哺乳動物細胞中糖鏈結構的改造
2020-05-11 05:08:52金則成喜多島敏彥藤田盛久高曉冬
食品與生物技術學報 2020年2期
金則成, 喜多島敏彥, 藤田盛久, 高曉冬
(江南大學 生物工程學院,江蘇 無錫214122)
生物藥物分子是一類由生物體包括細菌、酵母、昆蟲、植物和哺乳動物細胞生產的藥物。 藥物蛋白質如單克隆抗體具有高特異性和與靶分子的高親和性, 同時與傳統藥物相比具有更低的副作用。目前,重組藥物蛋白質已經在治療疑難雜癥如癌癥和自身免疫疾病方面變得越來越重要[1-3]。 目前,大部分藥物蛋白質主要由哺乳動物細胞合成,這主要是由于哺乳動物細胞生產蛋白質可以提升正確折疊蛋白質的產量和促進翻譯后蛋白質的折疊[4-5]。 由于大多數分泌蛋白質具有糖鏈,為了維持蛋白質的穩定性和功能以及抵御免疫反應,適當的糖鏈是非常必要的。
使用哺乳動物細胞生產生物藥物分子依然有幾個缺點,包括高成本和糖鏈的不均一性。 為了確保用作為生物藥物分子的蛋白質質量和穩定性,糖鏈的不均一性是一個必須要解決的問題。 舉例來說,紅細胞生成素和粒細胞集落刺激因子(G-CSF)之類的細胞因子在體外必須具有含唾液酸的復合型糖鏈才具有活性[6-9]。 同時,對蛋白質的糖鏈結構進行改造可以提高蛋白質的功能活性。 舉例來說,具有移除巖藻糖結構的N-糖鏈的抗體可以大幅提高抗體依賴性的細胞毒性(ADCC)作用[10-11]。 現在,為了生產抗體,不進行巖藻糖修飾的細胞已經被頻繁地使用。 可以生產均一的糖蛋白的哺乳動物細胞株的構建是目前生物醫藥品生產領域中最迫切的需求。
圖1 展示了糖鏈在哺乳動物細胞中的合成途徑,糖蛋白首先在內質網中合成,隨后被轉運到高爾基體中,蛋白質上的糖鏈在α-1,2 甘露糖苷酶的催化下轉變成Man5-GlcNAc2 型結構的糖鏈。 目前獲得Man5-GlcNAc2 型結構的糖鏈主要有兩種方法:一種是利用N-乙酰葡糖胺轉移酶I(GnT-I)編碼基因MAGT1 的突變或敲除細胞生成, 然而這種方法的缺點是該細胞株不能產生Man9-GlcNAc2或Man8-GlcNAc2 型結構的糖鏈;另外一種方法就是通過化學試劑幾夫堿(kifunensine)或脫氧野尻霉素(deoxynojirimycin)抑制細胞α-1,2 甘露糖苷酶的活性,缺點是如果想要獲得穩定均一的糖型,需要用化學試劑持續處理細胞。
溶酶體中含有多種水解酶,這些水解酶大部分是帶有糖鏈的糖蛋白,它們可以將蛋白質、粘多糖、糖脂等物質消化為小分子,提供細胞再次回收利用[13]。這些水解酶在內質網中合成,在高爾基體內進行糖鏈的修飾,然后通過特定的M6PR 受體的識別被轉運至溶酶體中。 高爾基體中糖鏈的修飾往往是通過在核心糖鏈的Man 的6 位上添加UDP-N-乙酰葡萄糖胺(UDP-GlcNAc)的N-乙酰葡萄糖胺-1-磷酸酯部分(GlcNAc-1-P),產生Man-6-P-1-GlcNAc,然后將GlcNAc 的部分除去, 形成具有酸性糖鏈的糖蛋白,然后通過特定的M6PR 受體的識別被轉運至溶酶體中[14-16]。

圖1 哺乳動物細胞N-糖鏈合成途徑Fig.1 Pathway of N-glycan synthesize in mammalian cells
當產生的水解酶因為代謝通路異常無法正常運送至溶酶體或者是因控制該溶酶體酶的基因發生突變,該酶反應鏈中的中間產物不能正常降解而在溶酶體中貯積,從而引起細胞組織器官功能的障礙,導致溶酶體貯積癥的發生[17-18]。 例如,法布萊病的病人由于缺少α 半乳糖苷酶,使得糖脂特別是一種中間產物globortriaosylceramide(Gb3)無法被分解而蓄積在細胞的溶酶體中, 從而威脅著他們的生命。 目前對于溶酶體貯積癥主要的療法有:酶替代療法、化學療法、基因水平上的基因修飾療法等,而其中最為經典的方法即為酶替代療法。 由于細胞膜表面存在M6PR,M6PR 可以識別藥物蛋白質上的糖鏈結構并將蛋白質帶到溶酶體, 從而能夠通過M6PR 用正常的水解酶替代自身損傷的水解酶,使溶酶體貯積癥得到改善[19-20]。
因糖鏈的不均一性而導致的糖蛋白的不均一性會對糖蛋白的生產及其應用產生不良影響。 由于M6PR 對于糖蛋白具有專一識別性, 當糖蛋白中存在部分糖鏈結構并非高甘露糖型或者說不存在6位被磷酸化的糖鏈時,會導致M6PR 對于藥物的吸收效率的降低,導致治療效率不高[21-22]。 另外,當糖鏈存在不均一的情況時,因為糖鏈的結構還可能導致糖蛋白被機體識別為外來的抗原物質,從而引起免疫反應。 出于藥物分子的安全性的考慮,需要盡量確保糖鏈的均一性。
本研究可提供一種可用于生產均一性藥物糖蛋白的細胞株, 即敲除MAN1A1 和MAN1A2 的雙敲除細胞株Double-KO 細胞株。該細胞株中高甘露糖型N-糖鏈所占比例增加,同時由于復合型N-糖鏈的下降也可以證明該細胞株內糖鏈的表現形式更均一。
1 材料與方法
1.1 細胞培養
人體胚胎腎細胞HEK293 使用10%胎牛血清(FCS) 的DMEM 培養基 (Gibco,Life technologies,USA)培養,培養條件為37 ℃、5%。
1.2 高爾基體α-甘露糖苷酶的基因敲除質粒的構建
利 用 CRISPR -Cas9 (Clustered Regularly Interspaced Short Palindromic Repeats)技術[23]敲除基因往往需要設計一個20 bp 長度的序列片段, 且該序列片段后要有一個PAM 位點(NGG/NAG)。 在本實驗中,需要敲除的兩個基因MAN1A1/MAN1A2 的基因序列從NCBI 上下載得到。 關于guide-RNA 的設計, 在Michael Boutros lab's Target Finder(http://www.e-crisp.org/E-CRISP/designcrispr.html) 上找到敲除基因所需的guide-RNA 的DNA 序列。 通過網站設計可以獲得MAN1A1 的兩條靶序列及各自所使用的引物序列為:
MAN1A1KO1:AAAACCACGAGCGGGCTCTCAGG PrimerKO1F:caccAAAACCACGAGCGGGCTCTC PrimerKO1R:aaacGAGAGCCCGCTCGTGGTTTT MAN1A1KO2:CCACCTTCTTCTTCTCCAGTAGG primerKO2F:caccCCACCTTCTTCTTCTCCAGT
primerKO2R:aaacACTGGAGAAGAAGAAGGTGG MAN1A2 的兩條靶序列及各自所使用的引物序列為:
MAN1A2KO1:CCTTTACCGGCATCTACATGTGG rimerKO1F:caccCCTTTACCGGCATCTACATG rimerKO1R:aaacCATGTAGATGCCGGTAAAGG AN1A2KO2:CATGGATCAGGAAGACTCCGGGG rimerKO2F:caccCATGGATCAGGAAGACTCCG rimerKO2R:aaacCGGAGTCTTCCTGATCCATG。如圖2 所示,將含有CRISPR-Cas9 系統的質粒pX330-EGFP 使用Bbs1(NEB:R0539S)切開,并使用Mighty Mix 將設計好的靶序列與質粒上guide-RNA 的DNA 序列序列相連接, 構建為含有MAN1A1/MAN1A2 靶點的質粒, 并將它們命名為:pX330 -EGFP -MAN1A1KO1/pX330 -EGFP -MAN1A1KO2、pX330-EGFP-MAN1A2KO1/ pX330-EGFP-MAN1A2KO2。
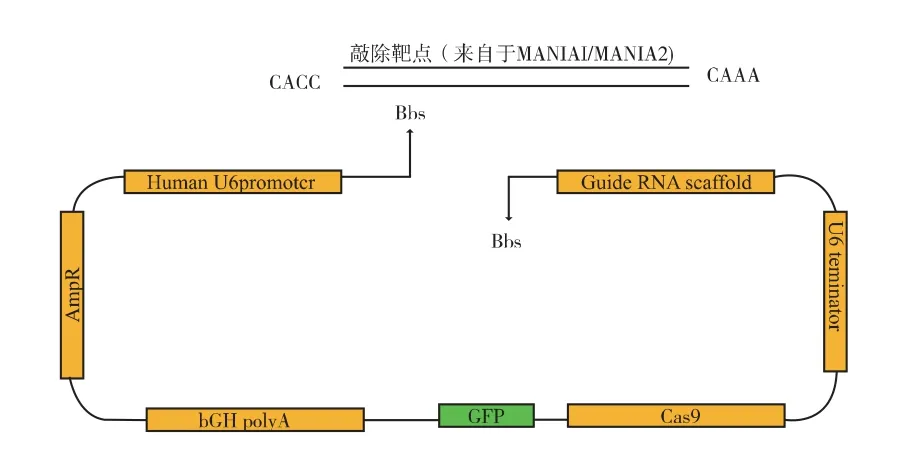
圖2 CRISPR/Cas9 敲除質粒的構建Fig. 2 Construction of CRISPR/Cas9 KO plasmid
1.3 轉染
將野生型細胞HEK293 使用10%FCS 的培養基培養過夜, 待其生長到約90%~95%時進行轉染。轉染試劑使用PEI-MAX[24](2 mg/mL pH 7.5),在轉染前先需要將PEI-MAX 與OPTI(life technologies:31985-070)混合均勻,其比例為1 μL PEI-MAX∶50 μL OPTI 培養基。將敲除所需質粒和攜帶抗性基因的質粒pME-puro 與OPTI 培養基混合均勻,質粒添加量的比例為4 μg DNA∶5 μL PEI-MAX。將PEIMAX 溶液和含質粒溶液混勻,常溫放置25 min,使質粒與PEI-MAX 結合。 之后將混合的溶液加入野生型細胞株的培養基中。 12 h 更換新鮮培養基,待生長恢復后(約24 h),更換為質量濃度1 μg/mL 的嘌呤霉素的培養基進行篩選。
1.4 單克隆細胞的獲得與驗證方法
篩選得到的細胞含有抗性質粒和敲除質粒,使用限制性稀釋使單個細胞生長于96 孔板中, 獲得單克隆細胞。 當細胞數量增加后將單克隆細胞轉移至12 孔板培養。 當其生長為100%狀態時,移去培養基,使用PBS 沖洗一次,加入胰蛋白酶(Sangon,CN)消化細胞,加入含10%FCS 的DMEM 培養基收獲細胞。 所得細胞液于3 000 r/min 離心2 min,并再次使 用 磷 酸 緩 沖 液 (phosphate buffered saline,PBS,Sangon,CN) 沖洗得到沉淀物。 在沉淀物中加入50 mmol/L NaOH 并于金屬浴中95 ℃反應20 min,反應結束后加入1 mol/L 的Tris (pH 7.5) 于15 000 r/min離心3 min,取上清液待用。 驗證方法使用測序驗證的方式,將提取的基因組使用驗證設計的驗證引物進行PCR 驗證反應, 反應后的PCR 產物寄送華大基因測序公司進行測序。 設計的驗證引物為:
check MAN1A1F:TTCCTGCCAGACTCCTCCAAG;check MAN1A1R:CCACTCACCTCTTTGATCTTTGC C;check MAN1A2F:CTTAGTGCCTTCATCACTCTGT GT;check MAN1A2R:GACAGATTCGATCCAATCAC CGT。
1.5 流式細胞分析
細胞被胰蛋白酶消化處理后收集細胞液,用磷酸緩沖液洗滌1~2 次。 取5×105個細胞重懸于流式細胞分析液FACS(PBS,1% BSA 和0.1% )中,加入帶有熒光標記的凝集素, 如PHA-L4-FITC 和ConA-FITC(J-oil mills,Tokyo,Japan),使其終濃度為1%,反應約15 min。 使用3 000 r/min 離心3 min后棄去上清液, 使用FACS 洗滌2 次后再次離心移去上清液并重懸于FACS 溶液中。 使用BD Accuri C6(BD,USA)流失細胞分析儀進行分析。
1.6 細胞株中LAMP2 蛋白糖鏈變化的免疫印跡法驗證
將細胞通過細胞裂解液裂解, 置于冰上孵育30 min。將細胞裂解提取液中蛋白質變性,使用糖類內切酶H(Endo-H,New England BioLabs)或PNGase F于37 ℃處理3 h。 處理后的蛋白質樣品在SDS-PAGE中上樣, 然后轉膜到PVDF 膜上。 使用鼠單克隆anti-LAMP2 抗體用于檢測細胞裂解提取液中的LAMP2 蛋白質[25]。
1.7 細胞生長曲線的測定
將細胞消化后使用細胞計數器計數。 將所得的細胞稀釋至2 000 個/dL 的細胞濃度,將該濃度的細胞分裝至96 孔板中,每孔體積為100 uL,將細胞放置于37 ℃、5%的條件下培養。 6 h 后在每孔內加入CCK-kit(Dojingo,Japan)后培養箱內培養1 h 后,在450 nm 納米波長下測定吸光度變化[26],該結果即為0 h 數據, 之后每隔24 小時重復使用CCK-kit 測定,記錄下0、24、48、72、96、120、144 h 的數據。
2 結果與分析
2.1 MAN1A1 或MAN1A2 的單敲除細胞的構建
當哺乳動物細胞表達糖蛋白時由于發生于內質網和高爾基體的糖基化過程會產生在蛋白質上糖鏈的異質性。 糖鏈的異質性是一個醫藥用糖蛋白生產中的一個重大的問題。 為了使糖鏈均一化,我們選擇使用人胚胎腎細胞HEK293 用于如病毒的生產和表達重組蛋白質。 由于該細胞株為頻繁使用的人源細胞株,它具有如染色體數目和核苷酸多態性等完備的基因組信息,同時積累了大量應用基因表達時的數據。 同時補充說明,在如中國倉鼠卵巢細胞(CHO)中已經建立了懸浮培養和無血清培養的方法,這些在構建表達糖鏈均一化的重組蛋白質細胞株中很有價值[27-28]。
為了構建生產Man9 和Man8 為主要糖鏈結構的高甘露糖型糖鏈的細胞株,我們著眼于定位在高爾基體的甘露糖苷酶Ⅰ。 大多數N-糖基化的蛋白質從內質網轉運至高爾基體時, 其主要N-糖鏈結構為Man9GlcNAc2 或Man8GlcNAc2 的結構。 在高爾基體中,1,2-連接的甘露糖會被高爾基體甘露糖苷酶Ⅰ酶切至Man5GlcNAc2 結構[29-30]。 在哺乳動物細胞中,存在3 種編碼高爾基體甘露糖苷酶Ⅰ的基因,MAN1A1、MAN1A2 和MAN1C1。 我們嘗試通過CRISPR/Cas9 系統在HEK293 細胞中敲除MAN1A1和MAN1A2 基因。 對于每一個基因的敲除,我們在同一個外顯子上設計了兩個目標序列。 當敲除質粒轉染至細胞,單克隆細胞被分離,單克隆細胞在外顯子區域的序列將會被分析,以確定基因是否成功敲除。 成功敲除基因的細胞在設計外顯子區域,兩個目標序列之間的DNA 片段將會被移除, 我們篩選所有染色體在該區域的DNA 片段均被移除的細胞株。 細胞株A1-KO24 和A2-KO37 被認為是MAN1A1-KO 和MAN1A2-KO 的單敲除細胞株。 相較于MAN1A1 基因序列,圖3 展示了對于單敲除細胞株MAN1A1-24 的測序結果,A1-KO24 細胞株在兩個目標序列之間存在一個62 bp 的移除。 Cas9 蛋白質正確地執行了對于前間區序列臨近基序(PAM)上游3 bp 處目標序列的移除,同時切口被非同源末端連接。 同時62 bp 的移除會造成MAN1A1基因內的移碼突變。 同理, 圖4 為單敲除細胞株MAN1A2-KO37 的測序結果,A2-37 細胞株中也存在兩個目標序列間的32 bp 的移除, 同時這個移除也會造成移碼突變。 這些結果顯示MAN1A1 和MAN1A2 成功地被敲除了。
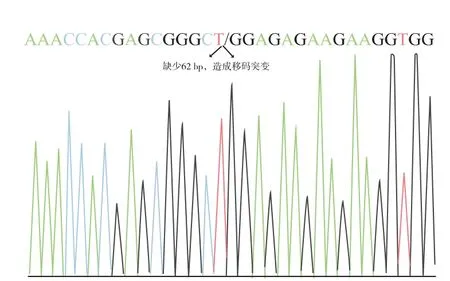
圖3 MAN1A1-KO24 單敲除細胞株的性狀Fig. 3 Genotype of MAN1A1-KO24 cells

圖4 MAN1A2-KO37 單敲除細胞株的性狀Fig. 4 Genotype of MAN1A2-KO37 cells
2.2 構建MAN1A1 和MAN1A2 的雙敲除細胞株
作者構建了MAN1A1 和MAN1A2 雙敲除細胞株(D-KO)。 以A1-KO24 細胞株為親代,在該細胞株上敲除MAN1A2 基因。 在轉染敲除質粒后,細胞被分離。 在敲除目標序列附近的序列被測序分析,用來驗證細胞株MAN1A2 被敲除。 對比A2-KO37細胞株, 雙敲細胞株double-KO 35 (D-KO35)在MAN1A2 目標序列片段被測序后顯示具有3 條不同的結果。 結果如下: 野生型細胞中設計的MAN1A2 靶點之間的序列為aattCCACATGTAGATG CCGGTAAAGGggctaaaaaCCC CGGAGTCTTCCT; 最小的片段是因為兩目標序列之間的DNA 片段被移除后裸露端重新連接而形成的aattCCACATAGTCT TCT,存在32 bp 的移除;中間的片段為在目標靶點1 切除位點處插入了75 bp 長度來自于敲除質粒的序列,75 bp 的序列為來自敲除質粒上的氨芐抗性基因片段的TGGTATGGCTTCATTCAGCTCCGGTTC CCAACGAT CAAGGCGAGTTACATGATCCCCCATG TTGTGCAAA AAAGC,同時在目標靶點2 的切除位點處有一個1 bp 的插入堿基;第3 種片段是由于目標靶點1、2 的切除位點處存在2 bp(GA)和207 bp的插入片段,同樣來自于敲除質粒的207 bp 的序列為AATAAAAATCATTAGGGGATTCATCAGGGCTTTG CGTTGTTTAATATCCTGATCCTGTCCGGGCGTCGG GCGACCTTTGGTCGCCCGGCCTCAGTGAGCGAGC GAGCGCGCAGAGAGGGAGTGGCCAACTCCATCA CTAGGGGTTCCTGCGGCCGCTCCCCAGCATGCCTG CTATTCTCTTCCCAATCCTCCCCCTTGCTGTCCTGC。因為3 種情況均造成移碼突變, 所以可以認為DKO35 為MAN1A1 和MAN1A2 的雙敲除細胞株。
2.3 野生型細胞WT、 雙敲除細胞D-KO35 的生長情況的測定
在獲得了雙敲除細胞后,由于破壞高爾基體α-1,2-甘露糖苷酶Ⅰ需要比較其是否對細胞生長狀態產生影響,同時也要確認獲得的細胞株能否適應生產的需要,需要對細胞的生長情況予以測定。 我們采用CCK-kit[26]對細胞的生長狀態進行測定,見圖5。從圖5 可以看出,細胞增殖速率D-KO 細胞要低于WT 細胞。 說明通過對細胞高爾基體內α1,2-甘露糖苷酶Ⅰ的破壞,會影響細胞增殖。 同時由于敲除α-1,2-甘露糖苷酶Ⅰ的基因對于細胞生長速率產生了影響, 我們也考慮之后通過其他手段,如過表達管家基因等方式實現細胞生長速率的上升。

圖5 野生型細胞和D-KO 細胞的生長曲線Fig. 5 Cell proliferation of WT and D-KO cells
2.4 通過流式分析對雙敲除細胞表面N-糖鏈結構變化進行比較
為了驗證D-KO 細胞株蛋白質上糖鏈結構的變化,我們使用多種凝集素來驗證該變化。 凝集素是來源于植物、無脊椎動物等的一類糖蛋白,具有凝集血紅蛋白的作用。 凝集素具有識別糖蛋白上特定結構的能力,因此在糖生物學研究中往往使用凝集素來驗證蛋白質上糖鏈的結構。 我們使用表1 中的凝集素來與細胞表面蛋白質反應,由于這些凝集素帶有異硫氰酸熒光素(FITC)標記,當凝集素與相應識別的目標糖鏈結構結合時, 帶有FITC 的熒光標記可以幫助我們檢測到熒光變化,從而說明蛋白質上具有可與該凝集素結合的結構。 熒光的變化通過流失細胞分析儀進行檢測。 通過所得到的結果可以計算樣品與凝集素的結合情況,即對應于流失細胞分析儀中的峰的Mean 值。 通過對比野生型WT和D-KO 細胞中Mean 值的相對變化可以得出相對熒光強度, 結果見圖6。 圖6 縱坐標表示以WT 的Mean 值為單位強度的反映D-KO 細胞株的各凝集素相對熒光強度值。 在圖6 中,對比表1 提供的各凝集素對于不同糖鏈結果的特異性識別數據,可以得出, 在D-KO 細胞相較于野生型,ConA-FITC 的相對熒光強度增加, 證明在D-KO 中高甘露糖型N-糖鏈有所增加,為野生型的5.02 倍。 同時,PHAL4-FITC、PHA-E4-FITC、ECA-FITC、DSA-FITC 的數據中,D-KO 細胞均具有明顯的變化,證明細胞表面的蛋白質上糖鏈中Galβ1、4-GlcNAc 和β1,4-GlcNAc 的結構有明顯的減少, 證明復合型N-糖鏈結構在D-KO 細胞株中明顯減少。

表1 流式分析使用的凝集素以及識別糖鏈特異性結構信息Table 1 Lectin use for FACS and its target glycan structure
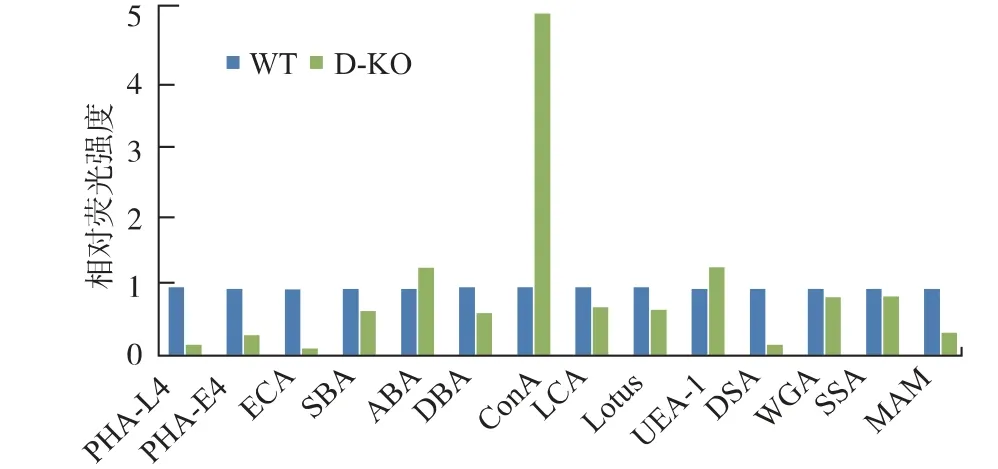
圖6 凝集素染色相對熒光強度Fig.6 Lectin staining and the relative fluorescence intensity
2.5 用western bolt 來驗證雙敲除細胞株內蛋白質上N-糖鏈結構的變化
在構建了MAN1A1 和MAN1A2 的雙敲除細胞株后,我們認為全細胞上的糖蛋白糖鏈的表型結構已經發生了一定程度的改變。 我們希望通過western blot 的手段來驗證糖蛋白上糖鏈類型的改變, 因此我們選擇一種與溶酶體相關的膜蛋白LAMP2 來作為模式蛋白質。 溶酶體相關膜蛋白2(Lysosome-associated membrane protein 2) 也稱作CD107b,LAMP2 蛋白質是一種與溶酶體相關的糖蛋白。將細胞用胰蛋白酶消化后,使用PBS 洗滌。在3 000 r/min 下離心3 min 棄去上清液, 將細胞重懸于細胞裂解液中,冰上放置30 min。 將細胞裂解細胞液的樣品通過SDS-PAGE 并使用anti-LAMP2 的鼠單克隆抗體識別LAMP2 蛋白質。 在圖7 中,野生型和雙敲除細胞D-KO 中的LAMP2 蛋白質大小有所不同,表明其蛋白質上糖基化水平存在不同。 通過EndoH 和PNGaseF 處理后的條帶變化來看,可以證實這一假設。由于PNGaseF 可以移除幾乎所有的糖蛋白上N-糖鏈, 而EndoH 則可以代謝糖蛋白上高甘露糖型N-糖鏈。 在野生型中EndoH 處理后與雙敲除對比發現,PNGaseF 可以移除的糖鏈,EndoH不能很好地移除, 對比雙敲除細胞,LAMP2 對于EndoH 的敏感程度與WT 明顯不同, 進一步表明LAMP2 上糖鏈表型發生了變化,即高甘露糖型糖鏈的比例增加,復合型糖鏈的比例在減少。

圖7 LAMP2 的免疫印跡結果Fig. 7 Western blot result of LAMP2
3 結 語
作者構建了一種基于敲除高爾基體α-1,2-甘露糖苷酶Ⅰ控制基因MAN1A1 和MAN1A2 的破壞來實現糖鏈高甘露糖型N-糖鏈增加的細胞株。 利用CRISPR/Cas9 系 統, 成 功 敲 除MAN1A1 和MAN1A2 基因,獲得雙敲除細胞株。 由于細胞內高甘露糖型糖鏈的增加,使細胞在不同的凝集素染色下,通過流式分析儀分析可以觀察到細胞表面蛋白質糖鏈表型變化。通過對比雙敲除細胞株和野生型細胞株的生長狀態以及胞內溶酶體膜蛋白LAMP2的糖鏈變化,可以基本確定雙敲除細胞株胞內蛋白質糖鏈對于EndoH 的敏感性, 即蛋白質糖鏈為高甘露糖型糖鏈的證明。 通過使高爾基體甘露糖苷酶基因的兩種基因被破壞或敲除,能夠獲得復合型糖鏈的含量大大降低、糖蛋白的穩定性和安全性優異的、 以高甘露糖型糖鏈為主要N-糖鏈結構的糖蛋白。同時作者也提供了一種用于生產以高甘露糖型糖鏈為主要N-糖鏈結構的糖蛋白的動物細胞株、 生產以高甘露糖型糖鏈為主要N-糖鏈結構的糖蛋白的方法、通過該方法制備的糖蛋白以及該糖蛋白的用途。
