趨化素樣因子超家族成員6的研究進展①
2020-06-06 02:45:02蔣敬庭蘇州大學附屬第三醫院腫瘤生物診療中心江蘇省腫瘤免疫治療工程技術研究中心蘇州大學細胞治療研究院常州213003
中國免疫學雜志 2020年9期
韋 俐 蔣敬庭 (蘇州大學附屬第三醫院腫瘤生物診療中心,江蘇省腫瘤免疫治療工程技術研究中心,蘇州大學細胞治療研究院,常州 213003)
含CKLF樣MARVEL跨膜結構域基因(CKLF-like MARVEL transmembrane domain-containing,CMTM)是一個新的蛋白質家族,包括八個CMTMs基因(CMTM1-CMTM8)和一個趨化因子樣因子基因(chemokine-like factor,CKLF)[1]。八個CMTMs基因由于含有CKLF樣的跨膜結構域,所以具有傾向于趨化因子的功能而廣泛參與體內的信號轉導、細胞增殖、細胞遷移和細胞活化等生命過程。尤其是在人源的多種腫瘤中都已發現有CMTMs家族蛋白的參與。2017年發表于Nature上兩篇關于CMTM4蛋白和CMTM6蛋白(主要是CMTM6蛋白)介導程序性死亡受體1配體(PD-L1)獨特保護機制的報道使得CMTM6分子得到廣泛關注[2,3]。盡管有關CMTM6功能和作用的研究還比較少,但是CMTM6在多種腫瘤以及腫瘤微環境中的高表達表明了CMTM6存在作為潛在研究靶點的價值。
1 CMTM6的促癌作用機制
CMTM6基因屬于CMTMs家族,位于3號染色體p22區。CMTM6蛋白由183個氨基酸組成,分子為20.4 kD。與該家族其他成員一樣,CMTM6蛋白是具有MARVEL樣結構域的3型跨膜蛋白,包含至少三個跨膜螺旋結構,主要定位于細胞質膜上,在細胞質內或中間絲上也都有定位[4]。有研究指出,CMTM6的缺失會降低腫瘤特異性T細胞活性并引起腫瘤“免疫逃逸”,從而顯示CMTM6基因可能是腫瘤發生中的抑癌基因[5]。但是,Li等[6]在HeLa宮頸癌細胞中過表達CMTM6蛋白后發現并沒有影響癌細胞的增殖情況,表明CMTM6可能不具有腫瘤抑制功能。并且更多的研究表明,CMTM6 在結腸癌、非小細胞肺癌、膠質母細胞瘤、黑素瘤等多種腫瘤中都有高表達現象[3,6-9],并且與“促癌基因”PD-L1的表達具有一致性。PD-L1的高表達水平可以通過PD-1/PD-L1途徑抑制效應T細胞的活性使腫瘤細胞獲得“免疫逃逸”,而CMTM6的高表達在黑素瘤中正是維持高PD-L1蛋白池含量的關鍵調節分子[3,10,11]。因此,越來越多的數據傾向于認為CMTM6在腫瘤中具有促進腫瘤的作用,當然這種作用可能是直接影響腫瘤細胞的增殖,也有可能是通過調節腫瘤微環境來實現的[7,12,13]。在臨床上高表達的CMTM6也與腫瘤預后不良相關[14]。
盡管在大多數的腫瘤組織或者腫瘤細胞系中CMTM6都是高表達的,但是在某些食管腫瘤中,初步研究卻發現CMTM6的mRNA和蛋白水平都是下調的,這提示CMTM6可能在不同的腫瘤中具有不同表達現象以及不同的調節作用[6]。
2 CMTM6介導的PD-L1穩定性
PD-1/PD-L1作為人體中最重要的免疫檢查點分子之一,靶向PD-1的單抗制劑已經成為治療PD-L1高表達的晚期非小細胞肺癌(NSCLC)患者的一線臨床用藥[11,15]。事實上,PD-L1在多種晚期高危腫瘤都具有高表達現象,同樣在這些腫瘤中也存在CMTM6蛋白的高表達現象。因為MARVEL結構蛋白具有調節跨膜運輸和蛋白分泌的能力,具有MARVEL結構的CMTM6同樣在蛋白質穩定和再循環中起作用[4]。Marian L.Burr等的研究發現,在用35S-半胱氨酸/蛋氨酸進行脈沖標記的CMTM6敲除細胞中,CMTM6基因的缺失會導致PD-L1蛋白的迅速降解并降低其在腫瘤細胞表面的表達,但是沒有損害PD-L1在內質網中的運輸及其在高爾基體中的交換。進一步研究發現,CMTM6與PD-L1的特異性結合可以維持PD-L1/CMTM6蛋白復合體在內含體中的再循環而保護PD-L1蛋白不被溶酶體識別并降解,維持PD-L1蛋白的高表達。更為關鍵的是,這種保護機制不僅對腫瘤細胞中固有的PD-L1存在作用,對干擾素(IFNγ)誘導的PD-L1同樣存在作用。而干擾素誘導PD-L1表達增加是通過激活轉錄因子增加PD-L1 mRNA來促進PD-L1蛋白表達的,CMTM6基因的缺失并沒有影響腫瘤細胞中固有的PD-L1 mRNA水平,也沒有影響IFNγ誘導的PD-L1 mRNA水平[2]。
在另一項研究中,Mezzadra等[3]發現CMTM6可增加PD-L1蛋白池,而不會影響PD-L1基因的轉錄水平。進一步研究表明,在CMTM6缺失的細胞中,PD-L1的泛素化修飾會增加,而被泛素化標記的底物蛋白會被26S蛋白酶體識別而有可能經歷溶酶體降解途徑。CMTM6與E3泛素連接酶STUB1可能存在某種調節機制,在腫瘤細胞中抑制了PD-L1的泛素化修飾,從而阻止了高表達的PD-L1被降解。事實上,除了CMTM6,CMTM家族的另一個成員CMTM4同樣具有調節PD-L1蛋白穩定性的作用。CMTM4與CMTM6具有55%的序列相似性。當CMTM6缺失時CMTM4會被作為“備胎”啟用來維持PD-L1的穩定性,只是CMTM4的調節效率要低于CMTM6[4]。
CMTM6對PD-L1特異性的保護機制可能是眾多腫瘤組織和細胞系中同時存在高表達的重要原因,而PD-L1陽性表達的腫瘤細胞或者抗原呈遞細胞一定程度上意味著腫瘤微環境中T細胞免疫衰竭[16,17]。相關研究也表明,CMTM6的敲除能極大地降低對腫瘤特異性T細胞活性的抑制[5]。總之,CMTM6介導的PD-L1內含體再循環機制有效保護PD-L1避免被泛素化-溶酶體降解途徑降解,從而使腫瘤細胞和部分抗原呈遞細胞表面的PD-L1處于高表達水平,進一步增加腫瘤細胞的獲得性免疫逃逸。因此,CMTM6與眾多腫瘤的發生和發展都存在著關聯。
3 CMTM6/PD-L1在DNA損傷中的作用
細胞對DNA損傷反應同時受到蛋白網絡的調控,這其中轉導蛋白ATM激酶和效應蛋白WEE1核激酶起著重要調控作用[18]。有研究表明,WEE1和ATM都可以促進DNA修復來維持腫瘤進展中的細胞存活[19]。相反,在許多具有DNA損傷反應的腫瘤中,抑制ATM和WEE1可以消除DNA損傷后的修復,起到協同殺傷腫瘤的效果[19-22]。然而,腫瘤中DNA損傷后會上調PD-L1的表達,但是在抑制ATM或WEE1后,CMTM6的表達顯著下降并且PD-L1和CMTM6的相互作用也顯著降低[23,24]。因為CMTM6可以促進PD-L1內含體再循環保護PD-L1不被溶酶體降解,另外有研究表明HIP1R能夠促進遞送PD-L1到溶酶體中[25]。因此,在腫瘤進程中 WEE1或者ATM可能促進了CMTM6翻譯過程表達并同HIP1R競爭與PD-L1的結合,降低PD-L1的溶酶體遞送,但是更有可能是促進了CMTM6基因組的修復,進而維持了腫瘤中CMTM6的高表達[23]。
4 CMTM6/PD-L1在基因組穩定性中的意義
Tu等[26]的研究發現細胞內PD-L1可以作為RNA結合蛋白并調節NBS1,BRCA1和其他DNA損傷相關基因的mRNA的穩定性。PD-L1通過與RNA外泌體競爭,保護目標RNA免受降解,增加細胞對DNA損傷的抵抗力。這說明,PD-L1除了發揮PD-1/PD-L1軸的“免疫剎車”效用,還存在其他重要的調控作用。事實上,PD-L1可以調節整改基因組RNA穩定性,但是在消除CMTM6與PD-L1之間的相互作用時,PD-L1的穩定性被破壞,同時PD-L1與靶基因的RNA的結合也被破壞,導致靶RNA的降解[26]。CMTM6/PD-L1對基因組RNA的穩定性調節也間接證明CMTM6/PD-L1對腫瘤發生及腫瘤細胞本身的生存狀態有著更重要的調節作用。
5 CMTM6與腫瘤的關系
隨著研究的不斷深入,人們發現CMTM6與多種腫瘤的發生發展存在密切的關系,見表1。
5.1CMTM6與NSCLC 靶向PD-1的單抗制劑Pembrolizumab已經成為治療PD-L1高表達的晚期非小細胞肺癌患者的一線臨床用藥[11,15]。而CMTM6介導的PD-L1的保護機制也同樣存在于非小細胞肺癌細胞中[3]。Riman等[13]發現在438個NSCLC臨床患者組織中70%的腫瘤和腫瘤基質中檢測到CMTM6的表達,并且CMTM6的表達與臨床特征或EGFR/KRAS突變狀態無關,但是與T細胞浸潤之間存在相關性。這說明在NSCLC中CMTM6可能與淋巴細胞的免疫活性有關,而與腫瘤基因表達的變化無關,也就是說CMTM6可能依然只是發揮著特異性調節PD-L1穩定性的作用。雖然在肺癌患者中CMTM6與PD-L1存在相關性,但是他們在腫瘤基質中的相關性要高于在腫瘤細胞中,并且CMTM6與PD-L1共表達的患者在接受免疫治療后會獲得更長的總生存期[3,27]。同樣,Yong等[8]在晚期難治性NSCLC患者中也發現CMTM6表達與PD-L1表達呈正相關,并且這種相關性是蛋白水平和基因水平上同時存在的。在接受PD-1抑制劑治療的患者中,對PD-1抑制劑有應答的患者具有較高的CMTM6表達,而PD-L1表達與這種響應性則無關。因為PD-1抑制劑主要是阻斷T細胞表面的PD-1受體,而CMTM6對PD-1并沒有特異性作用[11,28]。這提示,在PD-1抑制劑恢復的腫瘤中T細胞殺傷效應時腫瘤及腫瘤微環境中存在著某種機制去促進CMTM6的表達以恢復PD-L1介導的免疫抑制作用。這可能和PD-L1恢復基因組RNA穩定有關,其中CMTM6 的RNA 本身有可能就是PD-L1的靶點[26]。
5.2CMTM6與肝癌 表觀遺傳基因調控與多種腫瘤的發生有關,DNA甲基化是主要的表觀遺傳修飾之一并在控制基因表達中發揮作用,而啟動子鳥嘌呤殘基CpG島基因座上的異常甲基化常常會導致活躍基因失活或加劇失活基因的沉默[29]。在肝癌以及新生肝癌中CMTM6的DNA甲基化也參與了肝癌的發生和發展過程。Atsunori等[29]研究發現在用丁基哌啶丁酶(PBO)誘導小鼠新生病灶區周圍的非瘤肝細胞中CMTM6的DNA啟動子區域的甲基化水平有輕微增加的趨勢,但是CMTM6的翻譯水平是下降的。相反的是,在PBO誘導的肝細胞癌增生病灶的CMTM6的甲基化并沒有改變。這說明,新生腫瘤病灶區與周圍非癌肝細胞中CMTM6 DNA甲基化的改變可能是誘導正常肝細胞向癌細胞轉變的重要調節過程。
表1 CMTM6在不同腫瘤中的表達意義及作用機制
Tab.1 Expression significance and mechanism of CMTM6 in different tumors
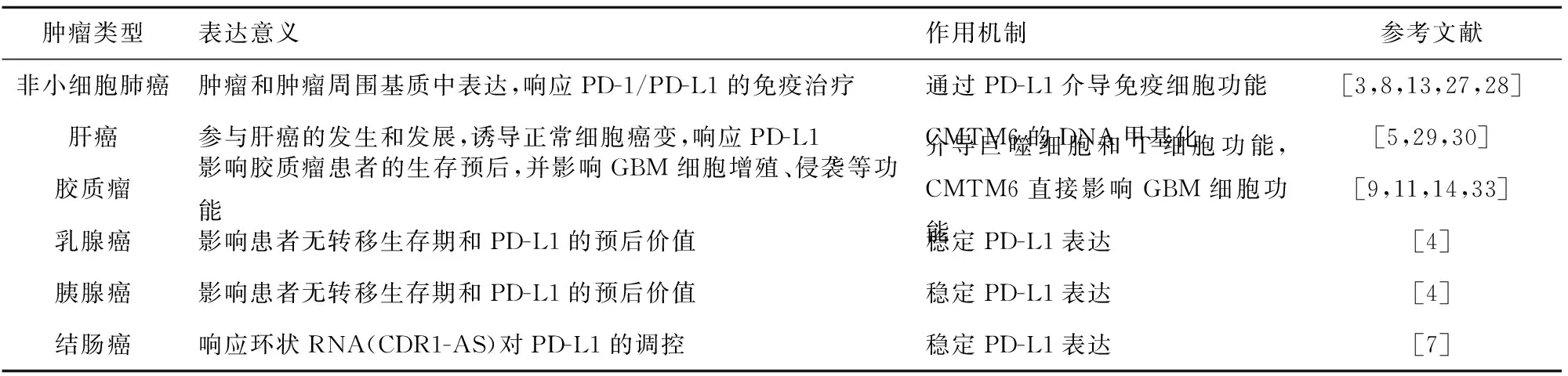
腫瘤類型表達意義作用機制參考文獻非小細胞肺癌腫瘤和腫瘤周圍基質中表達,響應PD-1/PD-L1的免疫治療通過PD-L1介導免疫細胞功能[3,8,13,27,28]肝癌參與肝癌的發生和發展,誘導正常細胞癌變,響應PD-L1 CMTM6的DNA甲基化[5,29,30]膠質瘤影響膠質瘤患者的生存預后,并影響GBM細胞增殖、侵襲等功能介導巨噬細胞和T細胞功能,CMTM6直接影響GBM細胞功能[9,11,14,33]乳腺癌影響患者無轉移生存期和PD-L1的預后價值穩定PD-L1表達[4]胰腺癌影響患者無轉移生存期和PD-L1的預后價值穩定PD-L1表達[4]結腸癌響應環狀RNA(CDR1-AS)對PD-L1的調控穩定PD-L1表達[7]
相反,Zhu等[5]在75對肝細胞癌和鄰近肝腫瘤組織的臨床標本中也發現CMTM6在肝癌組織中的陽性表達水平明顯低于相鄰的非腫瘤組織,并且CMTM6的下調與肝細胞癌的轉移以及與患者的預后有關。這樣的臨床統計結果也與其他腫瘤中CMTM6的高表達現象相反。這提示我們,CMTM6可能在腫瘤發生階段和腫瘤發展階段所發揮的作用不同。研究表明,在肝癌的早期階段腫瘤的抑癌基因甲基化程度是呈現增加趨勢的,而表現出抑癌基因翻譯水平降低,抑癌效果降低[30]。也就是說,CMTM6有可能在正常組織和肝細胞癌變發生階段是表現出抑癌作用的,但是在腫瘤發展階段又發揮了促癌作用。而CMTM6兩種不同情況的研究結果也可能和PD-L1的表達密切相關。正如上文所提的,PD-L1具有調節基因組穩定性的能力,而在惡性腫瘤中PD-L1普遍具有高表達水平。因此,CMTM6在肝癌發生、發展甚至與惡化程度可能都有直接的關聯。
5.3CMTM6 與腦膠質瘤 膠質母細胞瘤(GBM)是常見的惡性腫瘤,具有侵略性的生物行為和對治療的抵抗性[11]。而PD-L1的表達與腫瘤的惡性程度密切相關,所以多項研究也表明PD-L1在GBM細胞中高表達[31,32]。由于PD-L1的穩定性需要CMTM6介導的保護作用,理論上CMTM6在GBM細胞中應處于高表達。事實上,有研究發現CMTM6和膠質瘤的病理級別密切相關,更重要的是,CMTM6高表達與膠質瘤患者的生存預后也存在一定關系[14]。本課題組研究發現,CMTM6高表達患者的五年累積總生存率和無病生存率較低表達患者均明顯下降。該研究還認為,CMTM6與膠質瘤的強相關性除了與CMTM6對PD-1/PD-L1軸的調節有關外,還與CMTM6的激活腫瘤相關巨噬細胞和功能失調的T細胞有關[14]。Gabriele等研究發現CMTM6基因本身就可以調節巨噬細胞,在沉默CMTM6基因后巨噬細胞攝取低密度脂蛋白(LDL)的能力下降[33]。氧化型LDL可以誘導正常巨噬細胞向M2型極化的巨噬細胞轉化,M2極化巨噬細胞正是維持膠質瘤微環境并抑制T細胞活化的重要負性調節細胞[11]。此外,CMTM6對膠質瘤細胞本身也具有直接調控作用。Delic等[9]發現CMTM6 在腦膠質母細胞瘤中被上調,并且在沉默CMTM6基因后膠質瘤細胞U251的細胞增殖顯著受到抑制。本課題組也發現在膠質瘤細胞U87和U251中CMTM6的mRNA水平和蛋白水平都顯著增加,并且在沉默CMTM6基因后不僅顯著抑制細胞的增殖,還顯著降低膠質瘤細胞的遷移和侵襲。所以,在膠質瘤中CMTM6可能還發揮與其趨化因子功能相關的作用,而不僅僅只是保護PD-L1不被泛素-溶酶體途徑降解。
5.4CMTM6與其他腫瘤 在三陰乳腺癌中,CMTM6和PD-L1具有異常表達現象,但是高表達CMTM6比低表達組擁有更長的無轉移生存期[4],并且CMTM6表達能夠提高PD-L1的預后價值,即高表達PD-L1同時高表達CMTM6的患者擁有良好的預后,五年無轉移生存率占到70%。但是高表達PD-L1同時低表達CMTM6的患者五年無轉移生成率占到57%,而二者都低表達的患者五年無轉移生存率只有40%。在胰腺癌中也同樣觀察到CMTM6相似的異常表達和預后價值,但是在胰腺癌中CMTM6高表達與較短的總生存期有關,CMTM6低表達的胰腺癌患者兩年期總生存率占比49%,遠高于高表達的34%[4]。在結腸癌中,環狀RNA(CDR1-AS)表達可以顯著上調CMTM4和CMTM6的表達,因此增加細胞表面PD-L1蛋白的表達[7]。但在某些食管腫瘤中,CMTM6的mRNA和蛋白水平卻被發現是下調的[6]。
6 展望
CMTM6介導的PD-L1的內含體循環保護揭示了PD-L1調節的新機制,并且CMTM6分子也具備影響PD-1/PD-L1“免疫檢查點”的能力,從而表現出調控T細胞活化實現腫瘤免疫治療的潛在價值。盡管在膠質瘤、非小細胞肺癌、黑素瘤等腫瘤細胞中CMTM6表現出高表達現象,但是在肝癌以及部分食管癌中也存在低表達的現象,這種差異表達可能和研究對象中腫瘤的發展及惡性等級相關。雖然至今并沒有研究揭示CMTM6本身表達的受調控機制,但CMTM6在多種腫瘤中表現出與腫瘤惡性程度相關的發現讓我們相信,CMTM6在腫瘤不同階段可能受到了某種適應性調節機制的調節,而這種機制可能和CMTM6/PD-L1的基因組調節作用以及CMTM6基因的表觀遺傳調控有關,并且這兩種調節機制可能不僅僅存在于腫瘤細胞中。因此,深入探索CMTM6在腫瘤發生和發展中介導的受調控機制將會為腫瘤免疫治療提供新策略。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
中成藥(2016年8期)2016-05-17 06:08:14
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年16期)2015-03-01 02:03:11