基于SRAP標記的紫溪山華山松種子園無性系遺傳多樣性分析
2020-05-26 14:13:07劉成徐劍羅正平董云祥董章宏瞿紹宏李顯煌辛培堯
廣西植物 2020年4期
劉成 徐劍 羅正平 董云祥 董章宏 瞿紹宏 李顯煌 辛培堯


摘 要: 為明確云南楚雄市紫溪山華山松種子園內不同種源無性系間的遺傳背景,該研究收集了園內6個種源的60個華山松無性系單株針葉,采用改良CTAB法提取其總DNA,并利用SRAP分子標記對其進行遺傳多樣性分析。結果表明:(1)從100對引物組合中共篩選出15對具有多態性的SRAP引物,經SRAP-PCR擴增后,共獲得出194個位點,多態位點百分率(PPB)為85.05 %,Nei′s 基因多樣性指數(H)為0.233 7,Shannons信息指數(I)為0.341 9,種源間的遺傳分化系數(GST)為 0. 355 5。(2)華山松6個種源遺傳多樣性較高,且遺傳變異主要存在于華山松種源內,種源地會澤(HZ)與巍山(WS)種源的遺傳距離最近(D=0.050 1),會澤(HZ)與宜良(YL)種源的遺傳距離最遠(D=0.361 8)。(3)聚類分析顯示將6個華山松種源一共聚為3類:會澤(HZ)和巍山(WS)種源聚為一類;楚雄(CX)、南華(NH)和宜良(YL)種源聚為一類;騰沖(TC)種源單獨為一類。綜合上述結果顯示,紫溪山華山松種子園內無性系間的遺傳分化處于較高水平,為華山松雜交育種時親本的選配及種質資源的評價提供了分子水平的理論依據。
關鍵詞: 華山松, 種子園, 無性系, SRAP, 遺傳多樣性
中圖分類號: Q943 ?文獻標識碼: A
文章編號: 1000-3142(2020)04-0462-09
Abstract: In order to clear genetic background of different provenances Pinus armandii colnes from seed orchard in Zixi Mountain of Chuxiong City, Yunnan Province, the needles of 60 clones were taken from six provenances which was used to extract total DNA by modified CTAB method, and the genetic diversity was analyzed by SRAP molecular markers. The results were as follows: (1) Fifteen pairs of polymorphic SRAP primers were screened from 100 pairs of primer combinations. After SRAP-PCR amplification, a total of 194 loci were obtained, and the percentage of polymorphic loci PPB=85.05%, Nei′s gene diversity index H=0.233 7, Shannons information index I=0.341 9, genetic differentiation coefficient between population GST=0.355 5. (2) The genetic diversity of 6 provenances of P. armandii was high, and the genetic variation was mainly existed within provenances of P. armandii. The genetic distance between the Huize (HZ) provenance and the Weishan (WS) provenance was the closest (D=0.050 1), that the Huize (HZ) provenance and Yiliang (YL) provenance was the farthest (D =0.361 8). (3) The cluster analysis showed that six P. armandii provenances there were classified into three categories: Huize (HZ) and Weishan (WS) provenances were in one category; Chuxiong (CX), Nanhua (NH) and Yiliang (YL) provenances were in the second category; Tengchong (TC) provenance was a separate category. The results indicate that genetic differentiation in P. armandii clones from seed orchard in Zixi Mountain of Chuxiong City has a high level, and it can provide theoretical information and practical guidance for parent selections during the cross breeding and evaluation of germplasm resources of P. armandii as well.
Key words: Pinus armandii, seed orchard, clone, SRAP, genetic diversity
華山松(Pinus armandii)是松科(Pinaceae)松屬(Pinus)單維管束亞屬的植物(陳有民,1990; 袁紅寧和陳文輝,2010),是我國特有的五針松高大喬木,同時也是高海拔地區重要的造林樹種之一(趙楊等,2003;辛培堯等,2010)。20世紀70年代起,云南和貴州等地相繼對華山松進行遺傳改良,各自營建了數個優良無性系初級種子園,之后對其進行了遺傳測定、優良家系選擇及輔助授粉等工作(王秀榮等,2007;翟思萬等,2007)。紫溪山華山松無性系種子園位于楚雄市紫溪山,為國家級林木良種基地。由于缺乏對園內種質的不斷改良,該種子園仍然處于建園初期低級階段,繼而出現病蟲害嚴重及生產力低下的問題,因此,急需對園內種質進行改良和創新。已有實踐證實,雜交育種是迄今為止對植物不良性狀進行遺傳改良的最為有效的方法之一。多年來,華山松的雜交育種工作較為滯后,雜交親本的選配多以形態及種源地為主要參考依據,不能明確所收集的種質資源間遺傳多樣性的豐度及親緣關系的遠近,從而影響改良的程度及效果。因此,在種子園內進行華山松雜交育種時,需有針對性地明確園內種質的遺傳多樣性程度及各種質間的親緣關系顯得尤為重要。隨著分子生物學研究的不斷深入及相關領域技術水平的進步,華山松遺傳多樣性的檢測方法不斷得到完善。例如:趙楊等(2012)利用ISSR標記對貴州平壩華山松無性系種子園內的85個無性系進行遺傳多樣性分析后發現,其遺傳變異主要在種源內;祝娟(2016)利用SSR分子標記對華山松52個自然分布群體進行遺傳多樣性分析,結果表明遺傳變異主要存在于個體內,且其遺傳多樣性水平較高;朱曉丹(2006)利用RAPD分子標記對紫溪山華山松種子園內的部分無性系進行了遺傳多樣性研究,結果表明各種源之間具有較大的遺傳差異,且遺傳多樣性較豐富。但由于RAPD(Random Amplified Polymorphic DNA)標記相對較早,且與新型分子標記比較,存在一定的缺陷(阮楨媛等,2016),其研究結果可能存在一定的局限性。上述相關研究為華山松的遺傳改良及種質資源的分子評價提供了重要的理論基礎。
在眾多的分子標記中,SRAP(Sequence-related Amplified Polymorphism)憑借其多態性高、操作簡單、易于分離測序,且多數標記在相關實驗樣本基因組中相對均勻分布的特點(Li et al., 2001;Tilman et al., 2006),目前在較多的林木遺傳多樣性分析中得到廣泛應用(張冬東,2008;向暉等,2016;陳慧等,2017;孟慧等,2018;張安世等,2018;姜武等,2019)。趙楊等(2012)對貴州平壩華山松種子園無性系進行了SRAP-PCR反應體系的優化,但未對其遺傳多樣性進行具體分析。目前,對華山松遺傳多樣性進行SRAP分析的相關研究,國內外鮮見報道。
針對云南省楚雄市紫溪山華山松種子園的生產現狀,實驗選取6個種源共60個華山松優良無性系,開展了以雜交育種及種質資源評價為最終目的的華山松SRAP遺傳標記分析,研究結果可為該種子園內華山松種質資源的評價以及雜交育種時親本的選配提供分子水平的理論與實踐指導。
1 材料與方法
1.1 材料來源
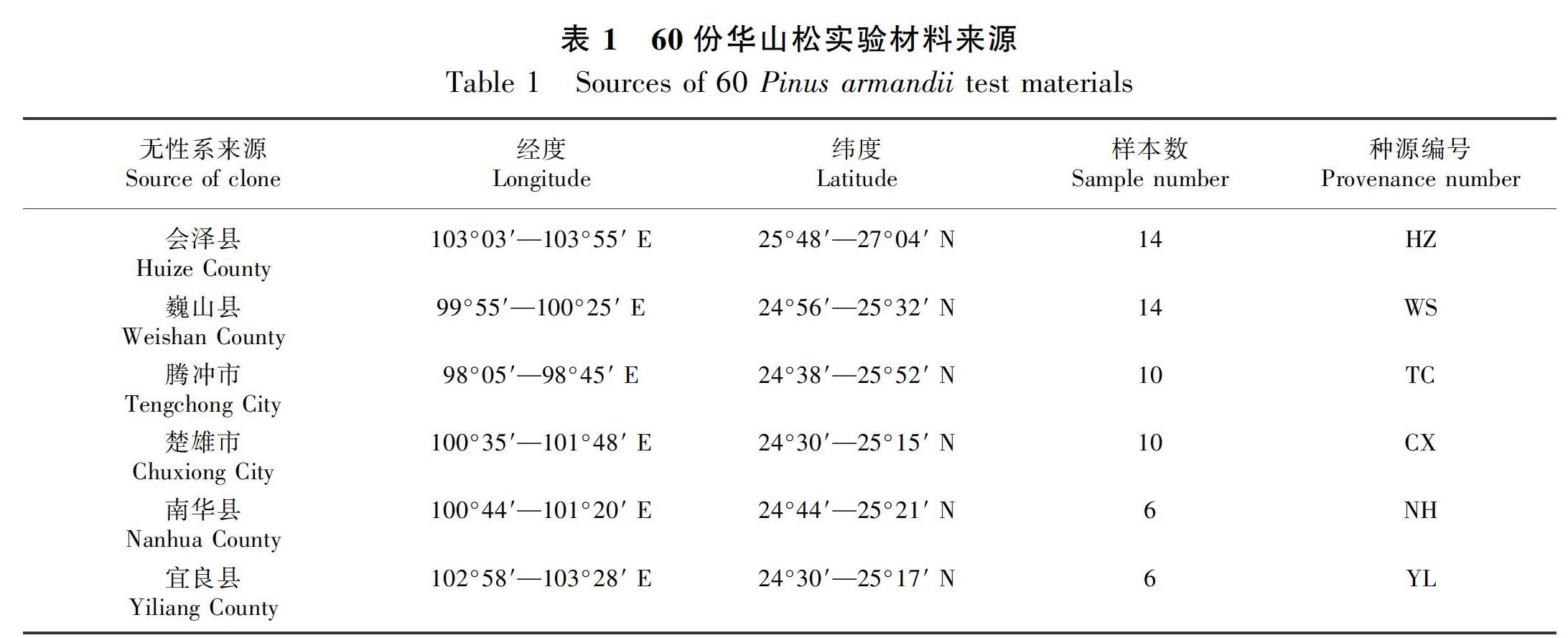
材料采集于云南省楚雄市紫溪山華山松種子園內60個無病蟲害的不同無性系新鮮幼嫩針葉,分別按無性系單株編號后裝入自封袋,帶回實驗室后置于 -80 ℃冰箱,備用。所有材料均來自云南省內的6個種源(表1)。
1.2 總DNA提取
取4~5根松針,采用改良的CTAB法,提取華山松基因組DNA(謝冬梅等,2014)。加入50 μL TE buffer稀釋所提取的DNA至50 ng·μL-1,然后用1%的瓊脂糖凝膠電泳進行檢測,并保存于-20 ℃冰箱中。
1.3 SRAP-PCR擴增
實驗擴增所用引物序列來自公開發表的相關報道(李正鵬等,2011;蔡元保等,2014)。選擇正向和反向引物各10條,隨機組合后獲得100對引物(表2)。經初篩和復篩后成功篩選出15對多態性較好且條帶清晰的SRAP引物(表3)。華山松SRAP-PCR 25 μL反應體系:DNA 模板1 μL、Green Taq Mix12.5 μL、ddH2O 9.5 μL、正向和反向引物各1 μL。其PCR擴增反應程序:94 ℃預變性5 min;94 ℃變性1 min,37 ℃退火1 min,72 ℃延伸1 min,5個循環;94 ℃變性1 min,53 ℃退火1 min,72 ℃延伸1 min,30個循環;72 ℃延伸10 min;4 ℃保存。
1.4 聚丙烯酰胺凝膠電泳
吸取6 μL PCR擴增產物和2 μL 6×Loading buffer混合后,加入到8%聚丙烯酰氨凝膠孔中進行電泳檢測,電壓設為230 V,電泳時間為150 min。電泳結束后進行凝膠的固定、染色和顯影,拍照保存檢測結果。
1.5 數據處理
將擴增得到清晰可辨的多態性條帶, 作為參考帶型,根據實驗得到的聚丙烯凝膠電泳圖,同一位點清晰的條帶,記“1”;無帶或者弱帶記“0”,模糊不清的不予計數,從而構成(0,1)矩陣,調整格式以符合軟件分析要求。
利用Excel表對60株華山松進行多態位點數、多態性百分比統計計算;通過POPGENE version 1. 32分析軟件進行遺傳多樣性分析[觀測等位基因(Na)、有效等位基因數(Ne)、基因多樣度(H)等參數];利用NTSYS 2.10軟件對6個種源的華山松進行聚類,形成聚類圖。
2 結果與分析
2.1 華山松基因組DNA提取結果
將華山松基因組DNA用1%的瓊脂糖凝膠檢測,部分結果如圖1所示,DNA條帶清晰且無拖帶和彌散現象,可滿足后續實驗的要求。
2.2 SRAP-PCR擴增結果
用15對具有多態性的引物對6個種源共60株不同華山松無性系進行PCR擴增反應,共擴增出165個多態性位點,其結果列于表4。
從表4可以看出,在各種源間,華山松多態位點百分率(PPB)為 85.05%;在各種源內,各個種源的PPB為41.24%~81.96%,平均值為59.37%。PPB最低的是宜良(YL)種源,為41.24%;PPB最高的是騰沖(TC)種源,為81.96%。把華山松6個種源根據PPB大小進行排序:騰沖(TC)>會澤(HZ)>巍山(WS)>楚雄(CX)>南華(NH)>宜良(YL)。
2.3 遺傳多樣性分析
華山松6個不同種源的Nei′ s基因多樣性(H)范圍為0.187 3~0.323 4,楚雄(CX)的最低,為0.187 3,騰沖(TC)的最高,為0.323 4。Shannons信息指數(I)范圍為0.277 0~0.471 6,楚雄(CX)的最低,為0.277 0,騰沖(TC)的最高,為0.471 6。等位基因數(Na)最高的是騰沖(TC),為1.819 6,最低的是宜良(YL),為1.142 4,有效等位基因數(Ne)最高的是騰沖(TC),為1.580 9,最低的是宜良(YL),為1.269 1。6個種源的H和I均表明,各種源的遺傳變異從大到小依次為騰沖(TC)>會澤(HZ)>巍山(WS)>南華(NH)>宜良(YL)>楚雄(CX)(表5)。
2.4 遺傳分化分析
由Nei′s指數估算可知,6個華山松種源的總基因多樣度(Ht)為0.342 0,種源內基因多樣性(Hs)為0.220 4。遺傳分化系數(GST)為0.355 5,說明在華山松種源間有35.55%的遺傳變異, 而在華山松種源內則有64.45%的遺傳分化,體現了華山松種源內的遺傳分化遠大于華山松種源間的遺傳分化。華山松各種源間的基因流(Nm)為0.906 5<1,表明華山松各種源間的基因交流較少。
2.5 遺傳相似度與遺傳距離分析
為了進一步分析華山松種源間的遺傳關系,對華山松種源的遺傳相似度和遺傳距離進行分析,其結果如表6所示。由表6可知,華山松6個種源的遺傳相似度在0.696 4~0.951 1之間,平均值為0.814 1;遺傳距離在0.050 1~0.361 8之間,平均值為0. 212 3。其中,遺傳距離最大的是宜良(YL)和會澤(HZ)(D=0.361 8),而它們之間的遺傳相似度相對較小(Gs=0.696 4),說明宜良(YL)和會澤(HZ)之間的親緣關系最遠;會澤(HZ)與巍山(WS)之間的遺傳距離最小(D=0.050 1),它們之間的遺傳相似度最大(Gs =0.951 1),這表明會澤(HZ)與巍山(WS)的遺傳差異較小,親緣關系最近。
2.6 群體間遺傳關系聚類分析
根據6個華山松種源遺傳相似度,對華山松種源進行聚類分析,其結果如圖2所示。以0.87為閾值,華山松6個種源被分為三個類群。其中,會澤(HZ)和巍山(WS)種源分為一類;楚雄(CX)、南華(NH)、宜良(YL)種源被聚為一類;騰沖(TC)種源單獨聚為一類。
3 討論與結論
遺傳分化系數(GST)是衡量種群基因分化程度的重要指標,能較好地反映群體的遺傳分化水平。按照GST的大小可分為4種情況:GST在0~0.05之間時,種群的遺傳分化水平基本不存在;GST在0.05~0.15之間時,種群遺傳分化水平為中等水平;GST在0.15~0.25之間時,種群遺傳分化水平較高;GST大于0.25時,種群遺傳分化水平相當高(孫文婷等,2016;蔣謙才等,2018)。本實驗結果表明,華山松6個種源的總基因多樣度(Ht)為0.342 0,種源內基因多樣性(Hs)為0.220 4,GST為0.355 5,其遺傳變異達35.55%,在華山松種源內有64.45%的遺傳分化,體現了華山松種源內的遺傳分化遠大于華山松種源間的遺傳分化,說明這6個種源的遺傳分化處于較高的水平。這和Wheeler & Jech(1992)研究發現種子園不會因為群體數量有限可能導致遺傳多樣性較天然種群低相一致。此外,朱曉丹(2006)曾對楚雄華山松種子園4個種源不同無性系進行RAPD遺傳多樣性分析,其PPB為71.42%,Ne為1.364 9,H為0.140 4,I為0.219 3,說明這4個種源64個個體間的遺傳差異較大,該種子園遺傳多樣性較豐富。本實驗利用與朱曉丹(2006)所研究的同一種子園內材料進行SRAP遺傳多樣性分析,得到多態位點百分率(PPB)為85.05%,Ne為1.386 6,H為0.233 7,I為0.341 9,各項參數均有顯著提高,且主要遺傳變異也存在于種源內。由于RAPD標記本身具有重復性低,擴增產物不穩定,特異性不強等缺點,該實驗結果可能有一定的局限性,而SRAP標記,多態性強、標記分布均勻且比較穩定(周延清,2008)。因此,相比RAPD標記可以獲得更加豐富且可靠的遺傳變異信息,同時,種子園內各單株間不同程度的遺傳變異,也有利于在園內更加方便地就地進行華山松的遺傳改良工作。
在生物自然進化中,基因流(gene flow)有助于提高物種種源的遺傳多樣性水平,防止種群分化(Ellstrand et al., 1993)。Widen & Svensson(1992)統計了124種植物的基因流(Nm)認為,遺傳漂變對種群分化的影響較大。而Wright(1950)認為當基因流Nm<1時,遺傳漂變是導致種群之間存在明顯遺傳分化的主要原因。有學者還指出,如果植物種群的基因流(Nm>0.5)從原來的較高水平,到后來呈現較大下降趨勢,則猜想原因可能是植物生存的環境斑塊化、傳粉的減少、自然選擇作用的加強等降低了基因流(楊章旗等,2014)。本研究結果表明,紫溪山華山松種子園內無性系Nm= 0.906 5<1,說明供試華山松6個種群間基因交流水平相對較低,華山松的遺傳變異主要存在于種源內,即種源內的不同優良單株之間的遺傳差異較大。那么,在雜交育種時,可在種源內進行親本的選配,這樣不僅有利用于提高其雜交親合性,同時還可豐富雜種后代的基因型,從而更有利于人工選擇。
在進行植物遺傳多樣性分析時,遺傳距離是一個用來反映物種和種源間遺傳變異水平及遺傳分化程度的重要指標,可為雜種優勢提供基本遺傳參數(睢鑫洲等,2017)。利用SRAP分子標記技術對6個華山松種源共60個單株樣本進行聚類分析發現:在閾值為0.95時,可以分成五類,其中,會澤(HZ)和巍山(WS)種源最先聚為一類,楚雄(CX)、南華(NH)、騰沖(TC)、宜良(YL)4個種源各自被聚為一類,而在閾值為0.87時,6個種源被聚為三類,其中騰沖(TC)單獨聚為一類;會澤(HZ)和巍山(WS)種源分為一類;南華(NH)、宜良(YL)和楚雄(CX)三個種源被聚為一類,說明會澤(HZ)和巍山(WS)之間的親緣關系較近。由于種源間的親緣關系與其分布的地理種源存在相關性。一般認為,地理位置相近的種源其遺傳基礎也較為接近。實驗結果表明,曲靖會澤(HZ)種源與大理巍山(WS)種源就被聚為一類,顯然這種聚類與物種地域性的分布不相符。鄢家俊等(2009)研究老芒麥遺傳多樣性時也被發現這一現象。而對裸果木利用SCoT標記進行遺傳多樣性分析時發現,其種群地理距離與遺傳距離也不存在顯著相關性(狄林楠,2018)。研究認為,植物群體間當前的遺傳構成不僅與目前的分布地理距離有關,還可能和該植物的進化歷史和各種群受人為干擾程度等各種因素有關(劉柳,2012;宋春鳳等,2014;狄林楠,2018)。因此,在進行植物雜交育種中,以目前兩親本地理位置的遠近為選配親本的主要標準,顯然存在較大的局限性。在今后的育種工作中,應以DNA水平分析的結果為主要依據,結合形態,地理位置等綜合評判,最后確定較為理想的親本組合。通過雜交,從其子代中進行選擇,有望獲得更多的具有優異性狀(如抗病蟲害能力的提高、速生性等)的優良個體,再通過無性擴繁的手段對其優良性狀加以固定,最終獲得良種,繼而在生產上大面積推廣。
參考文獻:
CAI YB, YANG XY, CHEN HJ, et al., 2014. Analysis of genetic diversity of Papaya germplasm by SRAP combined with SCoT marker [J]. J Plant Gene Resour, 15(2): 292-298. [蔡元保, 楊祥燕, 陳豪軍, 等, 2014. SRAP結合SCoT標記分析番木瓜種質的遺傳多樣性 [J]. 植物遺傳資源學報, 15(2): 292-298.]
CHEN H, QIE HL, HUANG YH, et al., 2017. Genetic diversity analysis of 66 Chinese Bayberry germplasm resources with SRAP [J]. S Chin Fruits, 46(4): 64-67. [陳慧, 郄紅麗, 黃穎宏, 等, 2017. 66份楊梅種質SRAP標記遺傳多樣性分析 [J]. 中國南方果樹, 46(4): 64-67.]
CHEN YM, 1990. The landscape dendrology [M]. Beijing: China Forestry Publishing House. [陳有民, 1990. 園林樹木學. [M]. 北京: 中國林業出版社.]
DI LN, 2018. Genetic diversity of Gymnocarpos przewalskii based on SCoT markers [J]. Bull Bot Res, 38(5): 725-732. [狄林楠, 2018. 基于SCoT分子標記的裸果木遺傳多樣性分析 [J]. 植物研究, 38(5):725-732.]
ELLSTRAND NC, ELAM DR, 1993. Population genetic consequences of small population size: implications for plant conservation [J]. Ann Rev Ecol Sys, 24(1): 217-242.
JIANG QC, TAN ZJ, LIAO HB, et al., 2018. Genetic diversity of Aquilaria sinensis in Wuguishan, Zhongshan City, Guangdong Province [J]. Guihaia, 38(6): 804-811. [蔣謙才, 譚宗健, 廖浩斌, 等, 2018. 廣東省中山市五桂山土沉香遺傳多樣性 [J]. 廣西植物, 38(6): 804-811.]
JIANG W, WU ZG, TAO ZM, et al., 2019. Genetic diversity analysis of Gardenia jasminoides based on ISSR and SRAP molecular markers [J]. Chin Trad Herb Drugs, 50(2): 510-516. [姜武, 吳志剛, 陶正明, 等, 2019. 基于ISSR和SRAP標記的梔子種質遺傳多樣性研究 [J]. 中草藥, 50(2): 510-516.]
LI G, QUIROS CF, 2001. Sequence-related amplified polymorphism (SRAP), a new marker system based on a simple PCR reaction: its application to mapping and gene tagging in Brassica [J]. Theoretical & Appl Gene, 103(2-3): 455-461.
LI ZP, LI TC, FAN HH, et al., 2011. Genetic diversity analysis of Papaya resources by SRAP and SCoT combination [J]. Acta Laser Biol Sin, 20(2): 236-244. [李正鵬, 李廷春, 樊洪泓, 等, 2011. 棉花遺傳多樣性SCoT和SRAP標記的研究及比較分析 [J]. 激光生物學報, 20(2): 236-244.]
LIU L, 2012. Population evolution history and speciation study of Pinus armandii and its three affinities [D]. Beijing: Graduate University of Chinese Academy of Sciences. [劉柳, 2012. 華山松(Pinus armandii)及其近緣種的群體進化歷史與物種形成研究 [D]. 北京:中國科學院研究生院.]
MENG H, CHEN B, YANG Y, 2018. Analysis of genetic diversity of Dalbergia odorifera growing in Hainan by SRAP [J]. Chin J Trop Agric, 38(12): 37-42. [孟慧, 陳波, 楊云, 2018. SRAP技術分析海南降香檀遺傳多樣性 [J]. 熱帶農業科學, 38(12): 37-42.]
RUAN ZY, WANG BY, OUYANG ZQ, et al., 2016. Characterization of microsatellites in genome of Pinus squamata, a critically endangered species in the world [J]. Bull Bot Res, 36(5):775-781. [阮楨媛, 王兵益, 歐陽志勤, 等, 2016. 極度瀕危植物巧家五針松基因組微衛星特征分析 [J]. 植物研究, 36(5):775-781.]
SONG CF, LIU QX, ZHOU YF, et al., 2014. Genetic diversity analysis of Glehnia littoralis (Apiaceae) revealed by SRAP [J]. Guihaia, 34(1): 15-18. [宋春鳳, 劉啟新, 周義峰, 等, 2014. 珊瑚菜居群遺傳多樣性的SRAP分析 [J]. 廣西植物, 34(1):15-18.]
SUI XZ, ZHENG Y, QIU YP, et al., 2017. Genetic diversity of 4 Betula alnoides [J]. J SW For Univ (Nat Sci Ed), 37(3): 21-25. [睢鑫洲, 鄭毅, 仇玉萍, 等, 2017. 4個西南樺品系的遺傳多樣性分析 [J]. 西南林業大學學報(自然科學版), 37(3): 21-25.]
SUN WT, YU DD, DONG ML, et al., 2016. Genetic diversity of control-pollinated progenies in seed orchard of Larix gmelinii var. principis-rupprechtii Mayr. [J]. Acta Bot Boreal-Occident Sin, 36(8): 1662-1670. [孫文婷, 于大德, 董明亮, 等, 2016. 華北落葉松種子園控制授粉子代遺傳多樣性分析 [J]. 西北植物學報, 36(8): 1662-1670.]
TILMAN D, REICH PB, KNOPS JM, 2006. Biodiversity and ecosystem stability in a decade-long grassland experiment [J]. Nature, 441(4): 629-632.
WANG XR, ZHAO Y, LI TS, 2007. Preliminary report on artificial supplementary pollination experiment in clonal seed orchard of Pinus armandii [J]. J SW For Univ, (3): 25-28. [王秀榮, 趙楊, 李桐森, 2007. 華山松無性系種子園輔助授粉試驗初報 [J]. 西南林學院學報, (3): 25-28.]
WHEELER NC, JECH KS, 1992. The use of electrophoretic markers in seed orchard research [J]. New For, 6(1): 311-328.
WIDEN B, SVENSSON L, 1992. Conservation of genetic variation in plants: The importance of population size and gene flow [M]//HANSSON L(ed.). Ecological principles of nature conservation:Application in temperate and boreal environments [M]. New York: Elsevier Science Publishers Ltd: 113-161.
WRIGHT S, 1950. Genetical structure of populations [J]. Nature, 166(4215): 247-249.
XIANG H, YUAN DY, FAN XM, et al., 2016. The analysis of genetic diversity of Castanea henryi (Skan) Rehder & E.H. Wilson by using SRAP [J]. J Plant Gene Resour, 17(6): 1072-1081. [向暉, 袁德義, 范曉明, 等, 2016. 錐栗種質資源遺傳多樣性的SRAP分析 [J]. 植物遺傳資源學報, 17(6): 1072-1081.]
XIE DM, CHEN GM, HE D, et al., 2014. Comparative study on the different organization of genomic DNA extraction in Pinus armandii Franch [J]. J Hubei Inst Natl(Nat Sci Ed), 32(3): 255-261. [謝冬梅, 陳國明, 何德, 等, 2014. 華山松不同部位DNA提取方法比較研究 [J]. 湖北民族學院學報(自然科學版), 32(3): 255-261.]
XIN PY, ZHOU J, DUAN AA, et al., 2010. Research advance of genetic improvement on Pinus armandii in China [J]. N Hortic, (19): 210-214. [辛培堯, 周軍, 段安安, 等, 2010. 我國華山松遺傳改良研究進展 [J]. 北方園藝, (19): 210-214.]
YAN JJ, BAI SH, ZHANG XQ, et al., 2010. Genetic diversity of wild Elymus sibiricus germplasm from the Qinghai-Tibetan Plateau in China detected by SRAP markers [J]. Acta Pratac Sin, 19(1):173-183. [鄢家俊, 白史且, 張新全, 等, 2010. 青藏高原老芒麥種質基于SRAP標記的遺傳多樣性研究 [J]. 草業學報, 19(1):173-183.]
YANG ZQ, FENG YH, WU DS, 2014. Analysis of genetic diversity of Pinus yunnanensis var. tenuifolia nature populations by SSR marker [J]. Guihaia, 34(1):10-14. [楊章旗, 馮源恒, 吳東山, 2014. 細葉云南松天然種源林遺傳多樣性的SSR分析 [J]. 廣西植物, 34(1):10-14.]
YUAN HN, CHEN WH, 2010. Development and utilization status and protection measures of Pinus armandii [J]. For Shaanxi, (4): 39. [袁紅寧, 陳文輝, 2010. 華山松的開發利用現狀及保護措施 [J]. 陜西林業, (4): 39.]
ZHAI SW, CHEN QG, DAI Y, et al., 2007. Heritability analysis and superior family selection for Half-Sib test of Pinus armandii [J]. Seed, 26(12): 5-8. [翟思萬, 陳啟貴, 代毅, 等, 2007. 華山松半同胞子代測定遺傳力分析及優良家系選擇 [J]. 種子, 26(12): 5-8.]
ZHANG AS, SI QL, QI XJ, et al., 2018. Genetic diversity and fingerprints of Actinidia germplasm resource based on SRAP markers [J]. Jiangsu J Agric Sci, 34(1): 138-144. [張安世, 司清亮, 齊秀娟, 等, 2018. 獼猴桃種質資源的SRAP遺傳多樣性分析及指紋圖譜構建 [J]. 江蘇農業學報, 34(1): 138-144.]
ZHANG DD, 2008. Study on genetic structure in different populations of Pinus koraiensis by SRAP [D]. Harbin: Northeast Forestry University. [張冬東, 2008. SRAP對不同地理種源紅松遺傳結構的研究 [D]. 哈爾濱: 東北林業大學.]
ZHAO Y, DAI Y, LI YP, 2012. Genetic diversity for clonal seed orchard of Pinus armandii [J]. J NE For Univ, 40(10): 4-6. [趙楊, 代毅, 李玉璞, 2012. 華山松無性系種子園遺傳多樣性分析 [J]. 東北林業大學學報, 40(10): 4-6.]
ZHAO Y, LI TS, DUAN AA, 2003. A research advance in genetic improvement and development strategy of Pinus armandii in China [J]. J SW For Univ, 23(1): 91-95. [趙楊, 李桐森, 段安安, 2003. 國內華山松遺傳改良的研究進展及發展策略 [J]. 西南林學院學報, 23(1): 91-95.]
ZHAO Y, LI YP, DAI Y, 2012. Optimization of SRAP-PCR system for Pinus armandii [J]. J NW For Univ, 27(5): 87-90. [趙楊, 李玉璞, 代毅, 2012. 華山松SRAP-PCR反應體系的優化 [J]. 西北林學院學報, 27(5):87-90.]
ZHOU YQ, 2008. Biological genetic marking and application [M]. Beijing: Chemical Industry Publishing House: 135-137. [周延清, 2008. 生物遺傳標記與應用 [M]. 北京:化學工業出版社: 135-137.]
ZHU J, 2016. Population genetics and speciation study of Pinus armandii and its three affinities [D]. Xian: Northwestern University. [祝娟, 2016. 華山松及其近緣種的群體遺傳學和物種形成研究 [D]. 西安:西北大學.]
ZHU XD, 2006. Study on the genetic diversity and seed production in clone seed orchard of Pinus armandii Franch. [D]. Kunming: Southwest Forestry College. [朱曉丹, 2006. 華山松無性系種子園遺傳多樣性及結實研究 [D]. 昆明:西南林學院.]
(責任編輯 何永艷)
