降低,且在第4周至第5周,小鼠抗體水平的降速最快。此外,我們發現雖然rHEP-Flury免疫組小鼠的血清水平最大值高于M2和M4免疫組小鼠,但是在5周后,M2和M4免疫組小鼠的血清抗體維持水平高于rHEP-Flury免疫組,這可能是因為M2和M4免疫組小鼠血清中有效清除病毒的抗體水平高。
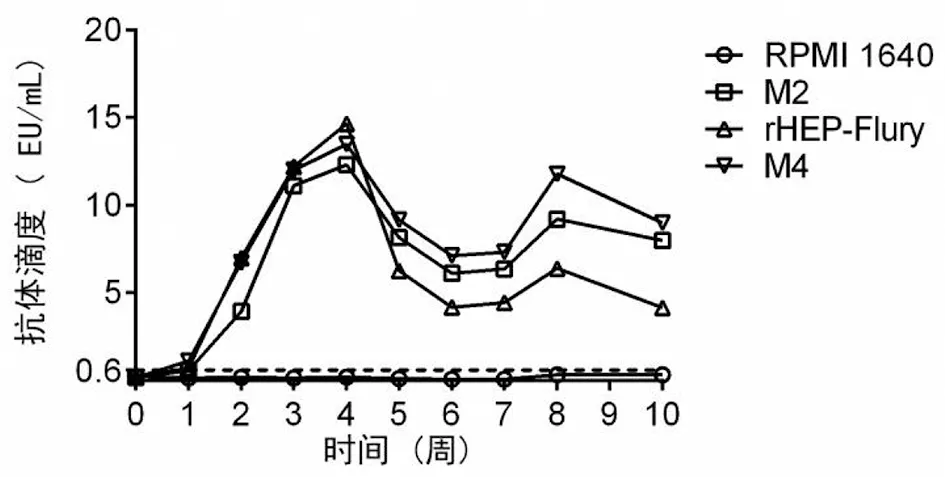
圖3 成鼠血清抗體水平的消長規律Fig.3 Antibody kinetics of viruses with M gene translocations
3 討 論
狂犬病病毒M蛋白主要調節病毒的轉錄和復制[11-12],協助G蛋白共同完成病毒粒子的出芽。Ben[13-14]等人研究發現M蛋白能夠抑制NF-κB依賴性的基因調節因子的表達,從而抑制細胞免疫反應,使病毒逃避宿主免疫系統的追蹤,但是疫苗株沒有這種能力[15]。因此,在本研究中,當M基因移至2位時,病毒毒力沒有增強。在前期的P基因重排研究中,我們發現P基因位置離啟動子位置越遠,病毒毒力越弱,不受其他基因位置被動改變影響[7],因此我們推測病毒M2的毒力降低,主要是因為M基因前移一位時,P基因被動后移一位。對于P基因位置一致的rHEP-Flury和M4,M4的毒力高于rHEP-Flury。病毒M4的M基因在后移一位時,G基因被動前移一位,前期研究表明非致病性狂犬病病毒G蛋白能夠誘導人類細胞調亡,從而限制病毒在中樞神經系統中擴散[16],且HEP-Flury的G蛋白也證實在高MOI情況下能夠誘導NA細胞發生凋亡[17-18],但是我們發現與rHEP-Flury相比,M4的毒力卻增強了。因此,我們推測基因重排只是改變了G蛋白的量而沒有改變其結構,對病毒在NA細胞中的擴散沒有顯著影響。在此,HEP-Flury 的M基因可能存在某種機制使得病毒毒力減弱,這還需要進一步研究確認。
狂犬病病毒疫苗株G蛋白能夠誘導機體產生中和抗體[19],N蛋白也能刺激機體Th細胞和中和抗體的產生,誘導機體產生細胞免疫[20]。在本研究中,我們發現當免疫接種劑量為103FFU時,雖然rHEP-Flury和M基因重排株產生的抗體水平差異沒有統計學意義,但是其對小鼠的保護率明顯低于M基因重排株,因此我們推測M基因重排株產生的有效抗體水平高于親本株,這在后面的抗體監測分析中可以看出。此外,是否M基因重排后能夠誘導更強的細胞免疫從而提高對機體的保護力還有待進一步研究。
綜上所述,在狂犬病病毒HEP-Flury的M基因重排研究中,我們發現M基因重排對小鼠的致病性和免疫原性都產生了影響,與親本株相比,M基因重排株M2具有更低的毒力和更高的保護性,可作為狂犬病疫苗候選株,為狂犬病疫苗的開發提供更好的選擇。
利益沖突:無
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
課堂內外·初中版(科學少年)(2023年10期)2023-12-10 00:43:06
全科護理(2022年10期)2022-12-26 21:19:15
中老年保健(2021年3期)2021-08-22 06:50:04
天津醫科大學學報(2021年2期)2021-03-29 05:31:08
國際放射醫學核醫學雜志(2021年10期)2021-02-28 08:41:58
現代臨床醫學(2021年1期)2021-01-26 00:56:02
學苑創造·A版(2020年9期)2020-10-13 09:41:02
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
中國合理用藥探索(2014年11期)2014-03-11 20:30:20
