鉀離子通道在黃曲霉菌孢子萌發(fā)中的作用
2020-05-11 05:08:52邵錚錚趙巖巖胡梁斌莫海珍李紅波
食品與生物技術(shù)學報 2020年2期
邵錚錚, 周 威, 趙巖巖, 胡梁斌,2, 莫海珍,2, 李紅波*,2
(1. 河南科技學院 食品學院,河南 新鄉(xiāng)453003;2. 陜西科技大學 食品與生物工程學院,陜西 西安710021)
黃曲霉菌(Aspergillus flavus,AF)作為一類重要的腐生病原真菌,其產(chǎn)生的次生代謝產(chǎn)物黃曲霉毒素B1(AFB1)是目前已知的最強化學致癌物之一,嚴重威脅著人類和動物的生命健康[1-2]。 黃曲霉菌廣泛存在于糧食及食品中,據(jù)AFO 統(tǒng)計全球每年超過30%的糧食作物受到真菌毒素污染, 造成數(shù)千億美元經(jīng)濟損失。 我國是世界上受黃曲霉污染最嚴重的國家之一[3]。 相關(guān)的流行病學研究顯示,AFB1 與肝癌發(fā)生密切相關(guān),對人類健康造成重大危害[4]。 此外, 黃曲霉能夠?qū)е略S多門類的動物感染曲霉病,降低或損傷機體的免疫系統(tǒng),尤其對于免疫缺陷病人感染侵襲性曲霉病往往是致命的[5-6]。 人們一直在尋求對其進行有效防治的策略和技術(shù),包括良好的田間管理、生化防治以及物理化學脫毒方法等[7-9]。但是由于黃曲霉菌污染的發(fā)生風險覆蓋了從田間原料生產(chǎn)到后期產(chǎn)品消費的整個過程,其控制依舊是亟需解決的世界性難題。 從危害發(fā)生過程來看,黃曲霉危害循環(huán)經(jīng)歷了從孢子萌發(fā)、侵染定植、生長產(chǎn)毒、產(chǎn)孢擴散等過程。 從危害循環(huán)上游制定控制黃曲霉菌策略可以更有效的緩解黃曲霉污染的發(fā)生。
黃曲霉菌以無性孢子作為主要傳播體。 而孢子作為休眠體需要被喚醒萌發(fā)后,才能生長發(fā)育并進行毒素代謝等次生代謝或者滲透并侵害人體組織導致肺曲霉病發(fā)生[10]。 因此黃曲霉孢子萌發(fā)可以作為黃曲霉危害發(fā)生過程中最上游的生理過程。 孢子萌發(fā)第一步就是感知外界環(huán)境,進而將信號傳遞到胞內(nèi)啟動萌發(fā)過程[11]。因此,尋找孢子感知環(huán)境萌發(fā)所依賴的關(guān)鍵因子作為靶標,從最上游阻斷黃曲霉孢子萌發(fā),是有效控制黃曲霉污染、開發(fā)新型抗菌藥物的突破口之一。
作者研究了K+對黃曲霉菌孢子萌發(fā)的影響,通過使用K+特異性熒光探針PBFI-AM 研究胞內(nèi)K+狀況。 鑒于K+與黃曲霉孢子萌發(fā)相關(guān),進一步研究了特異性電壓門控鉀離子通道阻滯劑4-氨基吡啶(4-Aminopyridine,4-AP)對孢子萌發(fā)的影響。 通過生物信息學預測分析了可能的電壓門控鉀離子通道蛋白KCNA,并構(gòu)建KCNA 非洲爪蟾卵母細胞表達體系,使用雙電極電壓鉗系統(tǒng)觀察了KCNA 的電生理學特性。 通過確認KCNA 電壓門控鉀離子通道特性,以期為進一步研究黃曲霉孢子萌發(fā)調(diào)控機制提供理論基礎(chǔ)。
1 材料與方法
1.1 黃曲霉孢子懸浮液的制備
菌株為黃曲霉菌CGMCC3.2890,購于中國普通微生物菌種保藏管理中心。 PDA 培養(yǎng)基4 ℃保存。取菌株接種于固體沙氏培養(yǎng)基平板上, 放置30 ℃恒溫培養(yǎng)箱中靜置培養(yǎng)2~3 d, 加入適量體積的0.1%Tritonx-100 于菌板中輕輕晃動使黃曲霉孢子均勻分散于溶液中, 再將孢子液吸取至離心管中,用血球計數(shù)板于顯微鏡下計數(shù),用液體沙氏培養(yǎng)基稀釋至所需孢子液濃度,孢子液現(xiàn)用現(xiàn)配。
1.2 KCl 脅迫及4-AP 處理對黃曲霉孢子萌發(fā)的影響
在蒸餾水中加入不同濃度的KCl,通過96 孔板培養(yǎng)實驗發(fā)現(xiàn),于30 ℃振蕩培養(yǎng),每隔2 小時用顯微鏡觀察拍照,統(tǒng)計孢子萌發(fā)率。使用4-AP 阻斷鉀離子通道活性,觀察其對孢子萌發(fā)的影響。 使用96孔板,在培養(yǎng)基中加入10 mmol/L 4-AP 處理野生型菌株孢子,于30 ℃振蕩培養(yǎng),每隔2 小時用顯微鏡觀察拍照,統(tǒng)計孢子萌發(fā)率。
進一步采用K+熒光探針檢測KCl 處理對黃曲霉菌萌發(fā)早期胞內(nèi)K+含量的影響。取黃曲霉孢子懸液(1×105個/mL),添加1 μL PBFI-AM K+熒光探針(體積比1 000∶1),30 ℃、120 r/min 避光搖培孵育60 min,孵育后孢子液于4 ℃放置30 min。之后使用黑色酶標板,每孔添加200 μL 孢子液開始檢測,50 s后添加KCl 母液,終濃度分別為0、0.01、0.1、1 mmol/L。該過程中,一直使用多功能酶標儀檢測該孔內(nèi)熒光強度,每5 秒一次,共檢測380 次。 激發(fā)波長和發(fā)射波長為340 nm 和500 nm。 以相對熒光強度(RFU)為縱坐標,時間為橫坐標作圖。
1.3 鉀離子通道蛋白生物信息學分析
借鑒人體電壓門控鉀離子通道α 亞基(GenBank: AAP46292.1)序列,在黃曲霉菌基因組數(shù)據(jù)庫進行BLAST 檢索, 得到其同源蛋白質(zhì)(XP_002376854.1),將其命名為KCNA。篩選已報道的不同物種電壓門控鉀離子通道α 亞基蛋白,通過NCBI 數(shù)據(jù)庫查找了各自序列信息, 使用DNAMAN軟件進行序列比對分析。 使用TMHMM 軟件TMHMM (http://www.cbs.dtu.dk/services/TMHMM/)對蛋白質(zhì)進行跨膜預測。 利用http://zhanglab.ccmb.med.umich.edu/I-TASSER/網(wǎng)站對KCNA 蛋白質(zhì)三維結(jié)構(gòu)進行模擬。 利用在線生物信息學網(wǎng)站http://services.mbi.ucla.edu/SAVES/對其三維結(jié)構(gòu)模型進行綜合評價。在zinc 數(shù)據(jù)庫(http://zinc.docking.org/)中下載4-AP(zinc_599985)分子的sdf 文件,利用Autodock 軟件進行KCNA 蛋白與4-AP 對接研究[12]。
1.4 KCNA 異源表達及電生理學測定分析
非洲爪蟾卵母細胞處理:取非洲爪蟾冰凍麻醉后,下腹部切口取1cm2左右的卵巢小葉,浸于無Ca2+的OR2 液(82.5 mmol/L NaCl,2.5 mmol/L KCl,1 mmol/L MgCl2,1 mmol/L Na2HPO4,5 mmol/L HEPES,50 U/mL青霉素,50 μg/mL 鏈霉素,pH 7.5)的培養(yǎng)皿中。 室溫下在含2 g/L 的膠原酶溶液中振蕩消化1.5~2 h后,用OR2 液反復沖洗,去除殘留于卵母細胞表面的膠原酶。 挑選出外型較圓、色澤清晰、有一定張力、細胞表面光滑、無殘留纖維組織及毛細血管的V、VI 期卵母細胞,置于含ND96 液培養(yǎng)皿中。 使用微量注射儀將KCNA 的cRNA 注射到卵母細胞中,每個細胞按2~5 ng/5 nL 注射。
制備cRNA:本研究使用pGEMHE 質(zhì)粒,引物序列見表1。采用快速克隆技術(shù),連接載體pGEMHE和KCNA 基因片段。 使用DpnⅠ限制性內(nèi)切酶線性化處理后, 將載體pGEMHE 和KCNA 混合物轉(zhuǎn)化到感受態(tài)細胞DH5α 內(nèi),最后挑取單克隆進行單酶切及測序鑒定[13]。 使用反轉(zhuǎn)錄試劑盒將線性化擴增產(chǎn)物體外轉(zhuǎn)錄得到cRNA。
雙電極電壓鉗測定: 注射后的卵母細胞在含2.5 mmol/L 丙酮酸鈉的ND96 中20 ℃培養(yǎng)2~3 d后進行電生理學測定。 電極內(nèi)灌注1 g/dL 瓊脂糖、3 mol/L 的KCl,電極電阻<1 MΩ。 細胞外液成分為ND96, 外源脅迫因子4-AP 使用基礎(chǔ)細胞灌流液ND96 調(diào)配。 實驗在25 ℃進行,數(shù)據(jù)采集和分析采用pClamp9.0、Clampfit9.0 和Origin9.1 完成。

表1 引物序列表Table 1 Primers list
2 結(jié)果與分析
2.1 KCl 脅迫及4-AP 處理對黃曲霉孢子萌發(fā)的影響
在蒸餾水中加入不同濃度的KCl,通過96 孔板培養(yǎng)實驗發(fā)現(xiàn),低濃度KCl(0.1 mmol/L)脅迫下,隨著外界KCl 濃度的增加(0~0.1 mmol/L),黃曲霉孢子萌發(fā)率呈現(xiàn)上升趨勢,在0.1 mmol/L 達到最高值,見圖1。 而隨著KCl 濃度的繼續(xù)增加,黃曲霉孢子萌發(fā)率則呈現(xiàn)下降趨勢,高濃度的KCl 脅迫(1~50mmol/L)顯著抑制孢子萌發(fā)。

圖1 不同濃度KCl 對黃曲霉孢子萌發(fā)的影響Fig. 1 Effect of KCl on spore germination of A.flavus
進一步采用K+熒光探針檢測KCl 處理對黃曲霉菌萌發(fā)早期胞內(nèi)游離K+濃度的影響。 結(jié)果見圖2。0.1 mmol/L KCl 處理后, 胞內(nèi)游離鉀離子濃度迅速升高,與圖1 孢子萌發(fā)數(shù)據(jù)相吻合,可見適合孢子萌發(fā)的特定濃度K+能夠激發(fā)黃曲霉孢子產(chǎn)生瞬間的高鉀離子流,K+響應為黃曲霉孢子萌發(fā)的早期信號。
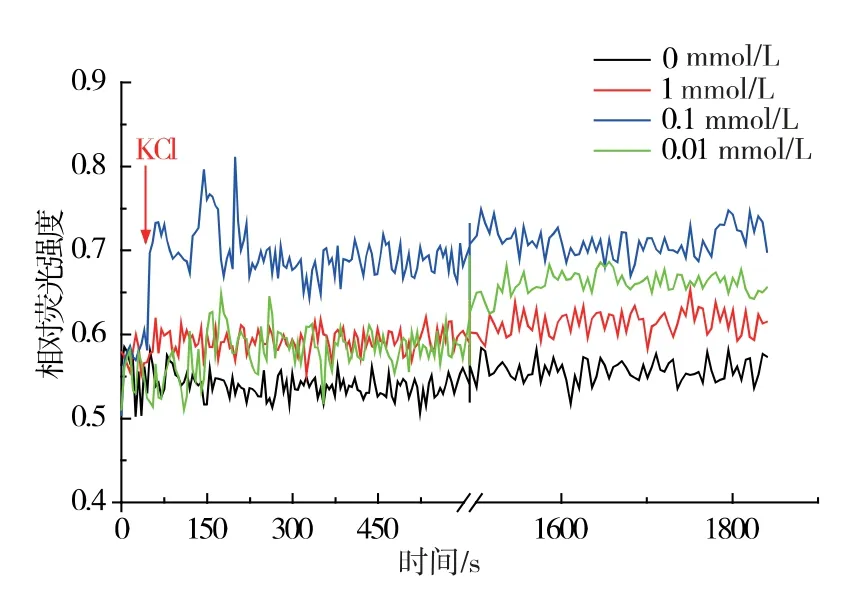
圖2 不同濃度KCl 誘導黃曲霉孢子內(nèi)鉀離子的改變Fig. 2 Changes of K + was induced by different KCl concentrations in spores of A. flavus
4-AP 是一種特異性電壓門控鉀離子通道阻斷劑,我們初步使用4-AP 阻斷鉀離子通道活性,觀察其對孢子萌發(fā)的影響。 使用96 孔板,在培養(yǎng)基中加入10 mmol/L 4-AP 處理野生型菌株孢子, 于30 ℃振蕩培養(yǎng),每隔2 小時用顯微鏡觀察拍照,統(tǒng)計孢子萌發(fā)率。結(jié)果發(fā)現(xiàn)10 mmol/L 4-AP 處理能夠明顯抑制并延遲孢子的萌發(fā),見圖3。

圖3 4-AP 處理對黃曲霉孢子萌發(fā)的影響Fig. 3 Effect of 4-AP treatment on the spore germination of A.flavus
2.2 鉀離子通道蛋白KCNA 預測及建模
篩選已報道的不同物種電壓門控鉀離子通道α亞基蛋白,通過NCBI 數(shù)據(jù)庫查找了各自序列信息,使用DNAMAN 軟件進行序列比對分析。 結(jié)果發(fā)現(xiàn),黃曲霉菌KCNA 的423-FAFCSLLTIGYGDITPTT-440 區(qū)序列與智人、秀麗隱桿線蟲、原雞等電壓門控鉀離子通道α 亞基蛋白孔區(qū)片段同源性較高,都擁有保守的TxGYGD 鉀離子選擇器特征片段,見圖4(a)。 紅色框表示在序列中一樣的氨基酸殘基。主要物種信息如下:Hsap = [智人,Homo sapiens,(AAP46292.1)];Ppen=[Polyorchis penicillatus,(AAB39750.1)];Cele = [秀麗隱 桿 線 蟲,Caenorhabditis elegans,(AAB95119.1)];Ggal = [原雞,Gallus gallus,(AAP94023.1)]。
使用TMHMM 軟件對KCNA 蛋白進行跨膜預測,發(fā)現(xiàn)其有明顯的6 個跨膜區(qū)域,見圖4(b)。同時胞內(nèi)還有較短的N 端和較長的C 端,符合典型的鉀離子通道蛋白特性。 利用http://zhanglab.ccmb.med.umich.edu/I-TASSER/網(wǎng)站對KCNA 蛋白三維結(jié)構(gòu)進行模擬,KCNA 三維結(jié)構(gòu)模擬圖見圖4(d)。利用在線生物信息學網(wǎng)站 http://services.mbi.ucla.edu/SAVES/對其三維結(jié)構(gòu)模型進行綜合評價。三維結(jié)構(gòu)模型綜合評價Ramachandran plot 分析發(fā)現(xiàn),KCNA三維結(jié)構(gòu)97.8%Φ-Ψ 二面角分布在允許區(qū)域, 見圖4(c)。不允許范圍的比例非常小,說明三維結(jié)構(gòu)中氨基酸的二面角是合理的。 證明模擬的KCNA 三維結(jié)構(gòu)合理,可以用于下一步分子對接。
在zinc 數(shù)據(jù)庫(http://zinc.docking.org/)中下載4-AP(zinc_599985)分子的sdf 文件,利用Autodock軟件進行KCNA 蛋白與4-AP 對接研究, 對接結(jié)果見圖4(e)。 4-AP 可作用于KCNA 鉀離子選擇器區(qū)域附近, 與403 位半胱氨酸、419 位谷氨酰胺、468位蘇氨酸以氫鍵方式結(jié)合, 這與4-Ap 可以抑制孢子萌發(fā)的數(shù)據(jù)相吻合,進一步支持了KCNA 作為電壓門控鉀離子通道的可能性。
2.3 KCNA 電生理學測定
將KCNA 克隆到質(zhì)粒載體pGEMHE 中, 成功構(gòu)建蛙卵細胞表達體系。將細胞置于ND96 溶液中,細胞膜電位鉗制在-40 mV,去極化至+60 mV,階幅為20 mV,刺激時間為500 ms。實驗發(fā)現(xiàn),有一個持續(xù)的外向電流出現(xiàn),并隨去極化程度增加而增大, 見圖5。當從鉗制電位-40 mV 去極化至+60 mV 時,可記錄到最大幅度的外向電流。 鉀離子通道抑制劑4-AP對該外向電流有明顯的抑制作用,使用10 mmol/L 4-AP處理可以顯著的阻斷KCNA 電流,結(jié)果進一步證實了KCNA 電壓門控鉀離子通道蛋白屬性。 前述4-AP 能夠抑制黃曲霉孢子萌發(fā), 可見鉀離子通道蛋白與孢子萌發(fā)密切相關(guān)。

圖4 鉀離子通道KCNA 生物信息學分析Fig. 4 Bioinformatics analysis of potassium channel KCNA

圖5 鉀離子通道阻滯劑4-AP 阻斷KCNA 外向電流Fig. 5 4-AP(potassium ion channel blocker)blocking the KCNA outward current
3 結(jié) 語
本研究發(fā)現(xiàn),一定濃度的胞外KCl 能夠影響黃曲霉孢子的萌發(fā)。 通過使用K+特異性熒光探針PBFI-AM,發(fā)現(xiàn)適合孢子萌發(fā)的特定濃度K+能夠激發(fā)黃曲霉孢子瞬間產(chǎn)生較高的鉀離子流,說明K+響應可能是黃曲霉孢子萌發(fā)的早期信號。 4-AP 作為特異性電壓門控鉀離子通道阻滯劑,能夠明顯抑制并延遲孢子的萌發(fā)。 這些結(jié)果表明,電壓門控鉀離子通道可能參與黃曲霉孢子萌發(fā)。 借鑒人體電壓門控鉀離子通道α 亞基序列,在黃曲霉菌基因組數(shù)據(jù)庫進行BLAST 檢索,得到其同源蛋白KCNA。 通過對KCNA 進行序列分析,發(fā)現(xiàn)其423-FAFCSLLTIG YGDITPTT-440 區(qū)序列與智人、秀麗隱桿線蟲、原雞等電壓門控鉀離子通道α 亞基蛋白鉀離子選擇器區(qū)域序列同源性較高,擁有明顯的鉀離子選擇器特征。利用Autodock 軟件進行KCNA 與離子通道阻斷劑4-AP 的對接研究, 對接結(jié)果支持了KCNA 作為電壓門控鉀離子通道α 亞基的可能性。研究證實4-AP 可以抑制孢子萌發(fā), 這說明鉀離子通道在黃曲霉孢子萌發(fā)中起到重要作用。 通過構(gòu)建KCNA 非洲爪蟾卵母細胞表達體系,用雙電極電壓鉗系統(tǒng)測得KCNA 外向型電流, 一定程度上證明了其電壓門控鉀離子通道蛋白屬性。
細胞內(nèi)K+穩(wěn)態(tài)是多數(shù)生物代謝機制和綜合性能優(yōu)化運行的先決條件,胞內(nèi)K+動態(tài)平衡由細胞質(zhì)和細胞膜上的許多K+通道和轉(zhuǎn)運蛋白所調(diào)控[14]。 在孢子萌發(fā)早期階段,膜通透性增加,導致孢子內(nèi)離子和水重新分配,孢子內(nèi)80%的K+快速釋放到孢子外,隨后K+依賴能量被再次吸收[15]。 而離子的移動往往涉及到離子通道的響應,這也是細胞識別外界信號的一個重要途徑。 阻斷離子通道能夠抑制外源信號的細胞內(nèi)傳導,研究發(fā)現(xiàn)使用鉀離子通道抑制劑四乙胺能夠完全抑制花粉或無性孢子的萌發(fā)[16]。電壓門控鉀離子通道是目前存在最廣泛且最復雜的一大類鉀離子通道, 是藥物設計的重要靶標之一。 Gobert 等通過敲除和過表達擬南芥中鉀離子通道TPK1, 發(fā)現(xiàn)TPK1 對保持胞內(nèi)K+動態(tài)平衡起著重要作用,能夠影響種子的萌發(fā)、生長和氣孔運動[17]。敲除擬南芥花粉管K+通道AKT6 后,花粉管萌發(fā)率與野生型相比大幅度降低, 并且AKT6 對外界pH變化非常敏感[18]。 然而關(guān)于真菌電壓門控鉀離子通道的相關(guān)文獻較少,黃曲霉孢子萌發(fā)過程中是否涉及到電壓門控鉀離子通道更是尚無研究報道。 本研究為從根源上防控黃曲霉菌污染,尋找抗性靶標提供了一個新思路。
