干旱脅迫下腐植酸對燕麥葉綠素熒光特性的調控效應
2020-05-13 08:58:30李英浩劉景輝趙寶平朱珊珊戴云仙
灌溉排水學報 2020年4期
關鍵詞:植物
李英浩,劉景輝,趙寶平,田 露,王 琦,朱珊珊,戴云仙
(內(nèi)蒙古農(nóng)業(yè)大學 雜糧產(chǎn)業(yè)協(xié)同創(chuàng)新中心,呼和浩特 010019)
0 引 言
由于全球氣候變化和人類活動的加劇,地球上的水循環(huán)狀況發(fā)生了劇烈的變化,導致很多地區(qū)和流域發(fā)生了嚴重的水資源問題和水環(huán)境危機,水資源緊缺已成為限制農(nóng)業(yè)可持續(xù)發(fā)展的瓶頸[1]。水分是半干旱地區(qū)作物生產(chǎn)的主要因素,提高作物抗逆性以及高效利用水資源,成為我國旱區(qū)促進農(nóng)業(yè)持續(xù)增長的必然選擇[2]。燕麥是禾本科燕麥屬草本植物,抗逆性強,廣泛種植于貧瘠、干旱、冷涼等地區(qū),是糧飼兼用型作物[3]。燕麥根系發(fā)達,吸水能力較強,且根冠比較大,調節(jié)水分能力較強,成為治理土地荒漠化的先鋒作物[4]。【本研究的切入點】干旱脅迫會破壞植物的很多代謝和生理過程[5]。干旱脅迫會導致植物生物量減少,總葉綠素量減少,葉綠素熒光參數(shù)發(fā)生變化[6-12],而光合作用是植物在干旱脅迫下被抑制的最重要的生理過程之一[10],但對于其復雜而精密的生理生化調節(jié)過程尚不明確[13]。水分脅迫對植物光合作用的影響可以通過分析葉綠素熒光動力學參數(shù)進行研究,植物葉片葉綠素熒光參數(shù)的變化可以準確地衡量植物光合潛能的高低和受傷害程度[14]。葉綠素熒光動力學的應用可以深入細致地分析環(huán)境脅迫對植物光合機構尤其是 PSⅡ的影響以及光合機構對環(huán)境變化的適應機制[15]。干旱和澇脅迫均可對作物熒光反應產(chǎn)生抑制作用,如水分脅迫下最大熒光產(chǎn)量、可變熒光、PSⅡ最大光化學量子產(chǎn)量、PSⅡ潛在活性、非光化學猝滅和光化學猝滅[16]。熒光反應是光合功能強弱的重要指標,并且初始熒光值與葉綠素a的量密切相關,可以反映PSⅡ反應活性強弱,而最大熒光值則表現(xiàn)PSⅡ反應中心的電子傳遞情況,F(xiàn)v/Fm反映 PSⅡ反應中心內(nèi)部光能轉換效率。【研究進展】目前,國內(nèi)外關于運用葉綠素熒光參數(shù)以及光合參數(shù)的變化來研究土壤水分脅迫對植物影響的報道較多[17-20],有研究表明重度干旱脅迫導致燕麥葉片凈光合速率,蒸騰速率、氣孔導度以及胞間 CO2摩爾分數(shù)急劇降低[21],前人在麥類作物葉綠素熒光參數(shù)的研究多集中在葉片部位,燕麥苗期遭受干旱脅迫,葉片光系統(tǒng)Ⅱ的原初光能轉化效率Fv/Fm和潛在活性相對較穩(wěn)定,燕麥灌漿前期各光合器官Fv/Fm相對較穩(wěn)定,隨灌水次數(shù)增加各器官值有增加趨勢,但變化很小[22],由此說明燕麥的光合性能對水分脅迫較為敏感,一定程度的干旱脅迫會嚴重影響到燕麥的光合能力。熒光動力學在小麥、玉米、水稻及豆科作物上都已有相關研究,但有關燕麥熒光參數(shù)對水分脅迫的響應研究鮮有報道。腐植酸是動植物遺骸經(jīng)過微生物分解和轉化等一系列過程形成的一類有機物質[23],腐植酸有助于提高作物耐旱性,延緩植株衰老。腐植酸處理明顯提高了小麥灌漿后期旗葉實際光化學效率和光合電子傳遞速率[30],但是至今國內(nèi)外對腐植酸在干旱脅迫下燕麥葉片葉綠素熒光參數(shù)變化研究尚少。【擬解決的關鍵問題】本試驗通過研究外源噴施腐植酸對燕麥葉片葉綠素量及各項熒光參數(shù)Fo、Fm、Fv等在干旱脅迫下的變化狀態(tài),試圖探討干旱脅迫下腐植酸對燕麥葉片葉綠素熒光特性的調控效應,【研究意義】進一步了解腐植酸提高植物耐旱性的機理,為開發(fā)利用腐植酸肥料和提高燕麥抗旱性提供一定理論依據(jù)和技術支持。
1 材料與方法
1.1 試驗材料及設計
試驗于2018年5月—2018年9月在內(nèi)蒙古呼和浩特市內(nèi)蒙古農(nóng)業(yè)大學燕麥產(chǎn)業(yè)研究中心溫室進行。供試燕麥品種為燕科二號,由內(nèi)蒙古農(nóng)牧業(yè)科學研究院提供。腐植酸水溶肥料(HA)由內(nèi)蒙古永業(yè)農(nóng)豐生物技術有限責任公司提供,其水溶腐植酸量≥50 g/L,N+P+K量≥200 g/L,微量元素(錳、硼、鉬、鋅等)量≥10 g/L。采用盆栽方法,土壤為蛭石與泥炭土按質量比為 1∶1的比例混合而成。泥炭土中N+P+K量>10 g/kg,有機質量>50 g/kg,pH值為5.5~8.5,土壤含水率為10%。塑料盆高25 cm,直徑20 cm,每盆裝混合土2.5 kg,播種前底施磷酸二銨(N18%,P2O546%)2 g,在拔節(jié)期與抽穗期各追施尿素(含N量為46%)2 g。5月1日播種,每盆播30粒,在三葉期(5月20日)定苗,每盆20株。試驗采用隨機區(qū)組排列,設置正常供水(75%田間持水率);中度干旱脅迫(60%田間持水率)和重度干旱脅迫(45%田間持水率)3個水分條件,不同水分條件下設噴施500倍腐植酸水溶肥料(HA)和等量清水(CK)2個處理,共6個處理,每個處理重復3次,每個重復種植5盆,共90盆。水分脅迫在拔節(jié)期(6月10日)開始,每天17:00點利用稱重法進行補水以保持各處理土壤含水率穩(wěn)定,土壤田間持水率、永久萎蔫點及土壤質量含水率按Ryan等[24]的方法測定,全生育期3個水分條件的總灌水量分別為27.23、21.78 和16.34 L。分別在拔節(jié)期、抽穗期和灌漿期噴施腐植酸,噴施后7 d進行取樣和測定熒光參數(shù)。
1.2 取樣方法
取植株上部新鮮的旗葉葉片,立即液氮速凍并保存,每個處理重復取樣3次,共6片葉。
1.3 測定指標與方法
1.3.1 熒光參數(shù)
每處理選擇3片生長完好的旗葉葉片,用FMS-2便攜式脈沖調制式熒光儀測定葉綠素熒光參數(shù)。在葉片自然生長角度不變的情況下測定穩(wěn)態(tài)熒光(Fs),隨后加一個強閃光[6 000 μmol/(m2·s)],脈沖 0.8 s測定光適應下的最大熒光(Fm');關閉作用光,暗反應3 s后再打開遠紅光,5 s后測定光下最小熒光(Fo');葉片暗適應15 min后測定初始熒光(Fo),隨后加一個強閃光(6 000 μmol/(m2·s)),脈沖0.8 s測定最大熒光(Fm)。計算如下參數(shù):PSⅡ最大光能轉換效率Fv/Fm=(Fm-Fo)/Fm;PSⅡ潛在光化學活性Fv/Fo=(Fm-Fo)/Fo;光化學淬滅系數(shù)qP=(Fm'-Fs)/(Fm'-Fo');非光化學淬滅系數(shù)NPQ=(Fm-Fm')/Fm'。
1.3.2 葉綠素量
稱0.2 g凍干的燕麥葉片,采用分光光度法[25]測定旗葉葉綠素a(Chla)與葉綠素b(Chlb),最后計算總葉綠素量(Chla+Chlb)和葉綠素a與葉綠素b的比值(Chla/Chlb)。
1.4 數(shù)據(jù)分析
試驗數(shù)據(jù)用Excel進行整理,用SAS 19.0進行統(tǒng)計分析。
2 結果與分析
2.1 噴施HA對干旱脅迫下燕麥葉片葉綠素的影響
干旱脅迫下噴施 HA后燕麥葉綠素量的變化如表1。由表1可知,從拔節(jié)期到灌漿期,燕麥葉片Chla+Chlb逐漸提高。干旱脅迫導致燕麥葉片Chla+Chlb和 Chla/Chlb降低,且降幅隨著干旱脅迫程度加劇而增加。噴施HA后,正常供水條件下二者與 CK相比差異均不顯著;中度干旱脅迫下,葉片Chla+Chlb和Chla/Chlb分別比CK分別平均增加6.69%和3.06%,但只有生育前期差異顯著;而重度干旱脅迫下二者分別比CK平均增加40.82%和30.09%,且均達到顯著差異(P<0.05)。綜上說明,干旱脅迫使燕麥葉片葉綠素總量的合成受阻,分解速率增大,并且 Chla的分解速率超過 Chlb,進而導致 Chla/Chlb降低,而噴施HA可不同程度地緩解干旱脅迫造成的影響,在重度干旱脅迫下對葉綠素合成的促進作用最顯著。
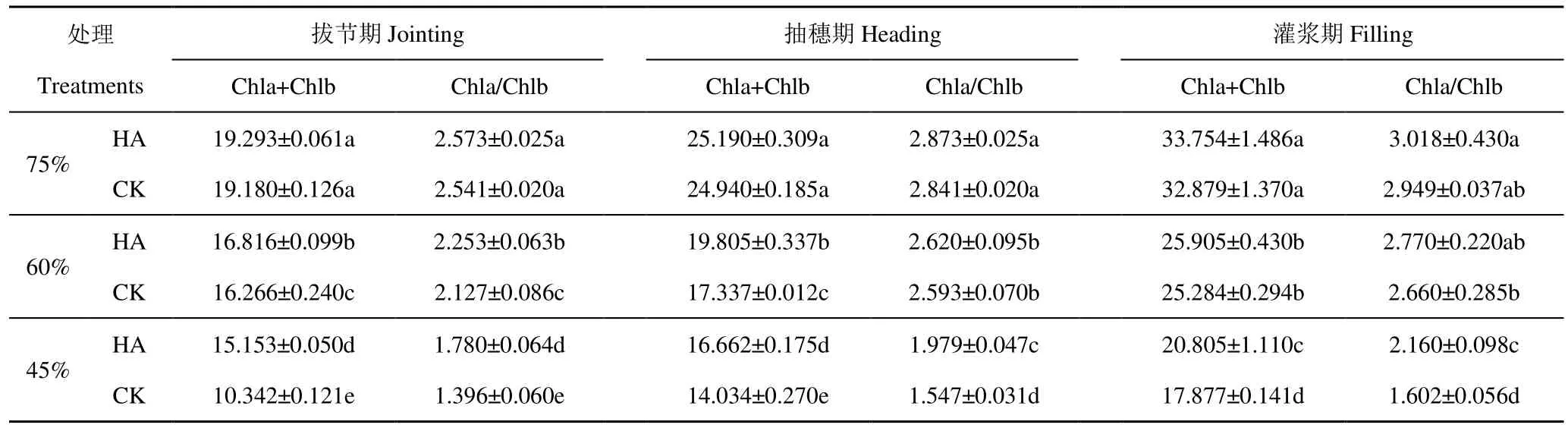
表1 不同處理燕麥葉片Chla+Chlb與Chla/ChlbTable 1 Effect of different treatments on the dry matter quality of oats Chla+Chlb and Chla/Chlb
2.2 HA噴施對干旱脅迫下燕麥熒光參數(shù)的影響
2.2.1Fo、Fm和Fv的變化
干旱脅迫下噴施HA后燕麥葉片F(xiàn)o、Fm和Fv的變化情況如表2。由表2可知,從拔節(jié)期到灌漿期,F(xiàn)o、Fm和Fv均逐漸提高。隨著干旱脅迫程度的增加,F(xiàn)o呈上升趨勢,而Fm和Fv卻顯著下降。干旱脅迫下噴施 HA后Fo下降,降低范圍為 0.70%~14.06%,而Fm和Fv升高,提升范圍分別為0.7%~121.19%和1.0%~171.79%。不同時期中,噴施HA與CK相比,在正常供水和中度干旱脅迫條件下3個指標差異均不顯著,而在重度干旱脅迫條件下三者均差異顯著(P<0.05)。綜上說明,干旱脅迫導致PSⅡ反應中心被破壞或可逆失活,同時會降低燕麥PSⅡ反應中心原初電子受體QA的還原能力,使電子傳遞活性受到抑制,但噴施HA會有效緩解干旱脅迫造成的影響,在重度干旱脅迫條件下效果最顯著。

表2 不同處理燕麥Fo、Fm和FvTable 2 Effect of different treatments on the dry matter quality of oats Fo、Fm and Fv
2.2.2Fm/Fv和Fv/Fo的變化
干旱脅迫下噴施 HA后燕麥葉片F(xiàn)m/Fv和Fv/Fo的變化情況如表3。由表3可知,從拔節(jié)期到灌漿期,F(xiàn)v/Fm和Fv/Fo逐漸提升,但干旱脅迫下Fv/Fm和Fv/Fo均顯著下降(P<0.05)。干旱脅迫下噴施HA與CK相比,F(xiàn)v/Fm和Fv/Fo值出現(xiàn)不同程度的提升,提升范圍分別為 0.2%~83.89%和1.9%~211.56%。3個時期中,噴施HA后相比于CK,正常供水和中度干旱脅迫條件下二者均無顯著差異,而在重度干旱脅迫條件下二者均達到顯著差異(P<0.05)。故干旱脅迫抑制了燕麥 PSⅡ的潛在活性并降低了PSⅡ的原初光能轉換效率,HA對重度干旱脅迫造成的傷害有最明顯的緩解效應。

表3 不同處理燕麥Fv/Fm和Fv/FoTable 3 Effect of different treatments on the dry matter quality of oats Fv/Fm and Fv/Fo

表4 不同處理燕麥qP和NPQTable 4 Effect of different treatments on the dry matter quality of oats qP and NPQ
2.2.3qP和NPQ的變化
干旱脅迫下噴施HA后燕麥葉片qP和NPQ的變化如表4。由表4可知,從拔節(jié)期到灌漿期,燕麥葉片qP和NPQ逐漸提高。水分脅迫會導致qP值顯著降低,NPQ值則出現(xiàn)顯著提升,表明水分脅迫下燕麥光能利用率上升,熱量散失減少。噴施HA后相比于CK,qP和NPQ值會有所提高,提高范圍分別為0.1%~68.30%和6.02%~73.36%,表明在干旱脅迫下HA會進一步提高燕麥對光能的利用效率,減少熱量散失。噴施HA后,同樣只在重度干旱脅迫條件下二者均差異顯著(P<0.05)。綜合以上結果說明,干旱脅迫對燕麥光反應中心造成了傷害,且干旱脅迫程度越大對植株的傷害越大,而噴施HA則會在一定程度上緩解脅迫對光反應中心造成的傷害,提高燕麥光能利用率,且重度干旱脅迫條件下HA效果最佳。
3 討 論
葉綠素是植物進行光合作用的物質基礎,葉綠素量的多少會影響植物光合作用能力。Chla/Chlb可反映出葉綠素a與葉綠素b對脅迫的敏感性及脅迫對二者的傷害程度,本試驗研究發(fā)現(xiàn),干旱脅迫導致燕麥葉片Chla/Chlb降低,說明葉綠素b對干旱脅迫的敏感性更大。總葉綠素量標志著該生命體光合能力的強弱,一般情況下,干旱脅迫會導致植物葉片失水,進而抑制葉綠素的生物合成,并促進已合成的葉綠素分解,使葉片葉綠素量下降,從而影響植株的光合作用[26-28]。但也有研究發(fā)現(xiàn),干旱脅迫會使植物葉綠素量增加,可能是因為干旱脅迫使葉片含水率降低,葉片擴展生長受阻,葉片單位面積的葉綠素量升高[29]。這2種不同的研究結果表明植物處于逆境時的葉綠素量變化趨勢可能與植物本身的耐旱性有關,本試驗中,重度干旱脅迫導致燕麥葉片總葉綠素量急劇下降,說明燕麥已無法適應重度干旱脅迫環(huán)境。施用適量的腐植酸會增加作物葉片葉綠素量[30],小麥經(jīng)腐植酸浸種后,使葉片葉綠素量提高,提高Chla/Chlb[31]。本研究得出,干旱脅迫下噴施腐植酸后,燕麥葉片Chla+Chlb和Chla/b均有不同程度的提高,與張金政等[26]、梁文斌等[27]的研究結果Chla/Chlb一致,同時腐植酸可以提高不同水分脅迫處理油菜葉綠素量[32],在重度干旱脅迫下噴施腐植酸后,燕麥葉片Chla+Chlb和Chla/b的提高幅度最大。
本試驗中,燕麥Fo上升,F(xiàn)m和Fv下降,說明干旱脅迫降低了PSⅡ反應中心原初電子受體QA的還原能力,QA→QB的能力下降,電子傳遞活性降低[33-36]。植物在逆境條件下或受到傷害時Fv/Fm 會明顯下降。干旱脅迫下燕麥Fo升高,尤其以重度干旱脅迫下升高幅度最大,說明干旱脅迫嚴重破壞光反應系統(tǒng)的穩(wěn)定性,使其光合速率降低[37]。干旱脅迫使PSⅡ捕光色素蛋白復合體(LHC-Ⅱ)量下降,使LHC-Ⅱ捕光色素吸收的能量減少[38],從而導致葉綠體激發(fā)能LHC-Ⅱ向PSⅡ的傳遞減少,以至光合電子由PSⅡ反應中心向QA、QB及PQ庫傳遞過程受到影響[39],本研究中Fv/Fm和Fv/Fo的顯著下降(P<0.05),表明干旱脅迫使燕麥PSⅡ反應中心受到傷害,抑制了燕麥PSⅡ的潛在活性并降低了PSⅡ的原初光能轉換效率,從而使燕麥光合作用的原初反應受到抑制[40]。熒光淬滅包括光化學淬滅qP和非光化學淬滅NPQ。qP反映的是PSⅡ天線色素吸收的光能用于光化學傳遞的份額,也表示PSⅡ天線色素吸收的光能用于暗反應固定能量的部分,其值越高表示PSII的電子傳遞活性越大,光能中轉變?yōu)榛顫娀瘜W能的能量越多,植物對光能的利用效率也越高[41]。NPQ反映的是PSⅡ天線色素吸收的光能不能用于光合傳遞而以熱的形式耗散掉的部分[42],其值大小表示光合機構的損傷程度[43]。NPQ在植物吸收光能過多情況時起到一定的自我保護作用[44-45]。本試驗發(fā)現(xiàn),干旱脅迫導致燕麥qP逐漸下降,NPQ逐漸上升,可能原因是干旱脅迫破壞了PSⅡ的反應中心系統(tǒng),導致燕麥葉片光化學活性降低,并且干旱逆境下使燕麥減少熱量散失,提高了光能利用率,這與孫雅麗等[46]在越桔上的研究結果類似。
腐植酸處理明顯提高了小麥灌漿后期旗葉的光能轉化效率(Fv/Fm)和反應中心的光化學活性(Fv/Fo),尤其以旱作條件下增加幅度較高[29],本試驗結果顯示,從燕麥的拔節(jié)期到灌漿期,干旱脅迫下腐植酸均可以提高其Fv/Fm和Fv/Fo,且重度干旱脅迫條件下提高幅度最大,驗證了上述研究結果。
4 結 論
干旱脅迫顯著降低燕麥葉片總葉綠素量、Chla/Chlb、最大熒光(Fm)、可變熒光(Fv)、PSⅡ原初光能轉化效率(Fv/Fm)、PSⅡ的潛在活性(Fv/Fo)和光化學淬滅(qP),而提高了最小熒光(Fo)和非光化學淬滅(NPQ)。噴施腐植酸可緩解干旱脅迫對葉綠素熒光反應的影響,且與正常供水和中度脅迫相比,重度干旱脅迫條件下腐植酸的抗旱效果最佳。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
