淀粉酶對竹黃菌固態發酵產竹紅菌素的影響
2020-05-11 05:08:52高瑞杰鄧華祥李韻雅管政兵廖祥儒蔡宇杰
食品與生物技術學報 2020年2期
關鍵詞:產量
高瑞杰, 鄧華祥, 李韻雅, 管政兵, 廖祥儒, 蔡宇杰*
(1. 江南大學 工業生物技術教育部重點實驗室, 江蘇 無錫214122;2. 江南大學 生物工程學院, 江蘇 無錫214122)
竹黃(Shiraia bambusicola)是中國常見的一種藥用真菌,寄生于特定的竹子的嫩枝上[1],其主要分布在中國的南部省份,國外的日本和斯里蘭卡也有報道[2-3]。
2008 年,第10 版的《真菌字典》將竹黃歸屬到子囊菌門(Ascomycota),座囊菌綱(Dothideomycetes),格孢菌目(Pleosporales)中科地位未確定的竹黃屬[4]。 Cheng 等研究人員通過對竹黃菌的18S rRNA和ITS-5.8S rRNA 的序列進行分析, 提議將竹黃菌歸屬于格孢菌科(Phaeosphaeriaceae)[5]。
作為中國一種傳統的中藥材,竹黃菌的子座中含有多種具有生理活性的物質。 竹紅菌素是竹黃中最主要的紅色色素組分,同時也是竹黃中最重要的生物活性物質,是由多種苝醌類化合物所組成。 目前市場上的竹紅菌素軟膏產品,主要是由昆明振華制藥廠和云南白藥大理制藥廠兩家制藥廠家生產,其有效成分就是竹紅菌素。 竹紅菌素作為一種優良的光敏劑, 在特定的光照條件下能夠產生活性氧,包括超氧自由基和單線態氧。 豐富的活性氧可以引起細胞的高氧化壓力, 損傷包括脂質、DNA 和蛋白質等生物大分子,進而殺死細胞[6-8]。 竹紅菌素光敏活性的研究成為竹紅菌素研究的熱點,國內外多個團隊在這一方面展開了大量的研究工作。 竹紅菌素及其衍生物在抗菌、抗病毒、抗艾滋、抗癌(肝癌、肺癌、乳腺癌等)方面展現出良好的醫療效果[9-14]。
目前市場上的竹紅菌素主要是從竹黃子座中分離獲得,然而天然野生竹黃由于地域分布極不平衡,生長季節短暫,產量低而限制了竹紅菌素的開發利用,因而通過生物發酵法獲取竹紅菌素成為研究熱點。 相對于液態發酵而言,作為絲狀真菌的竹黃菌在進行固態發酵時具有很大的優勢:能量消耗低,生產成本低;無廢水排放,污染小;體積生產率高且不易染菌; 一些應用無需進行提取純化等工藝,例如作為著色劑可直接添加于飼料應用。 因而通過固態發酵的方法來生產竹紅菌素成為本課題的研究重點。
作者所在課題組前期篩選到的竹黃菌Shiraiasp. SUPER-H168[15]能夠穩定高產竹紅菌素,通過對其進行固態發酵方面的初步探究,經過對8 種農作物的篩選后,發現玉米是生產竹紅菌素的最佳固態發酵基質[16]。玉米糝是由玉米的胚乳部分破碎制成,淀粉是玉米胚乳的主要組成成分, 玉米淀粉是由75%的支鏈淀粉和25%的直鏈淀粉組成。 為了提高對底物玉米淀粉的利用率,縮短發酵周期,提高竹紅菌素的產量,作者采用固態發酵的方法,研究了固態基質的初次攪拌、攪拌時間間隔、外加淀粉酶(包括細菌高溫α 淀粉酶、細菌中溫α 淀粉酶、真菌α 淀粉酶和糖化酶)對竹紅菌素產量的影響,以期為商業規模化生產竹紅菌素提供有益的參考及幫助。
1 材料與方法
1.1 菌種
竹黃菌 (Shiraia bambusicola)SUPER-H168:作者所在實驗室保藏[15]。
1.2 培養基
斜面保藏培養基(g/L):馬鈴薯200,葡萄糖20,瓊脂20;pH 自然。
液態種子培養基(g/L):馬鈴薯200,葡萄糖20,KH2PO42,MgSO4·7H2O 0.5;pH 自然。
固態發酵培養基(250 mL 錐形瓶):玉米糝(粒徑0.8~1 mm)25 g, 麥秸稈3 g, 葡萄糖5%,NH4Cl 1% ,CuSO40.05% ,CaCl20.10% ,KH2PO40.05% ,K2HPO40.01%,MgSO40.20%, 初始含水量50%,初始pH 自然[17]。
上述培養基均在121 ℃下濕熱滅菌20 min。
1.3 方法
1.3.1 孢子懸液的制備 斜面種子在30 ℃下培養5~7 d 至長出大量黑色孢子, 用20 mL 無菌水沖洗下黑色孢子,并用玻璃珠將黑色孢子充分打碎并配置成106個/mL 的孢子懸浮液。
1.3.2 種子液的培養 將1.3.1 所述的孢子懸液2 mL 接種于50 mL 的液態種子培養基中, 在30 ℃、200 r/min 的條件下振蕩培養24 h 以制備固態發酵的種子液。
1.3.3 固態發酵的培養 將2 mL 的種子液接種于固態發酵基質中,在30 ℃的條件下培養15 d。 在進行固態基質攪拌優化之前,每兩天對基質進行一次攪拌。
1.3.4 竹紅菌素的測定方法 將固態發酵物進行攪拌后取樣,放置在烘箱中70 ℃烘干直至恒質量,準確稱取1.00 g,用研缽研碎,接著使用30 mL 無水乙醇在80 ℃下浸泡提取兩次,抽濾后得澄清液,用旋轉蒸發儀濃縮蒸干后,用甲醇溶解并且適當稀釋至一定濃度,稀釋后體積為V,然后用分光光度計在465 nm 處測定吸光值, 根據回歸方程y=0.036 4x+0.044 1(R2=0.999)計算色素的含量,其中y為吸光值,x為色素的質量濃度(mg/L)。經換算后得到每克干固態基質的竹紅菌素的產量為x·V,單位為mg/g(每克干固態基質)。
1.3.5 初次攪拌及攪拌間隔時間的優化 首先對
初次攪拌時間進行優化,分別為第1 天、第2 天、第3 天和第4 天,此時隨后的攪拌間隔時間為2 d。 在初次攪拌時間確定后, 進行攪拌間隔時間的優化,分別為12、24、36、48 h。
1.3.6 外加淀粉酶的優化 將固態發酵初始和發酵第3 天作為淀粉酶的加入時間點。 4 種淀粉酶包括細菌高溫α 淀粉酶、細菌中溫α 淀粉酶、真菌α淀粉酶和糖化酶,均購買于江蘇銳陽生物科技有限公司。 4 種淀粉酶分別在固態發酵初始或者第3 天加入,其中高溫α 淀粉酶是在固態基質滅菌之前加入到基質中,于95 ℃保溫30 min。 在對各淀粉酶的添加計量進行優化之前,α 淀粉酶的使用添加量為4 U/g (固態干基質), 糖化酶使用添加量為50 U/g(固態干基質)。細菌中溫α 淀粉酶和真菌α 淀粉酶添加計量的優化:1、2、3、4、5、6 U/g (固態干基質);糖化酶添加計量的優化:10、20、30、40、50、60 U/g(固態干基質)。
α 淀粉酶和糖化酶兩種酶對竹紅菌素產量的協同作用也進行了進一步的探究。 在研究完外加淀粉酶的影響后,發現中溫α 淀粉酶和糖化酶對竹紅菌素的產量的影響顯著。 利用響應面設計進一步探究細菌中溫α 淀粉酶和糖化酶的最佳添加搭配。 采用軟件Design-expert 8.2 進行實驗設計和數據分析,響應面三維圖采用Origin 8.1 進行模擬合成。 得到二次多項式方程后,進而計算出細菌中溫α 淀粉酶和糖化酶的最佳添加濃度。 實驗方案設計中的中心組合設計因素和水平及編碼見表1。
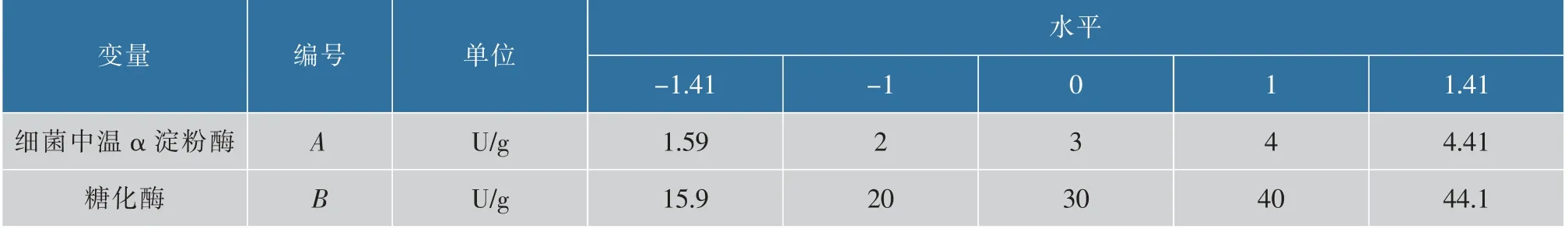
表1 響應面分析因素及水平Table 1 Factors and levels of the variables for response surface methodology experiment
2 結果與分析
2.1 初次攪拌的優化
攪拌是固態耗氧發酵最重要的參數之一,它可以通過固體基質表面液體和氣體形成的分界面的氣體傳輸,以確保溫度和氣體等傳熱和傳質的均一性。 在絲狀真菌進行固態發酵的過程中,攪拌會破壞真菌的菌絲, 尤其是在固態接種發酵后的初期,菌絲在固體基質中正處于延伸定殖并最終覆蓋整個固體基質的階段,過早的攪拌會干擾破壞菌絲的延伸, 不利于菌絲的延伸及固態發酵的順利進行,因而本實驗先對固態發酵的初次攪拌進行了優化。由圖1 可知,當在第3 天進行初始攪拌時,竹紅菌素的產量達到最高水平23.85 mg/g。 隨著初始攪拌從第1 天到第4 天進行的變化,可以觀測到生物量也隨之增加, 尤其是在第4 天進行初始攪拌時,整個固態基質表面附著著大量白色氣生菌絲,且固態基質結塊嚴重,不利于攪拌的進行。
2.2 攪拌間隔時間的優化
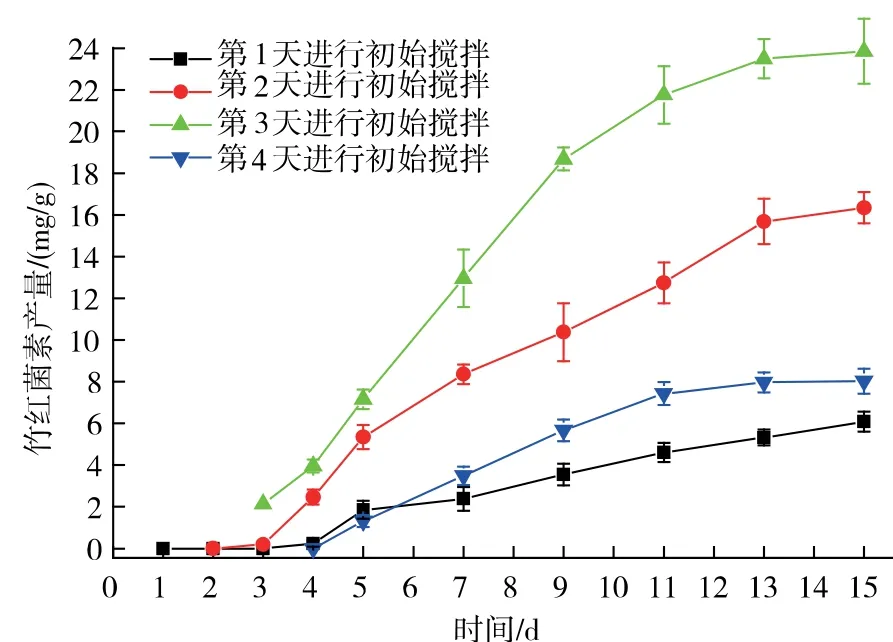
圖1 初始攪拌對竹紅菌素產量的影響Fig. 1 Effect of first agitation time on hypocrellin production
在第3 天確定為最佳初始攪拌時間后,進一步對攪拌間隔時間進行了優化,結果見圖2。當攪拌間隔時間為36 h 時, 竹紅菌素的產量達到最高水平25.37 mg/g, 攪拌間隔時間過短或過長都不利于竹紅菌素的產生。 攪拌間隔時間過短時,攪拌對菌絲的損傷較大以至于影響到竹紅菌素的產量;當攪拌間隔時間過長時,固態基質結塊嚴重,不利于氣體流通和溫度傳遞,也不利于竹紅菌素的產生。
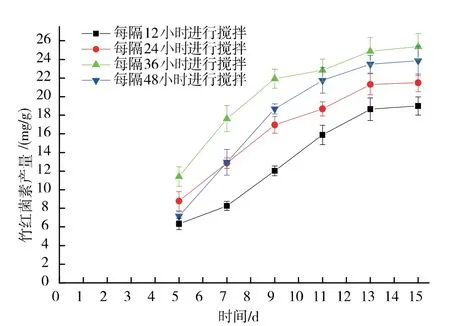
圖2 攪拌間隔時間對竹紅菌素產量的影響Fig. 2 Effect of intermittent agitation time on hypocrellin production
2.3 外加單一淀粉酶的優化
基于攪拌優化的結果,選定固態發酵初始和發酵第3 天作為淀粉酶的加入時間點。 與對照組相比,4 種淀粉酶的7 種加入方式都在不同程度上提高了竹紅菌素的產量,見圖3。中溫α 淀粉酶或真菌α 淀粉酶在發酵初始時加入要比發酵第3 天加入的竹紅菌素產量要高。 其中細菌中溫α 淀粉酶在發酵初始時加入,竹紅菌素產量達到45.10 mg/g。當糖化酶在發酵初始時加入,竹紅菌素產量(32.40 mg/g)要低于在發酵3 d 時加入(39.94 mg/g)。 細菌高溫α淀粉酶的加入處理,固態基質結塊非常厲害且難于進行攪拌處理, 竹紅菌素產量只有29.37 mg/g,因此,細菌高溫α 淀粉酶沒有進行進一步的研究處理。
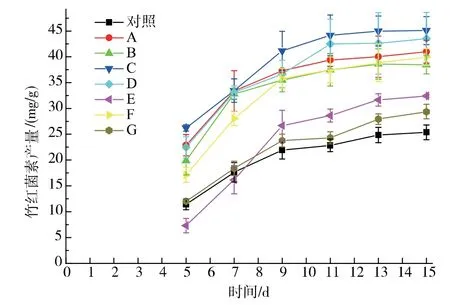
圖3 外加單一淀粉酶對竹紅菌素產量的影響Fig. 3 Effect of different amylase addition on hypocrellin production
2.4 淀粉酶添加量的優化
在選定細菌中溫α 淀粉酶、真菌α 淀粉酶和糖化酶進行進一步研究后,對3 種淀粉酶的使用添加量進行了優化。中溫α 淀粉酶和真菌α 淀粉酶選定在發酵初始時加入,糖化酶選定在發酵3 d 后加入。當細菌中溫α 淀粉酶的使用添加量為3 U/g 時,竹紅菌素產量達到48.12 mg/g; 當真菌α 淀粉酶的使用添加量為4 U/g 時, 竹紅菌素產量達到41.99 mg/g,見圖4。 2 種α 淀粉酶加入處理所造成的竹紅菌素產量的不同, 可能是由于2 種酶的最適pH 和最適溫度的不同造成的,同時說明細菌中溫α 淀粉酶在固態發酵的條件下具有更好的活性,更利于竹黃菌固態發酵產竹紅菌素的進行, 因而選擇細菌中溫α淀粉酶以進一步研究其對竹紅菌素產量的影響。

圖4 α 淀粉酶添加量對竹紅菌素產量的影響Fig. 4 Effect of dose of α -amylase addition on hypocrellin production
當糖化酶的使用添加量為30 U/g 時,竹紅菌素的產量達到40.64 mg/g。 伴隨著糖化酶使用添加量由30 U/g 到60 U/g 的增加, 竹紅菌素的產量逐漸下降,見圖5。 可能是由于酶量增加后,水解產生的大量葡萄糖造成了底物抑制。
2.5 淀粉酶協同加入的優化
兩種α 淀粉酶(細菌中溫α 淀粉酶和真菌α 淀粉酶)和糖化酶的協同作用對竹紅菌素產量影響的探究。 當α 淀粉酶在發酵初始時加入而糖化酶在發酵3 d 后加入時, 竹紅菌素的產量要高于α 淀粉酶和糖化酶二者同時在發酵初始時加入的處理。當中溫α淀粉酶和糖化酶在發酵初始時同時加入,竹紅菌素的產量被抑制到21.01 mg/g,對照組為25.37 mg/g。 當中溫α 淀粉酶在發酵初始時加入而糖化酶在發酵3 d 后加入時,竹紅菌素的產量達到最高,為55.26 mg/g,同時固態發酵周期由15 d 縮減到13 d,見圖6。
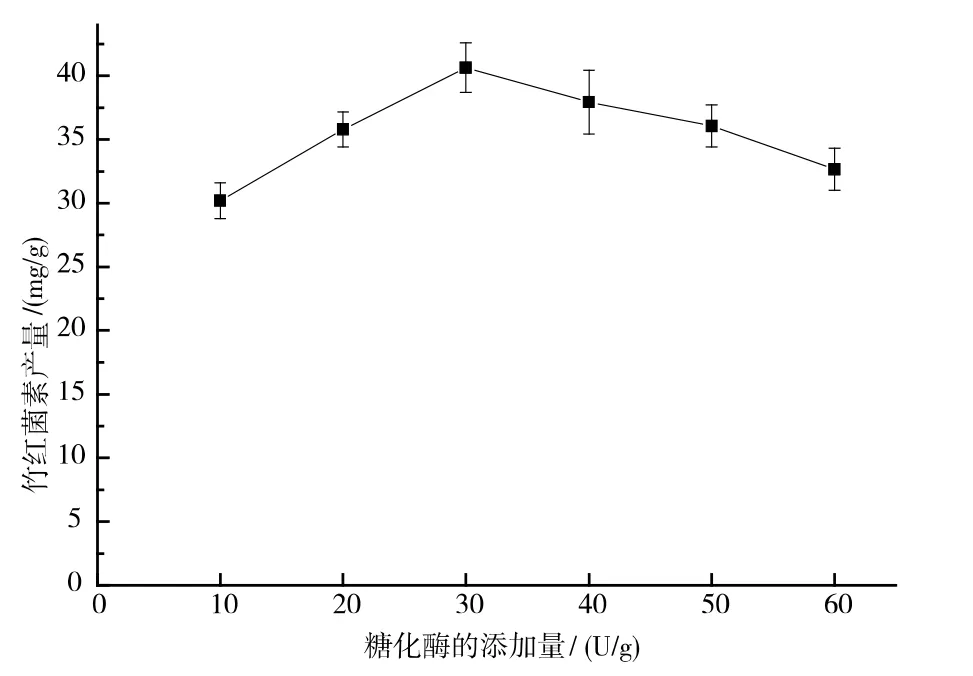
圖5 糖化酶添加量對竹紅菌素產量的影響Fig. 5 Effect of dose of glucoamlase addition on hypocrellin production
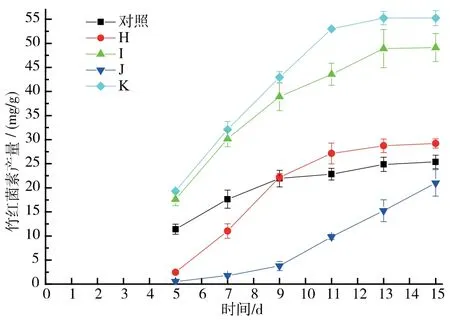
圖6 淀粉酶協同加入對竹紅菌素產量的影響Fig. 6 Effect of synergistic effect of two α-amylases and glucoamylase on hypocrellin production
2.6 細菌中溫α 淀粉酶和糖化酶的響應面實驗
通過上述的實驗研究,發現細菌中溫α 淀粉酶和糖化酶對竹紅菌素的產量影響較大,因而選擇添加這兩種淀粉酶進行響應面研究,結果見表2。
利用軟件Design-Expert 8.0.6 對表2 中的實驗數據進行二次多項回歸擬合,獲得回歸方程為:
Y=68.93-2.19A-0.75B-2.64AB-7.27A2-8.06B2其中Y為響應值(竹紅菌素產量,mg/g);A(細菌中溫α 淀粉酶,U/g)和B(糖化酶,U/g)為響應變量。
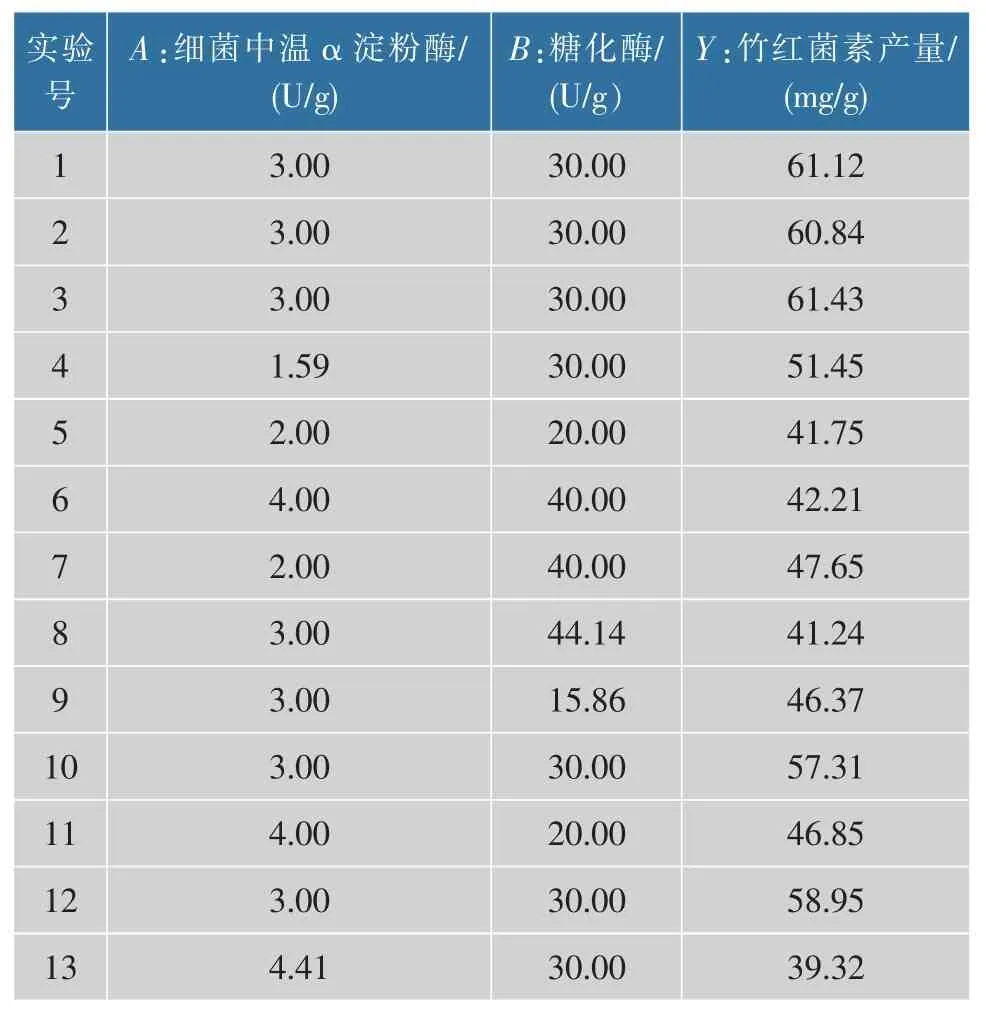
表2 實驗設計方案及實驗結果Table 2 Experimental design matrix and yields of hypocrellin
從回歸方程的系數分析,其所對應的拋物面開口向下, 因而Y具有最大響應值。 利用軟件對表2的數據進行分析, 結果見表3。 該模型的P值為0.000 5,遠小于0.05,說明此模型在95%的概率水平是顯著的;同時模型的失擬項不顯著(p=0.081 7,大于0.05), 說明未知因素對該模型預測的實驗結果的干擾強度比較小, 不必要再引入更多的因素;相關系數R2=0.933 5,說明響應值的變化在93.35%的概率上是由該模型決定的;R=0.966 1, 說明實驗中竹紅菌素的產量的實際值與模型預測值有96.61%的一致性。 因此,該模型能較準確的預測出竹紅菌素的產量。
對回歸方程進行求導,結合圖7 的響應面三維圖,求得該模型的最大響應值為60.10 mg/g,此時對應的中溫α 淀粉酶和糖化酶的添加量分別為2.85、
29.78 U/g。
經過響應面實驗優化,確定固態發酵中細菌中溫α 淀粉酶和糖化酶的添加量分別為2.85、29.78 U/g。 為了驗證預測結果的精準性,在此添加量下進行固態發酵以進行驗證實驗。 在驗證實驗中,竹紅菌素的產量達到60.74 mg/g,與預測值接近,可見該模型能夠較好的反應細菌中溫α 淀粉酶和糖化酶的使用添加量對固態發酵產竹紅菌素的影響。
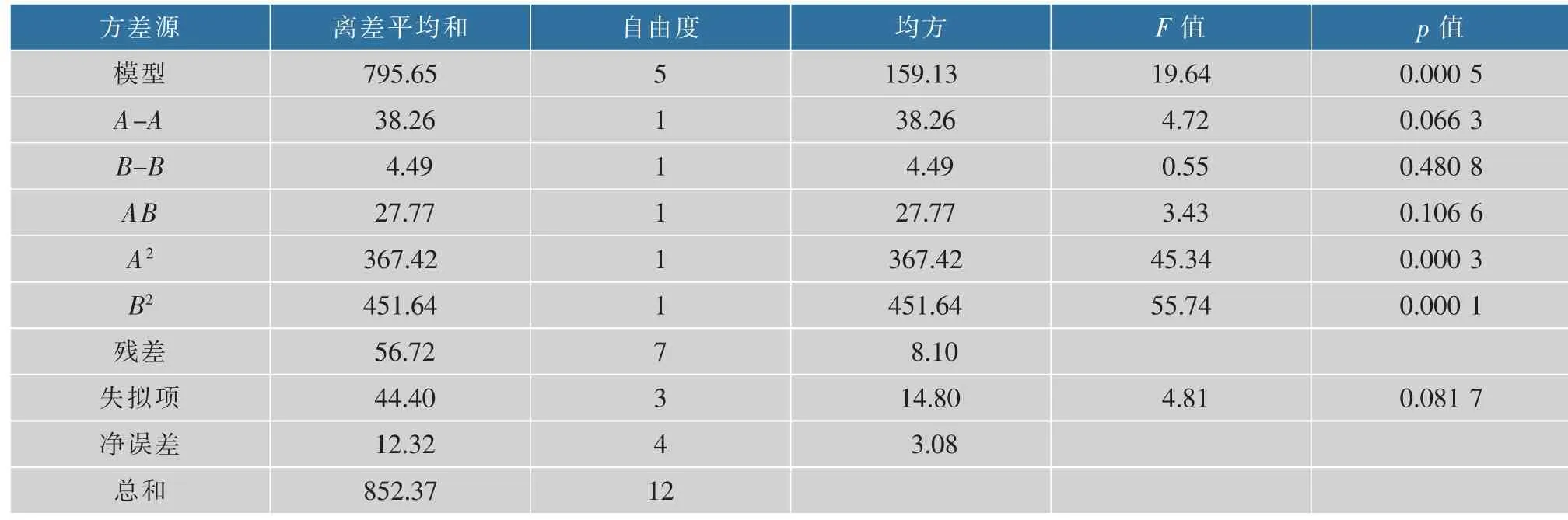
表3 二次多項式回歸模型的方差分析Table 3 Analysis of variance for response surface quadratic model
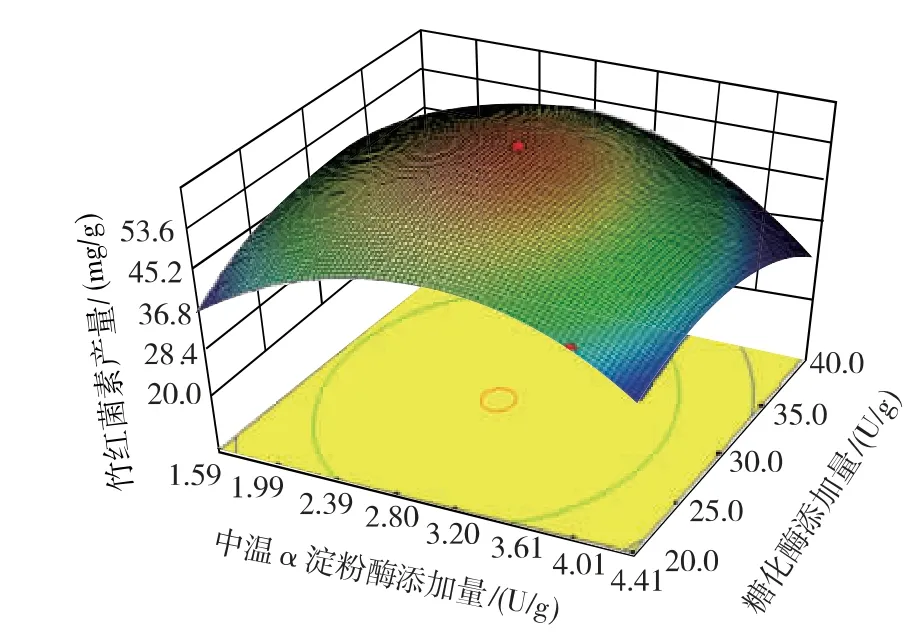
圖7 響應面三維圖Fig. 7 Three dimensional graph of the surface response plot
3 結 語
作者在竹黃菌產竹紅菌素的固態發酵過程中,研究了攪拌(初始攪拌和攪拌間隔時間)和外加淀粉酶對竹紅菌素產量的影響。 確定了固態發酵的最佳攪拌方案和外加淀粉酶的種類及添加量:初始攪拌在發酵第3 天進行, 此后的攪拌間隔時間為36 h, 外加淀粉酶以細菌中溫α 淀粉酶和糖化酶的復合添加為最佳,其中細菌中溫α 淀粉酶在固態發酵初始階段加入,添加量為2.85 U/g,糖化酶在發酵第3 天加入,添加量為29.78 U/g。 經過攪拌和外加淀粉酶的研究后,竹紅菌素的產量到達了60.74 mg/g,是先前竹紅菌素產量[17]報道的3.66 倍,并且固態發酵的周期由15 d 縮減到了13 d。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
當代水產(2021年10期)2021-12-05 16:31:48
今日農業(2021年14期)2021-11-25 23:57:29
今日農業(2021年13期)2021-08-14 01:37:56
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
中國果業信息(2019年10期)2019-11-13 01:21:34
中國化肥信息(2019年2期)2019-01-18 15:24:35
中國化肥信息(2019年1期)2019-01-17 21:31:12
中國化肥信息(2019年4期)2019-01-17 18:47:06
