廣西欽州灣海域游泳動物群落種類組成及多樣性研究
2020-05-22 08:32:50莫珍妮賴廷和何斌源劉昕明李英花寧秋云
應用海洋學學報 2020年2期
莫珍妮,賴廷和,何斌源,劉昕明,李 夢,李英花,寧秋云
(廣西壯族自治區海洋研究院,廣西 南寧 530022)
海灣是人類從事經濟活動的基地,是陸地與海洋相互作用的過渡區域,極易受到環境變化的影響,也是陸-海生態系統的敏感地帶[1],同時又是海洋生物的重要繁育場所和維持生物多樣性的關鍵區域[2],尤其對游泳動物類群變化影響極大。游泳動物處于海洋食物鏈的頂端,在海洋生態系統中占據重要地位;同時,它還是人類獲取動物蛋白質的重要來源,是海洋漁業的主要利用對象[3]。
近年來,隨著《廣西北部灣經濟區發展規劃》[4]的實施,以能源、石化等為首的重大臨海工業項目陸續建成投產或開工建設,使得社會經濟快速發展、海洋資源有效開發利用的同時,也造成了部分海域環境污染、海洋生態破壞、漁業資源受損等問題,尤以欽州灣為重災區。廣西防城港核電廠東臨欽州灣,其營運期間,產生的溫排水、余氯排放將使周邊海域生態系統受到較大擾動。針對該海域游泳動物群落現狀,本研究擬利用 2015—2016 年開展的防城港核電水域生物資源調查數據,分析欽州灣游泳動物群落結構及多樣性,為系統開展欽州灣漁業資源的管理與修復提供基礎信息。
1 材料與方法
1.1 生境概括
欽州灣位于北部灣沿岸灣頂中部,在 21°33′20″~21°54′30″N,108°28′20″~108°45′30″ E 之間。由內灣(茅尾海)、外灣(欽州灣) 以及連接兩灣的潮汐通道構成,中間狹窄、兩端寬闊,是一個半封閉的天然海灣[5]。全灣岸線長度為 511.65 km,海域面積為 1 910.07 km2。欽州灣潮汐為正規全日潮類型,氣候主要受季風環流影響,氣溫年差較大,熱量豐富,雨量集中[6]。調查海域欽州灣的溶解氧含量較高,硫化物、重金屬的含量較低,均優于一類海水標準;表征有機污染程度的化學需氧量、揮發酚含量基本滿足一類海水標準;無機氮和無機磷在分布上可能受河流輸入和臨港工業排污等的影響,均呈現出灣內高、灣外低,近岸高、遠岸低的分布規律。
1.2 站位布設和采樣分析方法
本研究于2015年8月和2016年8月,連續兩年對欽州灣6個站位及企沙西南、竹山西南海域各一個站位進行拖網采樣調查研究(圖1)。站位選擇以防城港核電監測站位為依據,欽州灣S1~S6站位位于防城港核電站附近海域;竹山西南D2站位位于中越交界的北侖河口海域,離核電廠較遠;企沙西南D1站位位于核電廠和北侖河口之間,從而形成6個核電廠鄰近站位與2個對照站位的監測格局。采樣按照《海洋生物生態調查技術規程》[7]要求使用單船有翼單囊拖網,網具網體長度為7 m,網口高1.5 m,寬3.2 m,囊網網目為15 mm,每站拖拽時間為15 min,平均拖速約為3 n mile/h。漁獲樣品分類、標記后用塑料樣品袋盛裝,放入裝有冰塊的泡沫箱中臨時保存,然后帶回實驗室進行分析鑒定,海上取樣及室內分析方法均按《海洋調查規范》[8]進行;魚類生態類型判別主要依據《南海魚類志》[9]、《廣西北部灣海洋硬骨魚類圖鑒》[10]等。
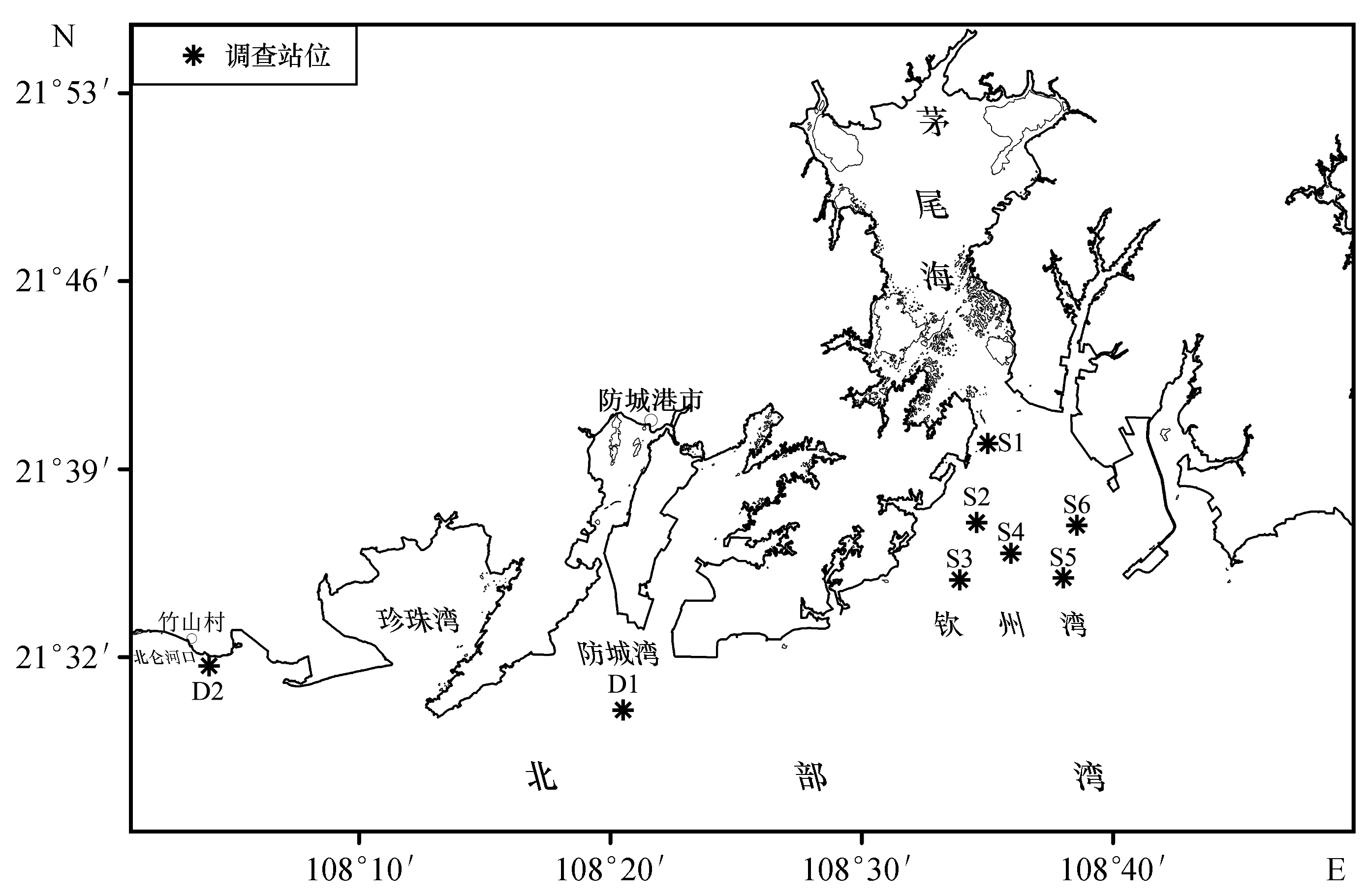
圖1 采樣站位示意圖Fig.1 Map of sampling stations
1.3 數據分析
1.3.1 漁獲物優勢種 漁獲物優勢種分析采用Pinkas(1971)的相對重要性指數IRI計算公式[11]:
IRI=(N+W)·F
(1)
式(1)中:N為某種類的尾數占總漁獲尾數的百分比;W為某種類的質量占總漁獲質量的百分比;F為某種類在調查中的出現頻率。
1.3.2 資源密度 資源密度D按下式計算[12]:
D=C/(q·A)
(2)
式(2)中:C(kg/h或ind./h)為漁獲物密度指數,即每小時取樣面積內的生物量(kg)或尾數(ind.);q為網具捕獲率,在此取與底拖網相同數值0.5;A(km2/h)為網具每小時掃海面積(km2)。
1.3.3 群落多樣性指數 游泳動物群落多樣性指數采用以下指數計算[13-15]:
Shannon-Wiener指數:
Pielou均勻度指數:
J′=H′/log2S
(4)
Margalef物種豐富度指數:
D=(S-1)/log2N
(5)
式(3~5)中:S為種類總數;N為所在群落的所有物種的個體數之和;Pi為調查中第i種漁獲物的個數占樣品漁獲個數的比例。
2 結果與討論
2.1 種類組成
2015年欽州灣拖網定點調查捕獲游泳動物共鑒定出55種[圖2(a)],其中魚類31種,占總種數的56.4%;蝦類11種,占20.0%;蟹類11種,占20.0%;頭足類1種,占1.8%;口足類1種,占1.8%。作為對照海區的企沙西南海域捕獲游泳動物20種,魚類居多,共15種,占75.0%;竹山西南海域共鑒定游泳動物16種,魚類13種,占81.3%。
2016年欽州灣拖網定點調查捕獲游泳動物55種[圖2(b)],其中魚類35種,占總種數的56.4%;蝦類13種,占23.6%;蟹類8種,占14.5%;口足類2種,占3.6%;頭足類1種,占1.8%。企沙西南海域捕獲游泳動物17種,魚類居多,共10種,占58.8%;竹山西南海域共鑒定游泳動物8種,魚類5種,占62.5%。
從年度變化來看,兩年度調查欽州灣游泳動物種類數目不變,其中各類群的種類變化也不大,但對照海域游泳動物種類數目有所減少,魚類種數在拖網漁獲中處優勢地位,蝦類第二,蟹類第三。
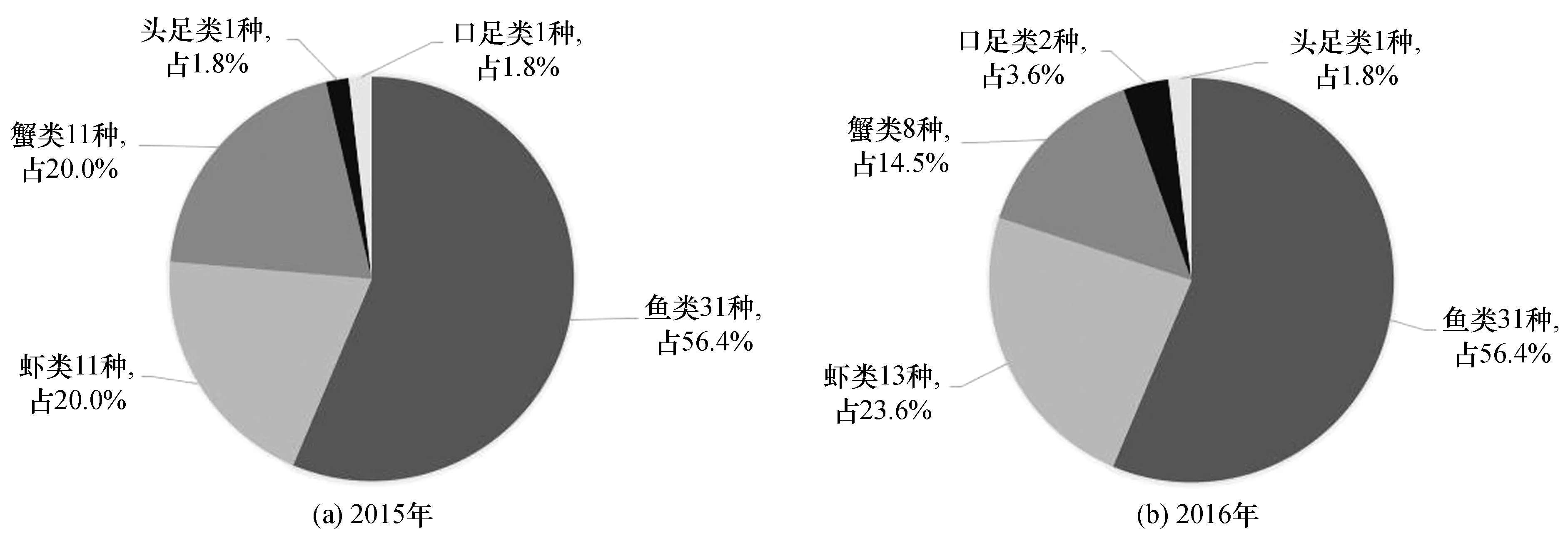
圖2 欽州灣調查游泳動物種類組成 Fig.2 Species composition of nektons in Qinzhou Bay
2.2 資源密度分布
2015年欽州灣游泳動物平均尾數密度為30 344ind./km2,其中蝦類平均達到13 381ind./km2,蟹類8 548ind./km2,魚類為4 757ind./km2,口足類為3 601ind./km2,頭足類僅57ind./km2,分別占總量的44.1%、28.2%、15.7%、11.9%和0.2%。游泳動物平均生物量為309.694kg/km2,其中蟹類達123.298kg/km2,魚類為89.881kg/km2,蝦類為58.083kg/km2,口足類為36.141kg/km2,頭足類僅2.290kg/km2,分別占總量的39.8%、29.0%、18.8%、11.7%和0.7%。對照站位企沙西南海域和竹山南部海域的密度分別為41 507ind./km2和106 011ind./km2,生物量分別為442.380kg/km2和732.160kg/km2,密度及生物量均高于欽州灣大多數站位水平,以魚類為絕對優勢類群。
2016年欽州灣游泳動物平均尾數密度為101 248ind./km2,其中魚類達到44 792ind./km2,蝦類為33 397ind./km2,蟹類18 493ind./km2,口足類為4 438ind./km2,頭足類僅127ind./km2,分別占總量的44.2%、33.0%、18.3%、4.4%、0.1%。游泳動物平均生物量為738.909kg/km2,其中魚類達326.081kg/km2,蟹類為234.856kg/km2,蝦類為150.274kg/km2,口足類為25.740kg/km2,頭足類僅1.958kg/km2,分別占總量的44.1%、31.8%、20.3%、3.5%、0.3%。企沙西南海域和竹山南部海域的密度分別為39 379ind./km2和25 457ind./km2,生物量分別為411.827kg/km2和72.941kg/km2,兩個站位的密度均低于欽州灣所有站位的密度水平,生物量水平偏低,其中魚類的絕對優勢地位顯著。
綜合兩年度調查數據來看,2016年欽州灣游泳動物平均尾數密度是2015年3倍多,平均生物量密度略大于2015年,而對照海域游泳動物的密度及生物量均有所降低。整個調查區以魚類資源密度最大。從資源密度分布來看(圖3),欽州灣灣外大部分站位資源密度均較高,但總體來說,欽州灣游泳動物資源密度分布極不均衡。
2.3 群落結構
由于生境狀況的差異,游泳生物優勢種組成具有明顯的季節變化特征,因此將游泳生物相對重要性指數IRI作為衡量的指標,來確定各游泳生物種類在群落中的重要性[12]。綜合出現率、漁獲尾數和重量,計算得出2015年欽州灣游泳動物的相對重要值IRI指數(表1),以IRI≥200為群落優勢種劃分的判斷指標,該次調查游泳動物優勢種為鈍齒蟳(Charybdis hellerii)、口蝦蛄(Oratosquilla oratoria)、周氏新對蝦(Metapenaeus joyneri)、強壯菱蟹(Parthenope validus)、近緣新對蝦(Metapenaeus affinis)、亨氏仿對蝦(Parapenaeopsis hungerfordi)、條紋叫姑魚(Johnius fasciatus)、斑鰶(Konosirus punctatus)和黃魟(Dasyatis bennetti),蟹類、口足類和蝦類等甲殼綱動物優勢度極為顯著。在企沙西南海域,游泳動物群落優勢種較為分散,鯻(Terapon theraps)、條馬鲾(Equulites rivulatus)、黑邊布氏鰏(Eubleekeria splendens)、真赤鯛(Pagrus major)、及達副葉鲹(Alepes djedaba)、斑條魣(Sphyraena jello)、遠海梭子蟹(Portunus pelagicus)、日本槍烏賊(Loligo japonica)、棕斑兔頭鲀(Lagocephalus spadiceus)、日本囊對蝦(Marsupenaeus japonicus)、亨氏仿對蝦、斑頭舌鰨(Cynoglossus puncticeps)和黑棘鯛(Acanthopagrus schlegelii)等13個種為優勢種群;竹山南部海域以條馬鲾、斑鰶、鹿斑仰口鲾(Secutor ruconius)、及達副葉鲹、刀額新對蝦(Metapenaeus ensis)、真赤鯛、日本囊對蝦、長鰭藍子魚(Siganus canaliculatus)、黃鰭棘鯛(Acanthopagrus latus)、二長棘犁齒鯛(Evynnis cardinalis)、李氏(Callionymus richardsoni)和多鱗鱚(Sillago sihama)等12個種優勢較為明顯。

圖3 2015、2016年游泳動物資源密度分布Fig.3 Density distributions of nekton resources in 2015 and 2016
表1 2015年主要游泳動物相對重要性指數
Tab.1IRIof important nekton species in 2015

種類出現率/%密度/(ind.·km-2)密度占比/%生物量/(kg·km-2)生物量占比/%IRI鈍齒蟳100.031 38617.24413.04022.233 946.77口蝦蛄100.021 60711.87216.84911.672 353.76周氏新對蝦66.746 85225.73164.1038.832 304.36強壯菱蟹66.714 7838.12241.63113.001 408.25近緣新對蝦83.317 1729.43125.4516.751 348.58亨氏仿對蝦100.010 8035.9319.3231.04697.37條紋叫姑魚66.77 0513.87115.8556.23673.83斑鰶33.37 0513.87125.8026.77354.76黃魟33.32270.12116.4226.27213.01
同比,2016年調查中顯示欽州灣海區游泳動物優勢種為9種(表2),分別為皮氏叫姑魚(Johniusbelengerii)、鈍齒蟳、亨氏仿對蝦、近緣新對蝦、日本囊對蝦、香港蟳(Charybdishongkongensis)、周氏新對蝦、口蝦蛄和伍氏平蝦蛄(Erugosquillawoodmasoni),蝦類和蟹類等甲殼綱動物優勢度極為顯著。在企沙西南海域,游泳動物群落優勢種群則較為集中,條馬鲾、日本囊對蝦、亨氏仿對蝦、尾紋雙邊魚(Ambassisurotaenia)、鹿斑仰口鲾、李氏、鲬、真赤鯛、及達副葉鲹、斑頭舌鰨、長鉤須鰨(Paraplagusiabilineata)和高體斑鮃(Pseudorhombuselevatus)等12個種均為優勢種群;竹山南部海域優勢種為刀額新對蝦、日本囊對蝦、康氏側帶小公魚(Stolephoruscommersonnii)、短吻鲾(Leiognathusbrevirostris)、條馬鲾、長鰭藍子魚和斑條魣。
綜合來看,鈍齒蟳、近緣新對蝦、周氏新對蝦、亨氏仿對蝦和口蝦蛄為欽州灣海域兩年共有的優勢種,有較高且較為穩定的優勢地位,而對照海域居前的優勢種以魚類為主,其他類群不突出。因此,本研究海區漁獲優勢種以小型魚類、蝦類和蟹類為主,其大多生命周期較短,生長速度快,屬沿岸性種類,且不少種類當年春季繁殖,生長至夏、秋季便可成為捕撈對象。
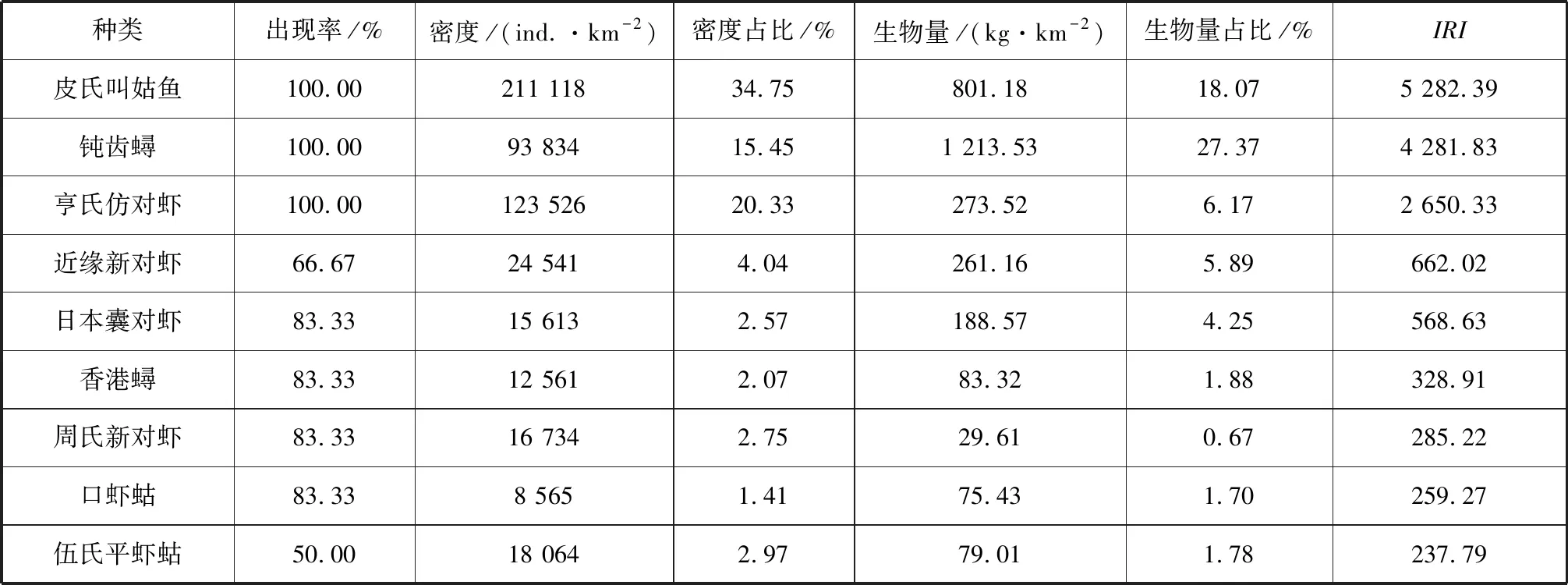
表2 2016年主要游泳動物相對重要性指數Tab.2 IRI of important nekton species in 2016
2.4 群落結構特征
2015年調查海域的游泳動物Shannon-Wiener指數在1.679~3.258范圍內[圖4(a)],平均2.823;Margalef豐富度指數介于1.633~3.611之間,均值為2.565;Pielou均勻度指數范圍在0.441~0.770,平均為0.650。
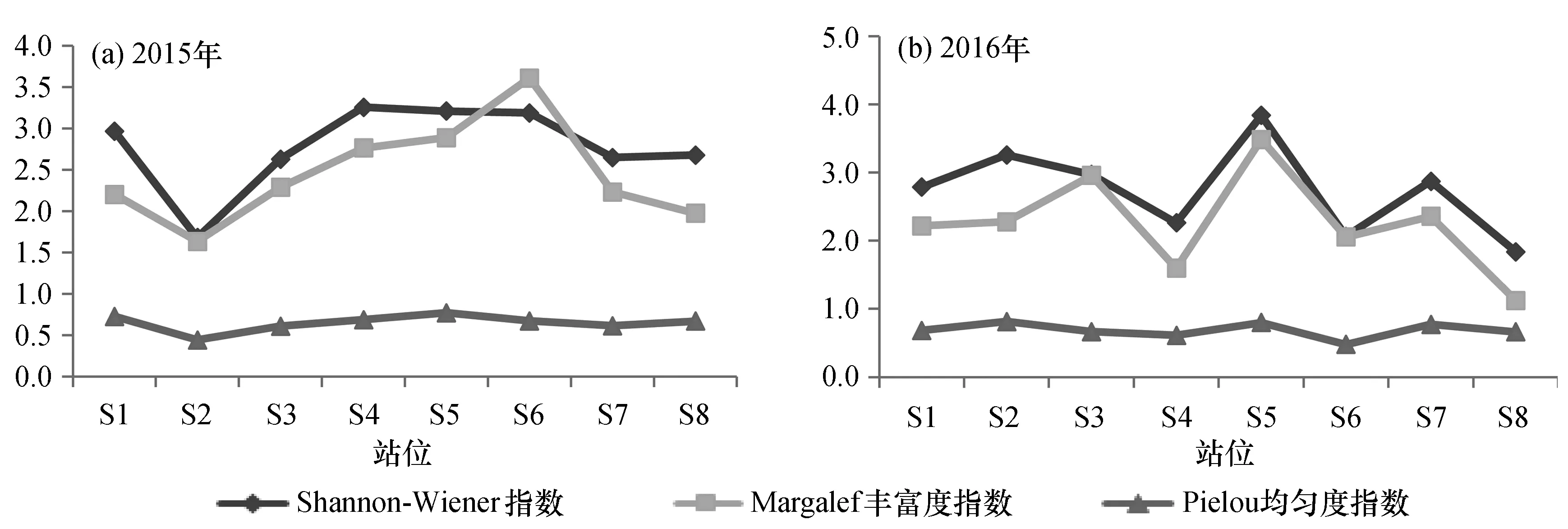
圖4 游泳動物多樣性指數變化Fig.4 Changes of nekton diversity indexes
2016年調查海域的游泳動物Shannon-Wiener指數在2.063~3.837范圍內[圖4(b)],平均2.611;豐富度指數介于1.595~3.485之間,均值為2.112;Pielou均勻度指數范圍在0.477~0.815,平均為0.666。
綜合兩年度調查來看,Margalef豐富度指數的波動范圍最大,與Shannon-Wiener指數相同,其中2015年調查的平均值高于2016年,變化范圍也略小于2016年,而Pielou均勻度指數則表現相反。從整體來看,欽州灣海域靠外站位的種類多樣性程度較高,企沙西南海域和竹山南部海域游泳動物多樣性稍低于欽州灣監測區的靠外站位,但稍高于靠內站位。
2.5 區系組成特征與生態類型
調查水域地處亞熱帶,從適溫性看,該海域魚類區系組成全部為暖水性和暖溫性種,沒有出現冷水性種類。其中,兩年捕獲的49種魚類漁獲物中,以暖水性種類占絕對優勢,為34種,占69.4%,暖溫性種為15種,占30.6%;這表明本次調查海域的魚類區系具有熱帶和亞熱帶的特征。從生態類型的分布看,底層魚類最多,有32種,占65.3%;其次為中上層魚類,有11種,占22.4%,中下層魚類種類有5種,占10.2%,近底層魚類1種。蝦蟹類、頭足類和口足類種類基本以暖水性和暖溫性種類為主,其中暖水性種類,蝦類13種,蟹類12種,頭足1種,多數種類棲息習性為河口型、近岸型和淺海型。
總的來看,欽州灣漁獲種類區系組成以暖水性和暖溫性為主,甲殼類主要以廣溫廣鹽性種類為主,棲息習性以底層種類占絕對優勢。
2.6 討論
2.6.1 種類組成特點 游泳動物種類組成受海洋環境多因子的長期影響[16],其中,鹽度、水溫和底質的影響最為重要。綜合兩年調查發現,欽州灣游泳動物種類組成以魚類和甲殼類為主,頭足類及口足類所占比例小。結合同年該海域其他調查顯示,2016年漁獲數量有較大上升,可能從“厄爾尼諾現象促進海洋生物繁榮”的推斷得到支撐。李菲萍(2011)等分析得出海洋捕撈產量增長率隨著厄爾尼諾的強度增加而增加,也隨著厄爾尼諾持續季度的增加而增加[17],這也與吳志強(2000)等分析閩南—臺灣淺灘漁場中上層魚類資源狀況時提出魚類產量的波動可能與厄爾尼諾現象強度及其持續時間有關的觀點[18]一致。同時,李耀先(2001)等認為厄爾尼諾發生時,哈得萊環流的異常增強造成了副熱帶高壓的增強,從而使得廣西前汛期降水偏多[19],而李寇軍(2007)等的研究結果發現北部灣漁業資源量的波動受沿海陸地降雨量年際變化的影響很大[20]。對比相鄰海域調查來看,企沙南部海域及竹山南部海域各1個站位調查的游泳動物物種數相對較少,資源密度較低。近年來,沿海工業化城鎮化進程加快,臨海工業項目紛紛落成,回顧該海區歷史調查數據,本次調查游泳動物的相對資源密度較2009年欽州灣夏季(8月)調查結果偏低。同時,根據2009—2010年北部灣東北部海域游泳生物資源夏季的調查數據進行對比發現,欽州灣水域游泳動物物種相對較少,2015年資源密度較其偏低,2016年小幅回升后則高于其平均生物量水平(622.514 kg/km2)[21]。游泳動物由于游動能力較強,采樣的偶然性及站位設置和采樣方法的不同很可能造成調查結果的差異[16],但欽州灣海域漁業資源呈現衰退的趨勢是有據可依的。
在適溫性上看,欽州灣漁獲以暖水性和暖溫性為絕對優勢。這可能是欽州灣夏季水溫南高北低,冬季水溫差異較小,水溫垂向梯度小,適于對溫度要求范圍比較廣的游泳動物棲息。從生態類型看,內灣、沿岸性種類絕大多數屬地方性種群,分布范圍很廣,大多數種類在北部灣沿海均有分布。整個生命過程的主要階段包括索餌生長和生殖活動等,均在沿岸、內灣水域度過,不作長距離洄游。
2.6.2 種群結構變化 優勢種在生態系統中占據重要地位,其變化能影響整個群落的數量變化和能量流動,同時也是海洋生態系統中的關鍵因子[22]。據王雪輝(2010)等曾將北部灣按空間劃分為5個群落進行分析表明,北部灣東北部沿岸海域的特征種類以鲾類、麗葉鲹(Caranxkalla)和二長棘犁齒鯛幼魚等小型魚類為主;海南島西岸海域為典型的巖礁魚類群落,以巖礁魚類為該群落全年的優勢種;北部灣中南部海域分布的是北部灣魚類的主要群落,以發光鯛(Acropomajaponicum)、大頭白姑魚(Pennahiamacrocephalus)、黃斑鲾(Leiognathusbindus)和竹莢魚(Trachurusjaponicus)為主;龍尾島附近海域魚類分布范圍最小,特征種較為復雜,沒有周年優勢種;瓊州海峽西側海域僅在春季形成以黃斑鲾、帶魚(Trichiurusjaponicas)幼魚、二長棘鯛(即二長棘犁齒鯛)和黃帶緋鯉(Upeneussulphureus)等為特征種的魚類群落[23]。本次調查區域位于北部灣北部沿岸海域,調查發現該海域優勢種類均以小型魚類、蝦類和蟹類為主,種類更替現象明顯,結合調查分析,海域深度影響海域水體環境,不同環境因素對魚類分布有明顯的限制作用,因此在不同的水系和水深往往會分布不同的種類,從而形成了魚類群落的優勢特征[20]。
2.6.3 群落多樣性比較 南海魚類多樣性指數明顯高于東海和黃海。這種現象與各海域所處的地理位置有密切關系,低緯度海域的物種多樣性比高緯度海域高[24]。由于受季節限制,兩年調查均為夏季,從群落多樣性指數來看,總體呈現南高北低的特點,兩個年度并沒有太大差別,說明調查海區游泳動物群落大體上維持輕度至中度擾動的格局。群落多樣性指數與對照站位相比,兩個對照海域游泳動物多樣性稍低于欽州灣監測區的靠外站位,但稍高于靠內站位;與北部灣海域和南海大陸架相比,低于北部灣東北部夏季的平均多樣性水平(3.36)[24],并且低于同季節南海北部大陸架海域的平均多樣性水平(3.65)[25],總體上處于較低水平。而夏季是很多游泳動物產卵的季節,漁獲物中幼魚比例會增高,但由于過度捕撈,違規拖網作業,已嚴重影響了游泳動物季節更替的過程,對幼魚幼蝦等生物資源破壞極大,使得漁獲量降低,多樣性指數降低;秋季來臨,游泳動物經歷了兩季的生長和休漁期的休生養息使得漁獲量增加,但同樣也隨著休漁期的結束,一些價值較高的經濟種被有選擇性地捕撈,造成生物種類比例改變,再隨著捕撈壓力增大,最終也將導致多樣性指數降低。因此,應加強欽州灣生態環境、生物資源繁殖保護,依據游泳生物群落的季節變化規律,控制過度捕撈,加強敏感典型海洋生態系統及優勢種群監測,促使其漁業資源總體價值和游泳動物多樣性的提高。
2.6.4 人為因素影響 隨著高強度的經濟開發建設和大型臨海工業園的密集布局,極大地改變了欽州灣海域的自然屬性。其中,防城港核電廠圍填海建設永久改變欽州灣海域水動力環境,長期影響水體交換能力及環境質量。核電廠排水明渠從廠區延伸至欽州灣西航道邊緣,溫排水排放在水動力的作用下使環境水體水溫有不同程度的上升,其近區溫升呈伴岸窄帶型分布,夏季熱效應對漁業資源的影響比其他季節更為明顯。魚類多為變溫動物,溫度變化對魚類生命活動過程影響很大;同時溫度過高還會造成甲殼類死亡。根據2015、2016年調查水域各站位監測數據,夏季海水表層溫度較高,但溫排水引起的溫升對魚類生長的影響僅局限于欽州灣西側核電廠溫排水排放口附近3℃溫升海域,且魚類本身可以感受到極其細微的水溫變化,對超過其適溫范圍的水體具有回避反應,能進行適溫洄游。
3 結論
海洋生物物種的生態健康程度,是評價海洋承載力及生態環境風險等級的最重要指標之一。綜合2015—2016年欽州灣拖網調查數據分析,雖然調查海區漁獲物種類和現存資源較歷史數據有所衰減,游泳動物群落呈現輕度至中度擾動的格局,但總體上暫無發現顯著影響。因此,保障后續連續定位監測力度,緊密跟蹤自然變化和核電站溫排水影響產生的生物累積效應,加強水域生態環境的監測保護,合理開發利用海洋資源,對促進欽州灣海域漁業資源可持續發展至關重要。
