3個羅非魚種群對4種病原菌的抗病力差異比較
2020-05-11 06:06:33敖秋桅朱佳杰
南方農業學報 2020年3期
敖秋桅 朱佳杰
摘要:【目的】比較吉富羅非魚F5代(以下簡稱吉富F5代)、吉富羅非魚F0代(以下簡稱吉富F0代)及奧尼羅非魚對4種病原菌的抗病力差異,為羅非魚苗種推廣養殖提供參考依據。【方法】檢測吉富F5代無乳鏈球菌(Ia和Ib血清型)、海豚鏈球菌和嗜水氣單胞菌對吉富F5代的半致死濃度(LC50),并對3個羅非魚種群進行人工腹腔注射感染4種病原菌,根據感染后的累計死亡率評估其抗病力差異。【結果】無乳鏈球菌Ia血清型對吉富F5代的LC50為2.14×105 CFU/mL,無鏈球菌Ib血清型對吉富F5代的LC50為2.14×106 CFU/mL,海豚鏈球菌對吉富F5代的LC50為2.14×107 CFU/mL,嗜水氣單胞菌對吉富F5代的LC50為5.62×107 CFU/mL。3個羅非魚種群感染病原菌后連續觀察168 h發現,感染無乳鏈球菌Ia血清型菌株(HN016)的累計死亡率排序為吉富F0代>吉富F5代>奧尼羅非魚;感染無乳鏈球菌Ib血清型菌株(GX26)的累計死亡率排序為吉富F0代>吉富F5代>奧尼羅非魚;感染海豚鏈球菌菌株(GX05)的累計死亡率排序為吉富F0代>奧尼羅非魚>吉富F5代;感染嗜水氣單胞菌菌株(GX03)的累計死亡率排序為吉富F0代>吉富F5代>奧尼羅非魚。方差分析結果顯示,吉富F5代感染HN016、GX26、GX05和GX03后的累計死亡率均顯著低于吉富F0代(P<0.05),與奧尼羅非魚差異不顯著(P>0.05)。【結論】經過連續5代選育后,吉富羅非魚對無乳鏈球菌(Ia和Ib血清型)、海豚鏈球菌及嗜水氣單胞菌的抗感染能力均得到顯著提高,并已接近奧尼羅非魚的抗病水平。
關鍵詞: 羅非魚;抗病性;無乳鏈球菌;海豚鏈球菌;嗜水氣單胞菌
0 引言
【研究意義】我國自20世紀90年代后期引進吉富羅非魚并大面積推廣養殖后,羅非魚產業得到飛速發展,并逐步發展成為全球最大的羅非魚養殖及出口國家(陳勝軍等,2007;袁永明等,2013)。但近年來,由于養殖模式落后、水體環境惡化、種質退化及大規模疾病頻發,養殖戶承受了巨大經濟損失,已嚴重制約羅非魚產業的健康發展(王世表等,2016)。在羅非魚的眾多疾病中,以細菌性疾病的危害最嚴重,代表性病原菌有無乳鏈球菌、海豚鏈球菌和嗜水氣單胞菌等,當前尚無有效的防治手段(孫永軍,2014)。此外,養殖戶為預防疾病暴發而濫用抗生素,導致商品魚藥物殘留超標,水體環境嚴重污染,尤其是耐藥菌株的產生導致病害頻繁發生。已有研究表明,從種質上提高羅非魚的天然抗病力以抵御病原菌入侵,是上解決病害防治及食品安全問題最有效的方法(紀榮興,2008;朱佳杰,2019)。因此,研究羅非魚對不同病原菌的抵抗力,對綜合評價其抗病力具有重要意義。【前人研究進展】在水產生物抗病育種上,日本科學家成功獲得虹鱒的抗病品種,并發掘了相關抗傳染性胰腺壞死病毒(IPNV)的QTL標記(Ozaki et al.,2001);挪威科學家對大西洋鮭進行家系育種及分子標記輔助抗病育種,建立了一批抗IPNV家系和易感病家系(Storset et al.,2007)。近年來,我國水產育種技術突飛猛進,大量學者圍繞水產動物的高產、抗逆、抗病等性狀開展品種選育研究工作。陳松林等(2008)通過人工感染牙鲆篩選、構建抗病力強家系,同時結合生長數據,獲得了抗鰻弧菌病、生長快的新品種鲆優1號;此外,又以養殖群體和渤海野生群體的半滑舌鰨為基礎群,通過家系構建和人工感染鰻弧菌方式,篩選獲得了一批抗病力強、生長速度快的家系(陳松林等,2010)。孔杰等(2012)開展中國對蝦生長和抗病的多性狀復合選育研究,獲得新品種黃海2號,其生長速度和抗病能力均得到明顯提升。鄭衛衛(2016)使用家系選育法構建一批抗鰻弧菌牙鲆家系,并通過育種值估計,發現抗病性能在親子代得到較好遺傳。白姍姍等(2017)開展鏡鯉抗鯉皰疹病毒(CyHV-3)新品種選育工作,發現其F3代的抗病力已得到穩定提升。奧尼羅非魚的抗病能力顯著優于吉富羅非魚,但存在生長速度偏慢、產苗率低、群體均勻度差等問題,影響養殖推廣;吉富羅非魚因具有生長速度快、產量高等優勢已成為目前羅非魚主養品種,但其抗病能力弱,急需改良(莫振明和林東年,2010;朱佳杰等,2015;祝璟琳等,2017)。朱佳杰(2019)自2012年起開展羅非魚抗無乳鏈球菌病新品種選育工作,至今已選育5個世代,并于2018年通過全國水產原良種委員會審定,命名為壯羅1號羅非魚,其養殖成活率比普通吉富羅非魚提高15.2%~21.4%,生長速度較普通吉富羅非魚提高9.1%~15.1%。【本研究切入點】羅非魚品種繁多,種質資源較豐富,不同品種間的天然抗病力存在差異。在水產品種抗病選育中,通常采用特定單一的病原菌對育種對象進行人工感染,從中挑選出耐受力強的個體或家系。而羅非魚病害監測結果顯示,羅非魚病害已從單一病原發病逐步過渡至多病原發病(國家羅非魚產業技術體系,2016),但目前關于羅非魚對多種病原菌的抗病力研究鮮見報道。【擬解決的關鍵問題】以吉富羅非魚F5代(以下簡稱吉富F5代)、吉富羅非魚F0代(以下簡稱吉富F0代)及奧尼羅非魚為對象,分別采用自然環境下的常見病原菌無乳鏈球菌(Ia和Ib血清型)、海豚鏈球菌及嗜水氣單胞菌進行注射感染,分析3個羅非魚種群對4種病原菌的抗病力差異,為壯羅1號羅非魚的推廣及后續新品種選育提供科學依據。
1 材料與方法
1. 1 試驗材料
試驗用吉富羅非魚為2003年從國家級青島羅非魚良種場引進,2012年以抗無乳鏈球菌病感染能力和生長速度為目標進行選育,現已選育至F5代(吉富F5代),同時每年采用群體繼代選育方法進行保種擴繁(吉富F0代)。奧尼羅非魚由國家級廣西南寧羅非魚良種場繁育提供。挑選健康有活力、規格統一(平均重56.48±0.70 g)的3個種群羅非魚各500尾,試驗前用雞血平板分離檢測各種群羅非魚是否攜帶無乳鏈球菌等病原。常規暫養8~10 d,提供充足氧氣及保持水質清新。試驗菌株分別為無乳鏈球菌Ia血清型菌株(HN016)、無乳鏈球菌Ib血清型菌株(GX26)、海豚鏈球菌菌株(GX05)和嗜水氣單胞菌菌株(GX03),均由廣西水產科學研究院魚病室保存提供。
1. 2 半致死濃度(LC50)測定
使用血平板菌落計數法計算菌液濃度,并以10倍梯度對病原菌懸液進行稀釋。各菌株的系列梯度濃度為:5.0×108、5.0×107、5.0×106、5.0×105、5.0×104和5.0×103 CFU/mL。挑選健康無外傷、規格統一的吉富F5代200尾,每個梯度濃度菌懸液腹腔注射10尾羅非魚,注射量為0.2 mL/尾,對照組注射等體積無菌生理鹽水,試驗組設2次重復。每組單獨飼養在容量為1 m3的大白桶中,保持水溫30~31 ℃。定時觀察魚的發病和死亡狀況,持續觀察168 h。LC50參照Reed-Muench法進行計算。
1. 3 不同品系羅非魚感染病原菌結果對比
選取規格無明顯差異的吉富F5代、吉富F0代和奧尼羅非魚,通過腹腔注射分別感染HN016(1.07×106 CFU/mL)、GX26(1.07×107 CFU/mL)、GX05(1.07×108 CFU/mL)和GX03(2.81×108 CFU/mL),菌株感染濃度根據上一步試驗所得的LC50計算。每30尾為1組,每組設2次重復,每尾注射量0.2 mL,總計感染720尾魚。3個羅非魚種群均設30尾的對照組,注射等體積無菌生理鹽水。注射感染后定時觀察試驗魚死亡狀況,持續觀察168 h,及時清理死魚,保持水質良好,水溫控制在30~31 ℃。
1. 4 統計分析
試驗數據使用SPSS 18.0進行統計及單因素分析(One-way ANOVA)。
2 結果與分析
2. 1 不同病原菌的LC50測定結果
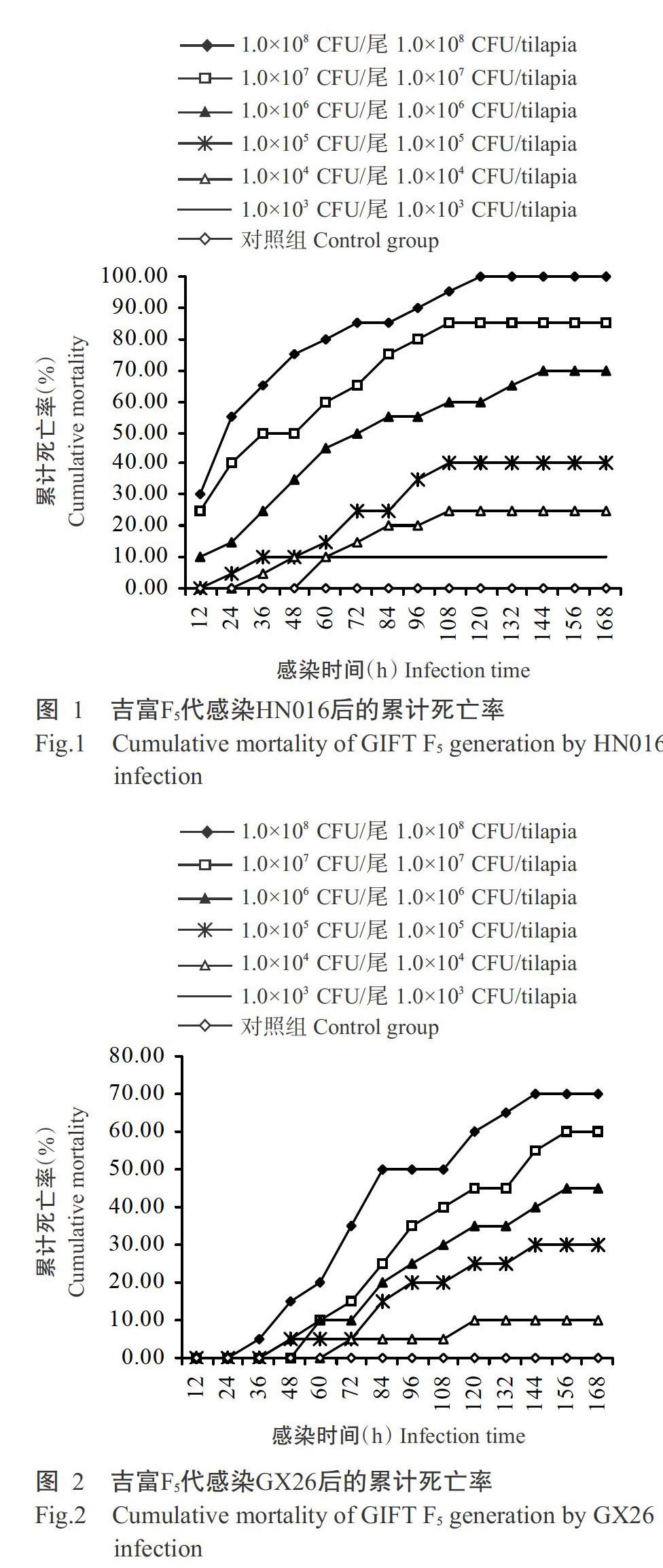
吉富F5代感染不同濃度HN016后,病魚眼睛和鰓蓋充血,魚體失去平衡轉圈亂游,解剖后發現魚體肝胰腺、腎臟和脾臟明顯腫大,腸壁變薄。從圖1可看出,吉富F5代感染不同濃度HN016后12 h開始出現死亡,而相應的對照組無死亡現象發生;感染后24~84 h為死亡高峰期,感染84 h后死亡魚數逐漸減少,感染168 h后基本停止死亡;大于或等于1.0×106 CFU/mL的3個濃度感染吉富F5代的累計死亡率均超過50.00%。經Reed-Muench公式計算得知,HN016對吉富F5代的LC50為2.14×105 CFU/mL。
從圖2可看出,吉富F5代感染不同濃度GX26后36 h開始出現死亡,發病狀況與感染HN016相似,相應的對照組也未見死亡發生;感染后48~108 h為死亡高峰期,感染120 h后死亡魚數逐漸減少;濃度大于或等于1.0×107 CFU/mL的2個濃度感染吉富F5代的累計死亡率均超過50.00%,低濃度(≤1.0×103 CFU/尾)GX26感染未出現死亡。經Reed-Muench公式計算得知,GX26對吉富F5代的LC50為2.14×106 CFU/mL。
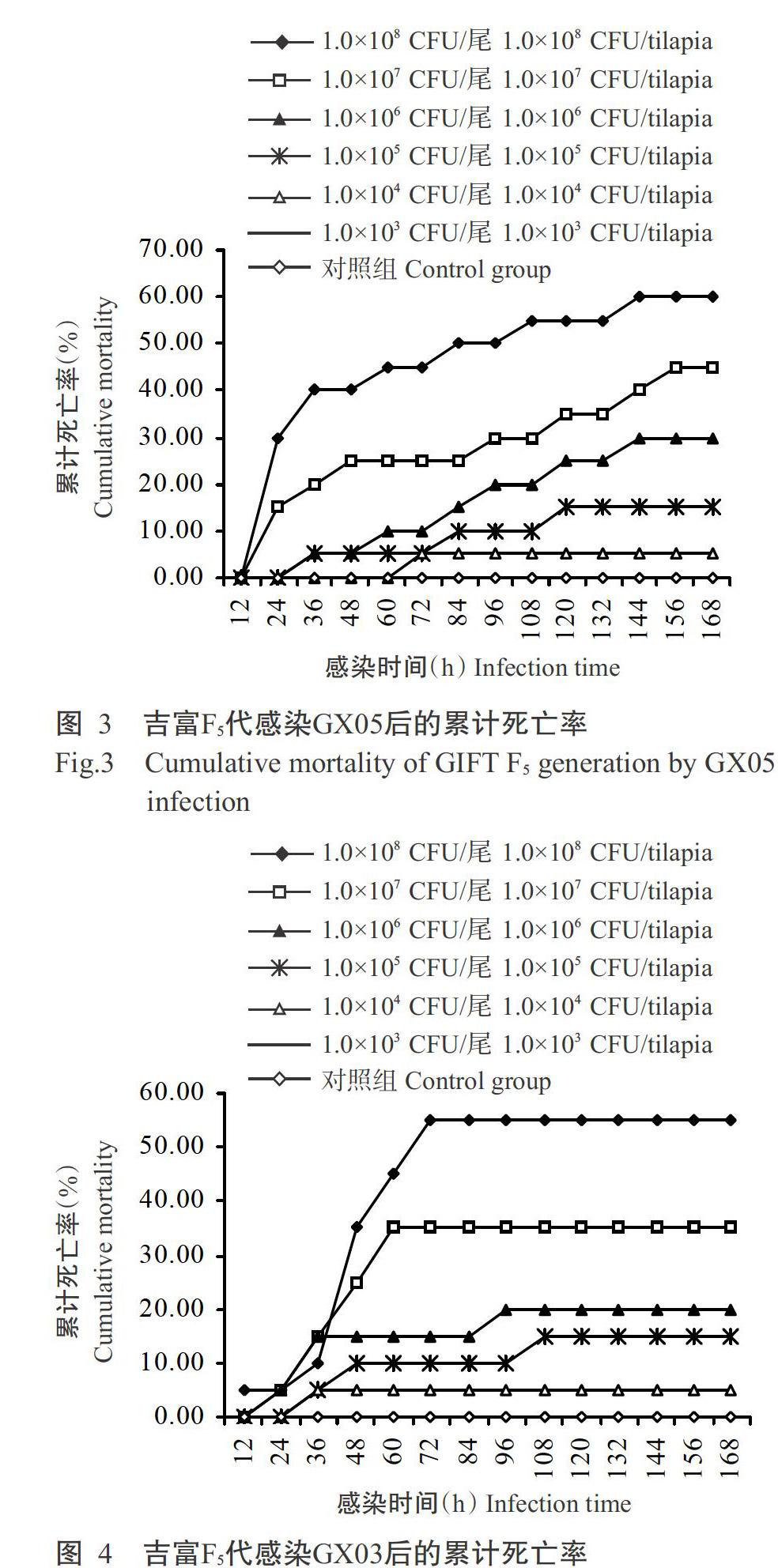
從圖3可看出,吉富F5代感染不同濃度GX05后24 h開始出現死亡,發病狀況與感染HN016相似,相應的對照組也未出現死亡;感染后24~72 h為死亡高峰期,感染96 h后死亡魚數逐漸減少;高濃度(≥1.0×108 CFU/尾)GX05感染吉富F5代的累計死亡率為60.00%,低濃度(≤1.0×103 CFU/尾)GX05感染未出現死亡。經Reed-Muench公式計算得知,GX05對吉富F5代的LC50為2.14×107 CFU/mL。
吉富F5代感染不同濃度GX03后,病魚魚體發黑脫鱗,皮膚潰爛,解剖發現魚體內肝胰腺和腎臟腫大,感染后12 h開始出現死亡,而相應的對照組未出現死亡。從圖4可看出,吉富F5代感染GX03后24~60 h為死亡高峰期,感染72 h后死亡魚數基本穩定;高濃度(≥1.0×108 CFU/尾)GX03感染吉富F5代的累計死亡率超過50.00%,低濃度(≤1.0×103 CFU/尾)GX03感染未出現死亡。經Reed-Muench公式計算得知,GX03對吉富F5代的LC50為5.62×107 CFU/mL。
2. 2 4種病原菌感染3個羅非魚種群后的死亡率比較
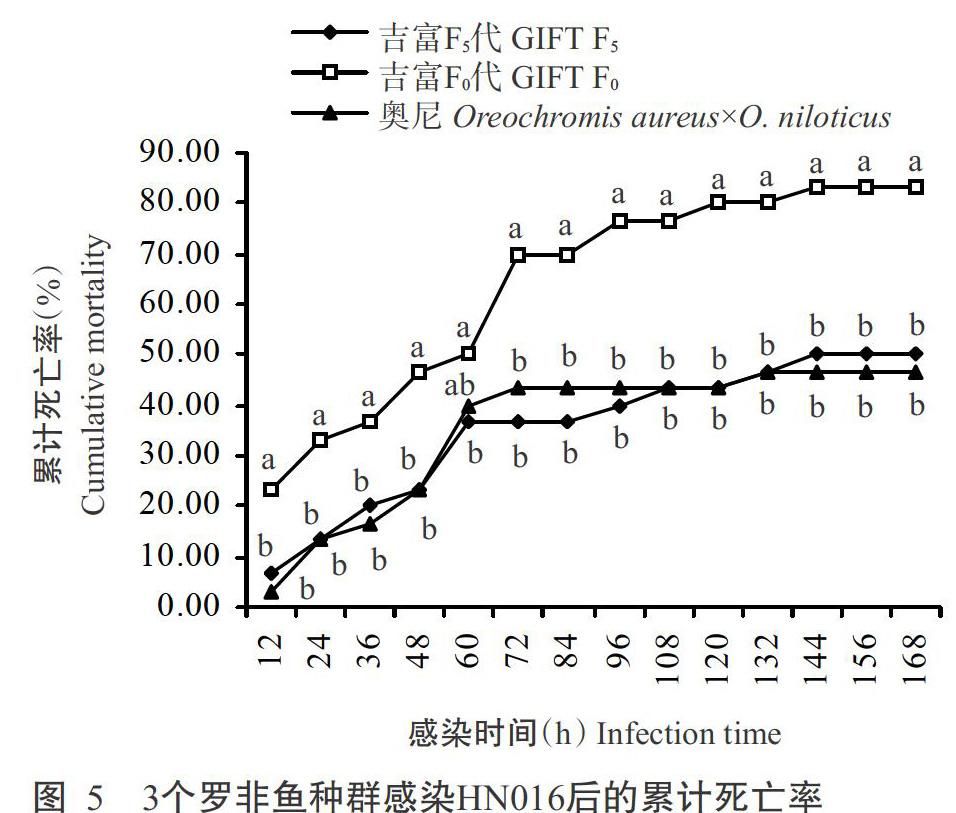
從圖5可看出,3個羅非魚種群注射感染HN016后12 h均開始出現死亡,死亡多集中在感染后96 d內,而各相應的對照組均未出現死亡。對發病魚解剖觀察發現,其病理特征均符合無乳鏈球菌的典型發病癥狀;對血瓊脂培養基上接種病魚腦組織和肝臟組織的優勢菌落進行革蘭氏染色鏡檢,也證實病魚感染的病菌為無乳鏈球菌。感染HN016后168 h,3個羅非魚種群的累計死亡率排序為吉富F0代(83.88%)>吉富F5代(50.00%)>奧尼羅非魚(46.67%)。方差分析結果表明,感染HN016后吉富F5代的累計死亡率顯著低于吉富F0代(P<0.05,下同),總體上略高于奧尼羅非魚,但差異不顯著(P>0.05,下同)。說明吉富F5代抗HN016感染能力已接近奧尼羅非魚的水平。
從圖6可看出,3個羅非魚種群注射感染GX26后36 h開始出現死亡,死亡多集中在感染后120 h內。對病魚腦組織進行取樣培養及革蘭氏染色鏡檢后確定為感染無乳鏈球菌。感染GX26后168 h,3個羅非魚種群的累計死亡率排序為吉富F0代(70.00%)>吉富F5代(46.67%)>奧尼羅非魚(43.33%)。方差分析結果表明,感染GX26后吉富F0代的累計死亡率顯著高于吉富F5代和奧尼羅非魚,但吉富F5代的死亡率與奧尼羅非魚差異不顯著。綜上所述,經選育后吉富F5代對無乳鏈球菌(Ia和Ib血清型)的抗感染能力顯著優于吉富F0代,已接近奧尼羅非魚的抗病水平。
從圖7可看出,3個羅非魚種群感染GX05后24 h開始出現死亡,死亡多集中在感染后96 h內。對病魚腦組織進行取樣培養及革蘭氏染色鏡檢確定為感染海豚鏈球菌。感染168 h后,3個羅非魚種群的累計死亡率排序為吉富F0代(56.67%)>奧尼羅非魚(43.33%)>吉富F5代(40.00%)。方差分析結果表明,感染GX05后24 h,3個羅非魚種群的死亡率表現為吉富F0代>吉富F5代>奧尼羅非魚,且相互間差異顯著,但感染后168 h,3個種群羅非魚的死亡率表現為吉富F0代>奧尼羅非魚>吉富F5代,吉富F0代的累計死亡率顯著高于奧尼羅非魚和吉富F5代,而奧尼羅非魚和吉富F5代差異不顯著。可見,吉富F5代與奧尼羅非魚對海豚鏈球菌的抗感染能力相近。
從圖8可看出,3個羅非魚種群感染GX03后12 h開始出現死亡,死亡多集中在感染后84 h內。對病魚腦組織進行取樣培養及革蘭氏染色鏡檢確定為感染嗜水氣單胞菌。感染GX03后168 h,3個羅非魚種群的累計死亡率排序為吉富F0代(70.00%)>吉富F5代(50.00%)>奧尼羅非魚(40.00%)。方差分析結果表明,感染GX03后168 h,吉富F0代的累計死亡率顯著高于吉富F5代和奧尼羅非魚,但吉富F5代的累計死亡率與奧尼羅非魚差異不顯著。可見,吉富F5代抗嗜水氣單胞菌感染的能力與奧尼羅非魚相近。
3 討論
已有研究表明,我國南方羅非魚主產區的羅非魚病害逐年加劇,主要表現為以細菌性疾病為主引起大規模發病及死亡,其中鏈球菌病發生最嚴重,其次是嗜水氣單胞菌(國家羅非魚產業技術體系,2016;蘇友祿等,2019)。吉富羅非魚是我國羅非魚養殖的主導品種,生長速度快,但抗病力弱,尤其對鏈球菌敏感,極易感染發病。本研究發現,HN016對3個羅非魚種群的毒力最強,GX26次之,GX05和GX03的毒力較弱,而HN016為本課題組前期開展吉富羅非魚抗病選育所選用的毒株,屬于Ia血清型(沈夏霜等,2018)。Ia血清型為羅非魚源無乳鏈球菌的優勢血清型,流行范圍廣,時間長,而Ib血清型無乳鏈球菌主要在廣東、廣西及海南小范圍流行(袁偉,2017)。本研究還發現,吉富羅非魚感染HN016后發病快,病死率高,而感染GX26后發病較晚,病死率較低,說明吉富羅非魚對不同血清型菌株的敏感性存在差異。強俊等(2012)采用2.95×10 8 CFU/mL海豚鏈球菌感染吉富和新吉富等羅非魚品系,感染后12 h出現死亡,吉富和新吉富羅非魚累計死亡率分別為36.67%和38.33%,與本研究羅非魚的發病死亡時間相近,但本研究高濃度(1.0×108 CFU/尾)GX05感染后的死亡率達60.00%,與強俊等(2012)的研究結果差異明顯,可能與吉富羅非魚種群的種質差異有關,也可能與致病菌株的毒力相關。本研究中,吉富羅非魚感染GX03后12 h已有死魚出現,且死亡率急劇上升,說明吉富羅非魚對該菌株非常易感,但感染108 h后死亡魚數逐漸趨于穩定,發病魚體表癥呈發黑脫鱗、解剖后見肝臟腫大壞死,發病過程及癥狀與柯劍(2011)、楊寧等(2014)研究發現嗜水氣單胞菌感染尼羅羅非魚的癥狀相似,說明本研究開展嗜水氣單胞菌感染吉富羅非魚試驗結果可靠。
在感染死亡率方面,本研究發現吉富羅非魚感染HN016和GX26后,其F0代的累計死亡率顯著高于奧尼羅非魚和吉富F5代,吉富F5代的累計死亡率略高于奧尼羅非魚,與柯劍(2011)、Huang等(2013)、祝璟琳等(2017)的研究結果一致;3個羅非魚種群感染GX05后,吉富F0代的累計死亡率最高,吉富F5代的累計死亡率最低,與強俊等(2012)研究發現感染海豚鏈球菌后吉富及新吉富羅非魚的累計死亡率顯著高于尼羅羅非魚的研究結果相似。Huang等(2013)、祝璟琳等(2017)研究表明,奧尼羅非魚由于是尼羅羅非魚和奧利亞羅非魚遠緣雜交而得,具有明顯雜種優勢,在抗病、抗逆及雄性率等方面均顯著優于吉富羅非魚。吉富羅非魚是由不同地理種群的尼羅羅非魚種間雜交選育而成,由于持續對生長性狀進行選擇,其抗病、抗逆等性能已衰減。但本研究發現,吉富F5代感染HN016、GX26、GX05和GX03的累計死亡率差異均顯著低于吉富F0代,且與奧尼羅非魚的死亡率相近,說明經過5個世代選育后,與調控羅非魚抗病性狀相關的優勢等位基因得到了很好的富集,其抗病性能已接近奧尼羅非魚的抗病水平,也進一步驗證了沈夏霜等(2018)對吉富F5代抗病和生長性能的評估結論。
4 結論
經過連續5代選育后,吉富羅非魚對無乳鏈球菌(Ia和Ib血清型)、海豚鏈球菌和嗜水氣單胞菌的抗感染能力均得到顯著提升,其抗病能力已接近奧尼羅非魚的水平。
參考文獻:
白姍姍,賈智英,郎延賀,石連玉. 2017. CyHV-3感染鏡鯉選育世代免疫基因表達及抗病能力比較[J]. 上海海洋大學學報,26(5):651-657. [Bai S S,Jia Z Y,Lang Y H,Shi L Y. 2017. Comparisons of the immune gene expression and disease resistance between mirror carp breeding generations after CyHV-3 infection[J]. Journal of Shanghai Ocean University,26(5):651-657.]
陳勝軍,李來好,楊賢慶,岑劍偉,吳燕燕,刁石強. 2007. 我國羅非魚產業現狀分析及提高羅非魚出口競爭力的措施[J]. 南方水產,3(1):75-80. [Chen S J,Li L H,Yang X Q,Cen J W,Wu Y Y,Diao S Q. 2007. Present situation of tilapia industry and measures of improving export competition power[J]. South China Fisheries Science,3(1):75-80.]
陳松林,杜民,楊景峰,胡喬木,徐營,翟介明. 2010. 半滑舌鰨家系建立及其生長和抗病性能測定[J]. 水產學報,34(12):1789-1794. [Chen S L,Du M,Yang J F,Hu Q M,Xu Y,Zhai J M. 2010. Development and characterization for growth rate and disease resistance of families in half-smooth tongue sole(Cynoglossus semilaevis)[J]. Journal of Fisheries of China,34(12):1789-1794.]
陳松林,田永勝,徐田軍,鄧寒,劉壽堂,劉本偉,季相山,于過才. 2008. 牙鲆抗病群體和家系的建立及其生長和抗病性能初步測定[J]. 水產學報,32(5):665-673. [Chen S L,Tian Y S,Xu T J,Deng H,Liu S T,Liu B W,Ji X S,Yu G C. 2008. Development and characterization for growth rate and disease resistance of disease-resistance population and family in Japanese flounder(Paralichthys olivaceus)[J]. Journal of Fisheries of China,32(5):665-673.]
國家羅非魚產業技術體系. 2016. 中國現代農業產業可持續發展戰略研究羅非魚分冊[M]. 北京:中國農業出版社. [National tilapia industry technology system. 2016. Research on sustainable development strategy of modern agricultural industry in China[M]. Beijing:China Agriculture Press.]
紀榮興,鄒文政,鄢慶枇,龔三妹,葉兆弘. 2008. 日本對蝦抗白斑病子三代的抗白斑綜合征病毒感染能力及免疫特性[J]. 水產學報,32(1):98-104. [Ji R X,Zou W Z,Yan Q P,Gong S M,Ye Z H. 2008. Immunogenicity and WSSV resistance of anti-WSSV Penaeus japonicus F3[J]. Journal of Fisheries of China,32(1):98-104.]
柯劍. 2011. 羅非魚病原菌分離鑒定及不同品系間抗病力初步比較[D]. 上海:上海海洋大學. [Ke J. 2011. Bacteria pathogeny isolation,identification and primary comparative study of disease resistance in tilapia species[D]. Shanghai:Shanghai Ocean University.]
孔杰,羅坤,欒生,王清印,張慶文,張天時,孟憲紅,王偉繼,阮曉紅. 2012. 中國對蝦新品種“黃海2號”的培育[J]. 水產學報,36(12):1854-1862. [Kong J,Luo K,Luan S,Wang Q Y,Zhang Q W,Zhang T S,Meng X H,Wang W J,Ruan X H. 2012. The new variety of Fenneropenaeus chinensis “Huanghai No.2”[J]. Journal of Fisheries of China,36(12):1854-1862.]
莫振明,林東年. 2010. 吉富羅非魚與奧尼羅非魚養殖對比試驗報告[J]. 現代漁業信息,25(8):24-27. [Mo Z M,Lin D N. 2010. Report on comparative experiment between geneticcally improved farmed tilapia and Oreochromis aurea[J]. Modern Fisheries Information,25(8):24-27.]
強俊,楊弘,王輝,徐跑,柒壯林,何杰. 2012. 海豚鏈球菌感染對不同品系羅非魚血液生化指標和肝臟HSP70 mRNA表達的影響[J]. 水產學報,36(6):958-968. [Qiang J,Yang H,Wang H,Xu P,Qi Z L,He J. 2012. Studies on blood biochemical indices and expression of hepatic HSP70 mRNA of different tilapia strains artificially cha-llenged with Streptococcus iniae[J]. Journal of Fisheries of China,36(6):958-968.]
沈夏霜,敖秋桅,甘西,譚蕓,羅永巨,梁軍能,朱佳杰. 2018. 吉富羅非魚抗病品系F5代抗病性能和生長性能的評估[J]. 南方水產科學,14(3):83-90. [Shen X S,Ao Q W,Gan X,Tan Y,Luo Y J,Liang J N,Zhu J J. 2018. Estimation of disease resistance and growth in F5 generation families of GIFT tilapia[J]. South China Fisheries Scien-ce,14(3):83-90.]
蘇友祿,劉嬋,鄧益琴,郭志勛,馮娟. 2019. 羅非魚無乳鏈球菌病的研究進展[J]. 大連海洋大學學報,34(5):757-766. [Su Y L,Liu C,Deng Y Q,Guo Z X,Feng J. 2019. Research on Streptococcus agalactiae disease in tilapia:A review[J]. Journal of Dalian Ocean University,34(5):757-766.]
孫永軍. 2014. 廣西羅非魚產業發展研究[D]. 南京:南京農業大學. [Sun Y J. 2014. The study of tilapia industry development in Guangxi[D]. Nanjing:Nanjing Agriculture University.]
王世表,宋懌,黃磊. 2016. 中國羅非魚產業現狀、存在問題和發展對策[J]. 中國漁業質量與標準,6(5):27-31. [Wang S B,Song Y,Huang L. 2016. The status,problems and development countermeasures of Chinese tilapia industry[J]. Chinese Fishery Quality and Standards,6(5):27-31.]
楊寧,黃海,張希,姜芳燕. 2014. 尼羅羅非魚嗜水氣單胞菌病的病原分離鑒定和藥敏試驗[J]. 水產科學,33(5):306-310.[Yang N,Huang H,Zhang X,Jiang F Y. 2014. Isolation,identification and drug sensitive test of pathogen Aeromonas hydrophila from Nile tilapia Oreochromis nilotica[J]. Fisheries Science,33(5):306-310.]
袁偉. 2017. 中國羅非魚源無乳鏈球菌流行特征及耐藥性研究[D]. 上海:上海海洋大學. [Yuan W. 2017. Epidemiological characteristics and drug resistance of Streptococcus agalactiae isolated from tilapia in China[D]. Shanghai:Shanghai Ocean University.]
袁永明,袁媛,賀艷輝,代云云,龔赟翀,張紅燕,王紅衛. 2013. 我國羅非魚產業發展趨勢分析[J]. 中國漁業經濟,31(3):127-132. [Yuan Y M,Yuan Y,He Y H,Dai Y Y,Gong Y C,Zhang H Y,Wang H W. 2013. Trend analysis on tilapia industry development in China[J]. Chinese Fisheries Economics,31(3):127-132.]
朱佳杰. 2019. 吉富羅非魚抗病與易感家系的篩選及感染無乳鏈球菌后轉錄組和蛋白質組研究[D]. 南寧:廣西大學. [Zhu J J. 2019. Transcriptomics and proteomics analy-ses of high-resistance and susceptible families of genetically improved farmed tilapia after Streptococcus agalactiae infection[D]. Nanning:Guangxi University.]
朱佳杰,周宇,譚蕓,甘西,沈夏霜,劉溪,敖秋桅. 2015. 不同品系羅非魚抗無乳鏈球菌病性能的評估[J]. 安徽農業科學,43(10):168-170. [Zhu J J,Zhou Y,Tan Y,Gan X,Shen X S,Liu X,Ao Q W. 2015. Evaluation on the resistance of different tilapia cultivars to Streptococcus agalactiae[J]. Journal of Anhui Agricultural Sciences,43(10):168-170.]
祝璟琳,鄒芝英,李大宇,肖煒,徐跑,楊弘,薛良義. 2017. 四個羅非魚選育品種抗鏈球菌病能力差異研究[J]. 水生生物學報,41(6):1232-1241. [Zhu J L,Zou Z Y,Li D Y,Xiao W,Xu P,Yang H,Xue L Y. 2017. Study on resistant difference among four bleeding species of tilapia Oreochromis spp. following Streptococcus agalactiae cha-llenge[J]. Acta Hydrobiologica Sinica,41(6):1232-1241.]
鄭衛衛. 2016. 牙鲆家系生長和抗病性能及抗病性狀的遺傳力和育種值分析[D]. 上海:上海海洋大學. [Zheng W W. 2016. Analysis of the trait of growth and disease-resistance and the heritability and breeding value of di-sease-resistance in Japanese flounder[D]. Shanghai:Shanghai Ocean University.]
Huang B F,Zou L L,Xie J G,Huang Z C,Li Y W,Li A X. 2013. Immune responses of different species of tilapia infected with Streptococcus agalactiae[J]. Journal of Fish Diseases,36(8):747-752.
Ozaki A,Sakamoto T,Khoo S,Nakamura K,Coimbra M R,Akutsu T. 2001. Quantitative trait loci(qtls) associated with resistance/susceptibility to infectious pancreatic necrosis virus(ipnv) in rainbow trout(Oncorhynchus mykiss)[J]. Molecular Genetics & Genomics,265(1):23-31.
Storset A,Strand C,Wetten M,Kj?glum S,Ramstad A. 2007. Response to selection for resistance against infectious pancreatic necrosis in atlantic salmon(Salmo salar L.)[J]. Aquaculture,272(S1):62-68.
(責任編輯 思利華)
