嗜水氣單胞菌非核糖體肽合成酶基因功能研究
2020-05-13 14:56:18張麗珊孫莉娜林鎮平林向民
生物技術通報 2020年4期
關鍵詞:生長
張麗珊 孫莉娜 林鎮平 林向民
(福建農林大學生命科學學院 福建省農業生態過程與安全監控重點實驗室,福州 350002)
隨著全球人口的增加,水產養殖正成為人類飲食中魚蛋白的主要來源。然而,伴隨著水產養殖業的發展,魚類的各種傳染性疾病也日益嚴重[1-2]。常見的水產致病菌有嗜水氣單胞菌、副溶血性弧菌、溶藻弧菌等,可引起出血性敗血癥[3]。這些疾病的爆發與傳播給水產養殖業造成的損失也越來越嚴重。為了防治水產養殖業的感染性疾病,大量的抗生素(慶大霉素、利福平、四環素等)被投入使用,而抗生素濫用使得細菌的耐藥性日益嚴峻,耐藥菌株層出不斷,同時也伴隨著環境中藥物殘留污染的現象越發嚴重[4-5]。因此,探究其致病機制被認為是控制水生病原體感染最有效的途徑之一。
嗜水氣單胞菌(Aeromonas hydrophila)作為水體中一種常見的致病菌,能產生腸細胞毒素、溶血素、蛋白酶和血凝素等毒力因子,引發魚類患出血性敗血癥[6]。在體內,鐵通常由于其特殊的物理化學性質而被氧化成不溶形式,并與細胞內的血紅素、鐵蛋白、血紅蛋白和轉鐵蛋白結合,因此細菌不易獲得[7]。為了應對這種鐵缺乏癥,微生物已經進化出一系列復雜的機制來與宿主競爭,例如鐵載體的分泌,以從轉鐵蛋白、血紅蛋白和鐵蛋白中捕獲鐵并維持鐵動態平衡以促進細菌生長、增殖和毒素分泌[8]。非核糖體肽是一類次級代謝產物,通常在細菌和真菌等微生物中產生。與在核糖體上合成的多肽不同,非核糖體肽合成酶(Non-ribosomal peptide synthetase,NRPS)由相互獨立的多個模塊組成,模塊具有的特異結構域及酶活,催化相鄰兩模塊上供體多肽和受體氨基酸的肽鍵形成[9-10]。作為次生代謝產物,NRP 可以產生比核糖體更廣泛的多肽。它們在結構上是一個非常多樣化的天然產物家族,具有極其廣泛的生物活性和藥理學特性,可被用作激素、抗生素、鐵載體、抗真菌、抗癌等豐富多樣的生物活性[11]。微生物鐵載體在人類,動物和植物的可持續性方面的應用范圍很廣。基于非核糖體肽作為鐵載體時在各個方面的有益應用,如細菌利用合成的鐵載體攫取外界游離Fe3+來維持正常的生理活動。在嗜水氣單胞菌ATCC7966 基因組中AHA_2473-AHA_2479是一個反映鐵載體的生物合成和調節的基因簇,在這個基因簇中AHA_2474、AHA_2476是兩個編碼非核糖體肽合成酶的基因,目前對于這兩個基因功能的研究比較少。本研究通過同源重組方法構建嗜水氣單胞菌AHA_2474(nonribosomal peptide synthetase)、AHA_2476(nonribosomal peptide synthetase)基因缺失菌株(ΔAHA_2474、ΔAHA_2476),利用Chrome azurol sulfonate(CAS)檢測法對缺失菌產鐵情況進行測定,并利用M9 限制性培養基觀察及補充鐵的條件下野生菌和缺失菌的生長情況,最后通過DIP 條件下測生長曲線及測定溶血性、胞外蛋白酶活性、過氧化氫耐受性比較AHA_2474、AHA_2476基因的缺失對細菌生長的影響,分析AHA_2474、AHA_2476的功能與調節機制,為更好地認識細菌生理功能調控機制提供理論基礎[6,12]。
1 材料與方法
1.1 材料
本研究使用的嗜水氣單胞菌ATCC7966、大腸桿菌(MC1061、S17)及質粒(pRE112)均保存在本實驗室中,除嗜水氣單胞菌培養在30℃,其余都培養在37℃的LB 培養基中;LB 培養基成分:胰蛋白酶胨、酵母粉、氯化鈉均購于生工生物工程(上海)股份有限公司;限制性內切酶購于TaKaRa 公司,連接酶購于諾唯贊生物公司,Taq 酶購于翊圣生物科技有限公司,無菌脫纖維綿羊血、脫脂牛奶購買于北京索萊寶科技有限公司
1.2 方法
1.2.1 構建缺失菌株 利用與自殺載體pRE112 同源重組原理,以嗜水氣單胞菌ATCC7966 的基因組DNA 為模板構建兩側為AHA_2474、AHA_2476基因上、下游同源序列片段的基因敲除質粒,將構建好的質粒轉入大腸桿菌MC1061 感受態細胞,經菌液PCR 驗證正確后提取質粒再轉入大腸桿菌S17感受態細胞,再經菌液PCR 驗證正確后利用接合法與野生株(wild type,WT)同源重組,通過100 μg/mL 氨芐青霉素、30 μg/mL 氯霉素抗性篩選,獲得第一次同源重組菌株之后在含有20%蔗糖的LB培養基中進行第二次同源重組,經氯霉素抗性平板篩選、菌液PCR 和DNA 測序確認嗜水氣單胞菌AHA_2474、AHA_2476基因缺失突變株。最后通過穩定遺傳20 代后,測序正確后保菌在-80℃冰箱中。
1.2.2 菌株培養 表型驗證的菌株均從LB 平板上分別挑取野生株、ΔAHA_2474和ΔAHA_2476單一菌落接種于5 mL LB 培養基中,30℃,200 r/min 過夜培養16h至穩定狀態。取過夜菌按1∶100 比例轉接至5 mL 新鮮 LB 中,30℃,200 r/min,搖至OD=1.0后使用。
1.2.3 溶血性和胞外蛋白酶活性 將OD=1.0 的野生株、ΔAHA_2474和ΔAHA_2476各取5 μL 接種于已打孔的綿羊血平板(綿羊血含7%)和脫脂牛奶平板(脫脂牛奶含1%)上,置于30℃培養箱培養12-18 h,觀察溶血圈和水解圈的大小,測量圈的大小以及拍照保存,實驗均重復3 次。
1.2.4 ΔAHA_2474、ΔAHA_2476突變株在DIP條件下生長曲線的測定 將OD=1.0 的野生株、ΔAHA_2474和ΔAHA_2476按1∶100加入1 mL LB培養基中并加入終濃度為200 μmol/L 鐵離子螯合劑2,2'-聯吡啶(DIP),最后以每孔300 μL 加入微孔板中,對照組、實驗組均做3 個生物學重復,放入30℃、波長600 nm 全自動生長曲線分析儀中,測定16 h,每小時測定一次吸光值。
1.2.5 CAS 檢測液測定 配制CAS 檢測液:(1)配制CAS 染色液,溶液A:將0.079g的鉻天青(CAS,上海試劑公司)溶于50 mL 去離子水中,再加入10 mL 1mmol/L 的FeCl3溶液(含有10 mmol/L 的HCl);溶液B:將0.069g的十六烷基三甲基溴化銨(HDTMA)溶于40 mL 的去離子水中;溶液C:將A 溶液沿著燒杯的壁緩緩加入到B 溶液中,輕輕晃動,使溶液AB 混勻,得到溶液C:CAS 染色液,121℃滅菌15 min。(2)配制0.1 mol/L 磷酸鹽緩沖液(pH6.8),121℃滅菌15 min,使用時稀釋10 倍。(3)配制CAS 培養基:先配制好1 mmol/L 的CaCl2溶 液、1 mmol/L 的MgSO4·7H2O溶液、10%的酸水解酪蛋白溶液、20%蔗糖;再分別取0.1 mL 的CaCl2溶液、2 mL 的MgSO4·7H2O溶液、3 mL 的10%的酸水解酪蛋白溶液,20%蔗糖1 mL,去離子水定容到100 mL,115℃滅菌15 min。當以上的培養基滅完菌后,溫度降低到60℃時,加入稀釋好的0.1 mol/L 磷酸鹽緩沖液5 mL 及8 mL CAS 染色液,每100 mL CAS 培養基的量沿著三角瓶壁加入,混合均勻后取5 mL 分裝至已滅菌的試管中。將OD=1.0 的野生株、ΔAHA_2474和ΔAHA_2476菌株按1∶100 轉接至分裝好CAS 試管中,30℃,200 r/min 搖床中培養16h后觀察生長狀況并拍照保存。
1.2.6 M9 限制性培養基測定 配制M9 限制性培養基:5×M9 鹽溶液(Na2HPO4·7H2O 0.678 g,KH2PO40.3 g,NaCl 0.05 g,NH4Cl 0.1 g)20 mL、20%葡萄糖2 mL、1 mol/L MgSO4200 μL、1 mol/L CaCl210 μL,加入蒸餾水補足100 mL,并且每100 mL 加入1.8g瓊脂粉,121℃,滅菌15 min 后,倒在培養皿中。分別制備LB 平板,M9 平板及加有終濃度為50 μg/mL FeCl3的M9 平板,將OD=1.0 的野生株、ΔAHA_2474、ΔAHA_2476分別取2 μL 劃在平板中,置于30℃恒溫培養箱中培養16h后觀察其生長狀況并拍照保存,并重復3 次。
1.2.7 對過氧化物的耐受性測定 將OD=1.0 的野生株、ΔAHA_2474、ΔAHA_2476按1%的比例將菌體加入到1 mL 液體LB 中,實驗組加入稀釋5 倍的H2O2,使其作用濃度為2.5 mmol/L,對照組不作處理,室溫應激10 min 后對樣品進行梯度稀釋(10-2-10-7),對每個稀釋梯度的樣品各取2 μL 點板接種于LB 瓊脂平板上;30℃培養12-16h后觀察結果并拍照保存實驗記錄,實驗需重復3 次。
2 結果
2.1 非核糖體肽合成酶基因敲除
利用軟件CE、Design.V1.03 及NCBI 數據庫在目的基因上下游各500 bp 設計引物分別為P1P2、P3P4(表1),之后進行PCR 擴增,通過多片段連接酶無縫連接到pRE112 載體上再分別轉入MC1061、S17 感受態中,再經同源重組和含20%蔗糖的LB 平板篩選及氯霉素抗性篩選,獲得AHA_2474、AHA_2476敲除菌株,通過目的基因兩端引物P5P6,P1 上游和P4 下游100 bp 左右設計引物P7P8 進行菌液PCR 驗證,并以野生株為對照獲得敲除菌ΔAHA_2474、ΔAHA_2476(圖1)。
2.2 ΔAHA_2474、ΔAHA_2476對胞外蛋白酶活性與溶血性的影響
胞外蛋白酶和外毒素是嗜水氣單胞菌產生的兩大毒力因子,毒力因子的存在導致了嗜水氣單胞菌的致病性,嚴重影響魚類的健康生存甚至威脅到人類的身體健康,因此研究嗜水氣單胞菌的毒力因子具有重要的意義。本研究通過脫脂牛奶平板和綿羊血平板檢測胞外蛋白酶活性和溶血性,發現野生菌和ΔAHA_2474和ΔAHA_2476突變株在其菌落外圍均能形成水解圈,即均具有胞外蛋白酶活性,但ΔAHA_2474突變株的水解圈略微大于野生菌的水解圈,ΔAHA_2476突變株的水解圈顯著大于野生菌的水解圈(圖2-A),表明AHA_2474和AHA_2476基因缺失后細菌胞外蛋白酶活性增強,說明AHA_2474和AHA_2476基因參與了蛋白酶的產生,使該菌毒性增強;而溶血性實驗的結果表明野生株和AHA_2474、AHA_2476突變株在其菌落外圍都能形成溶血圈,但ΔAHA_2474和ΔAHA_2476突變株的溶血圈明顯大于野生菌的溶血圈(圖2-B),表明AHA_2474和AHA_2476基因缺失后細菌溶血性的形成能力增強,說明AHA_2474和AHA_2476基因參與了溶血素的產生,使該菌毒性增強。

表1 基因敲除引物
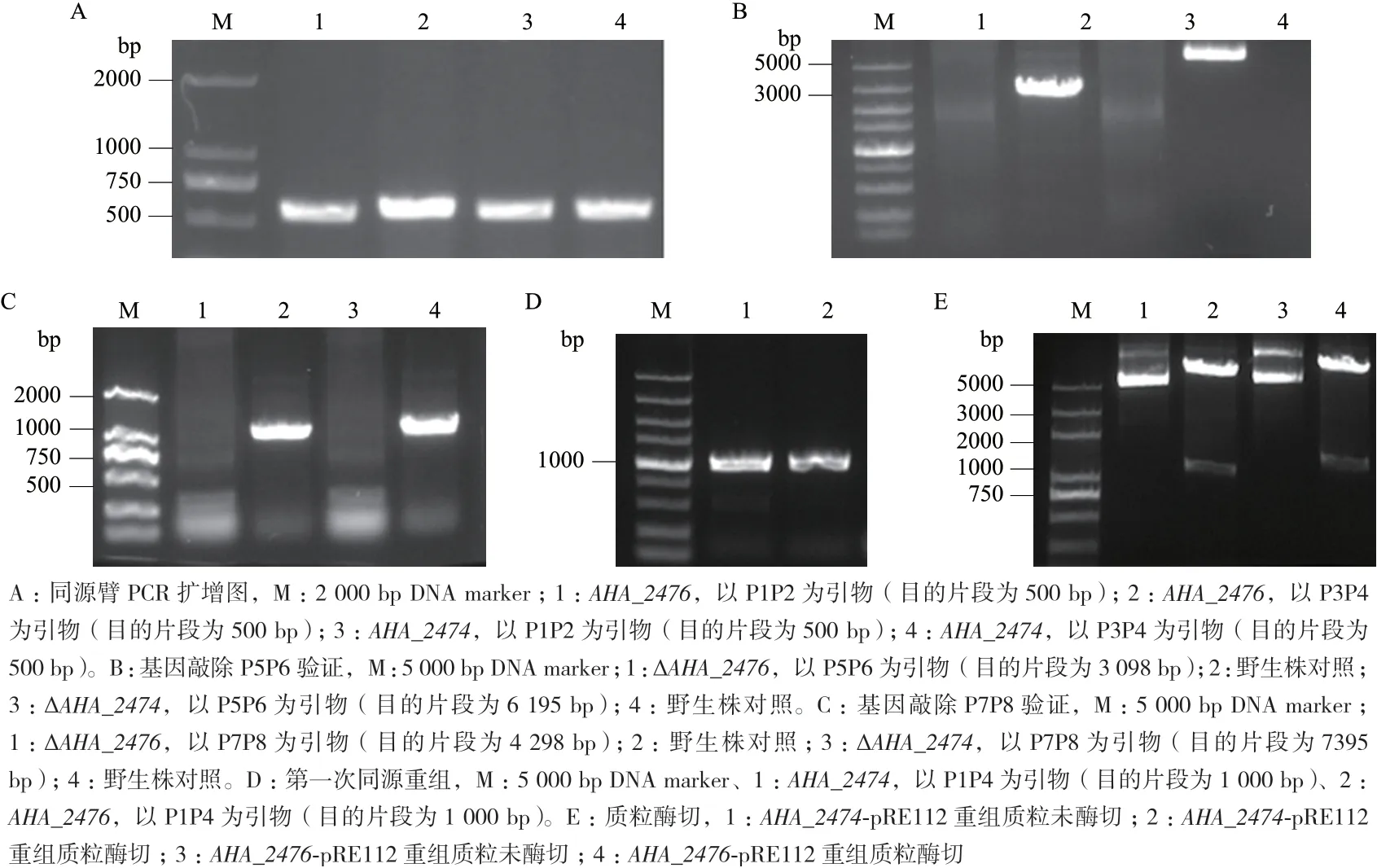
圖1 基因缺失突變株的構建
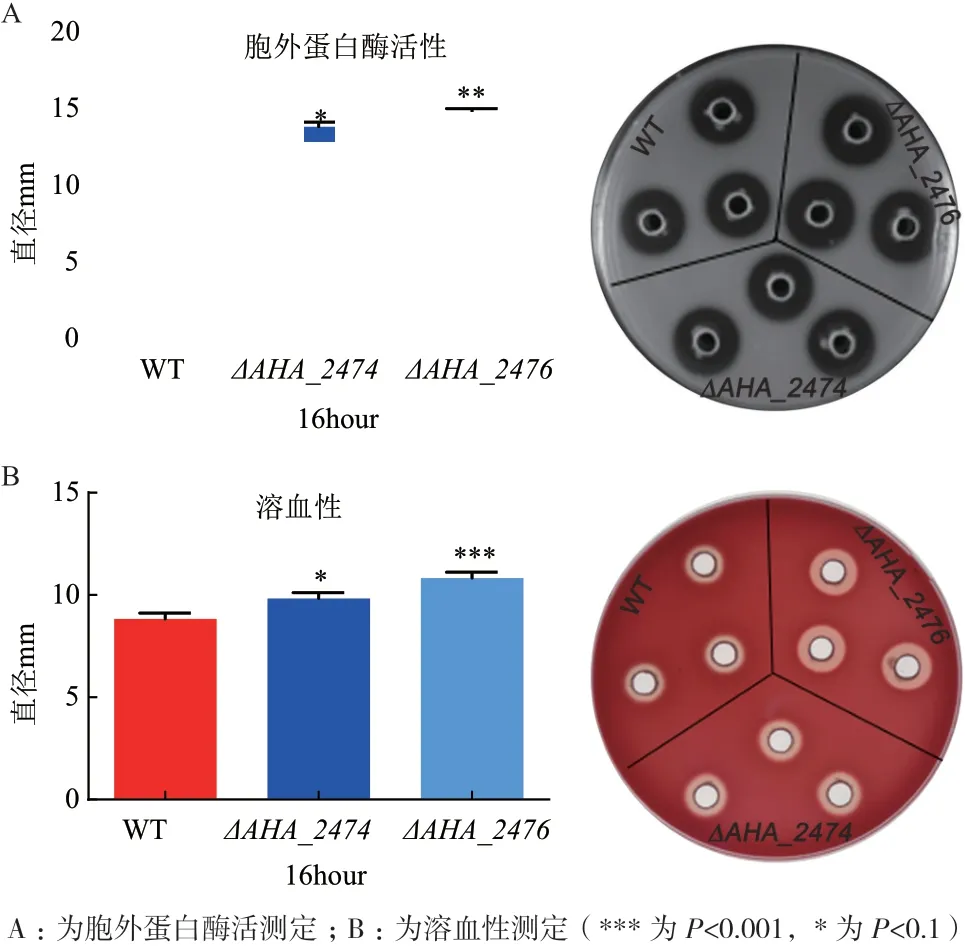
圖2 野生菌與ΔAHA_2474、ΔAHA_2476 敲除菌在體外胞外蛋白酶和溶血性的比較
2.3 DIP對ΔAHA_2474、ΔAHA_2476生長情況的 影響
因ΔAHA_2474、ΔAHA_2476與鐵離子相關,為了探索在鐵離子限制條件下是否對缺失菌的生長產生影響,本研究比較了野生型菌株與ΔAHA_2474、ΔAHA_2476敲除菌株在缺鐵條件下的生長能力。當加入鐵離子螯合劑DIP 后,野生株生長顯著高于ΔAHA_2474、ΔAHA_2476(圖3),說明ΔAHA_2474、ΔAHA_2476可能通過影響鐵離子的動態平衡來影響突變株的生長。
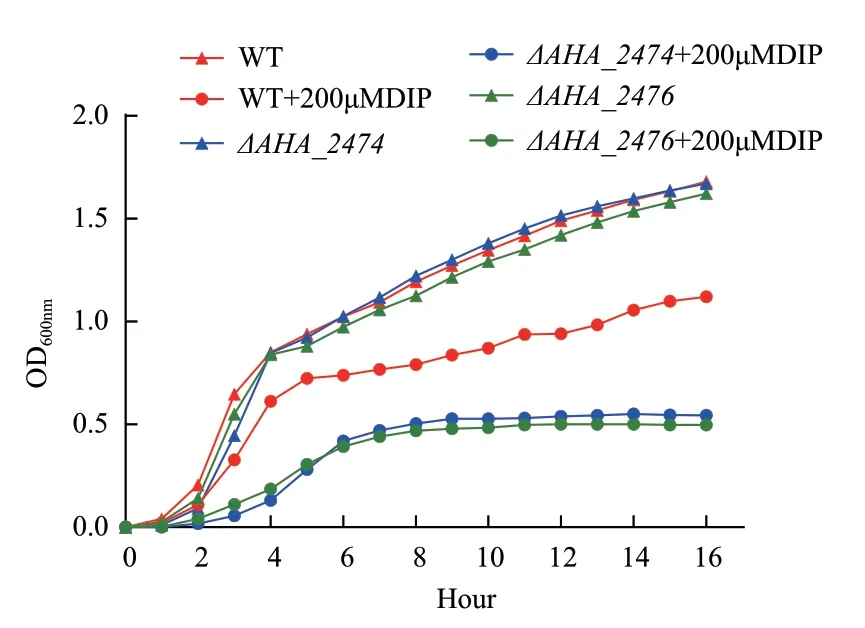
圖3 DIP 條件下野生型菌株和DAHA_2474、DAHA_2476生長能力的比較
2.4 CAS培養基測定
細菌分泌的鐵載體能鰲合CAS 染液中Fe3+而導致顏色由藍變橙,從而測定微生物合成嗜鐵素的能力。利用CAS 液體培養基觀察細菌鐵載體的產生,菌株在 CAS 液體培養基中的顏色的變化。與野生株相比,缺失菌ΔAHA_2474、ΔAHA_2476在檢測培養基上的生長和顏色變化慢,培養12h就可見試管中野生型菌株出現明亮醒目的橙黃色液體(圖4),而缺失菌ΔAHA_2474僅出現微弱的鐵載體,使得試管中僅出現微弱的橙黃色,但ΔAHA_2476在同一時間是顏色還處于藍色,說明缺失菌ΔAHA_2474、ΔAHA_2476影響鐵載體的產生。

圖4 CAS 培養基檢測野生株和ΔAHA_2474、ΔAHA_2476鐵載體產生情況
2.5 M9限制性培養基測定結果
M9 限制性培養基通過對鐵離子的限定來測定。 通過補充50 μg/mL 的FeCl3測定野生株與ΔAHA_2474、 ΔAHA_2476突變菌株的生長情況,發現當在缺鐵的M9 培養基下,缺失菌ΔAHA_2474、ΔAHA_2476的生長要比野生型弱,而在M9 培養基中補充終濃度為50 μg/mL FeCl3時發現野生株與缺失菌ΔAHA_2474、ΔAHA_2476的生長能力相當(圖5)。
2.6 對過氧化物的耐受性測定
為了解ΔAHA_2474、ΔAHA_2476是否參與調節對氧化壓力的應激過程,從而有助于嗜水氣單胞菌適應外界環境的突變情況,本研究測定了ΔAHA_2474、ΔAHA_2476對過氧化物的應激能力。實驗結果表明(圖6),ΔAHA_2474突變株對H2O2(2.5 mmol/L)敏感;而ΔAHA_2476不敏感,與野生菌株相比,ΔAHA_2474菌株具有更大的耐受性,以上結果說明,AHA_2474基因可能參與嗜水氣單胞菌的氧化應激生理過程。
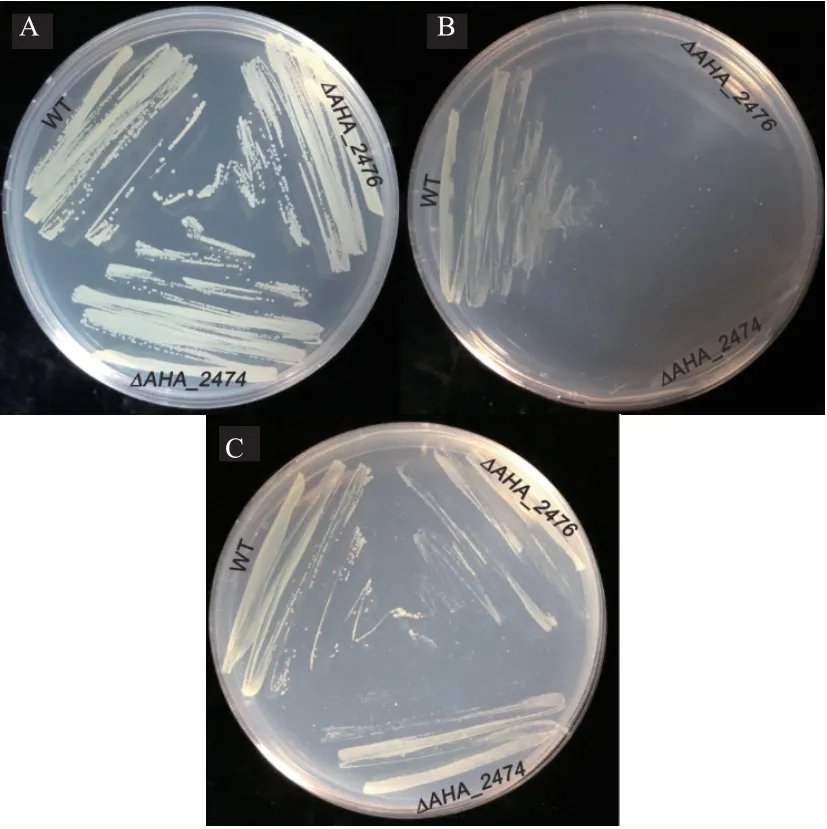
圖5 測定鐵離子限制條件下野生株和ΔAHA_2474、ΔAHA_2476 生長能力的比較
3 討論

圖6 在2.5 mmol/L H2O2 處理下,不同稀釋倍數下野生株、ΔAHA_2474 和ΔAHA_2476 菌株耐受性測定
通常,細菌必須從宿主或復雜的自然環境中捕獲有限的鐵資源,以滿足生物體對鐵的正常需求。因此,細菌已經進化出多種獲取鐵的方法,包括通過特定受體和轉運系統的鐵載體介導鐵的獲取。鐵載體是一種低分子量小于10 kD 的高親和力鐵離子特異性螯合劑,在鐵饑餓時會被細菌分泌出胞外,以螯合劑的形式與宿主或者環境競爭可利用的鐵離子,從而維持細菌的正常生長[13]。鐵在病原體的代謝過程中起重要作用,例如病原體中的毒力,氧化應激耐受性等[14]。已有研究表明,鐵載體在不同的細菌中的功能具有多樣性,如有研究發現,當鐵離子相關基因(tonB)缺失后,愛德華氏菌的產鐵能力下降,毒力、溶血能力以及形成鐵載體的能力都會產生顯著變化[15-17];研究表明,大腸桿菌儲鐵蛋白Dps 可以通過鐵氧化酶活性的變化對過氧化氫的耐受性產生影響,從而進一步影響細菌的存活[18-19]。嗜水氣單胞菌的致病性與溶血素、胞外蛋白酶、粘附因子等多種毒力因子的分泌密切相關[20]。有研究表明,鐵限制條件下能夠誘導細菌的胞外蛋白酶活性[21]。因此,當敲除掉嗜水氣單胞菌的非核糖體肽合成酶基因AHA_2474,AHA_2476基因后,可能影響非核糖體肽整個基因簇的功能,造成該菌對非核糖體肽的合成能力減弱,從而導致鐵載體的量減少,使該菌處于低鐵狀態,進而誘導AHA_2474,AHA_2476敲除株的胞外蛋白酶活性增強。而非核糖體肽是細菌的次生代謝產物,可以以鐵載體的形式與鐵離子高親和性螯合,參與病原菌的毒性、抗逆性等多種重要的生理功能。Qassim 等[22]研究發現,伯克霍爾德菌的非核糖體肽合成酶在鐵載體的合成過程中起著非常重要的作用,當菌株在鐵受限的條件下生長時,非核糖體肽合成酶的基因簇會指導合成鐵載體,進而維持細菌對鐵的正常需求。本研究發現敲除掉嗜水氣單胞菌非核糖體肽合成酶的基因后,該菌的生理功能也有著顯著變化,如胞外蛋白酶活性及溶血性均增強,產鐵能力下降,對過氧化氫的耐受性增強,表明非核糖體肽在嗜水氣單胞菌的生理功能中起著非常重要的作用。綜上所述,本研究通過基因敲除的方法,獲得敲除株ΔAHA_2474、ΔAHA_2476,對其基因功能進行研究,為探究該菌的致病機制及防治策略提供理論依據。
4 結論
本研究通過同源重組方法成功構建了嗜水氣單胞菌ATCC7966 中AHA_2474與AHA_2476的突變株。通過比較缺失株與野生株之間的胞外蛋白酶活性,溶血性存在顯著性的差異,表明這兩個基因的缺失可能使該菌毒性增強。又發現AHA_2474、AHA_2476的缺失導致細菌在鐵離子限制條件下,菌株的生長能力顯著下降,而補充鐵離子后又能恢復生長。一系列的生物學實驗表明AHA_2474、AHA_2476基因的缺失可能是導致嗜水氣單胞菌產鐵能力下降的重要原因之一。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14