銀杏-杜仲葉復合發酵工藝優化及發酵物對肉雞腸道功能的影響
2020-06-08 03:33:10曹銀娣張旭暉孫智遠汪貴斌曹福亮
畜牧與獸醫 2020年6期
關鍵詞:黃酮
曹銀娣,張旭暉,孫智遠,汪貴斌,曹福亮*
(1. 南京林業大學林學院南方現代林業協同創新中心,江蘇 南京 210037;2. 青島農業大學科技處,山東 青島 266109;3. 江蘇農業職業技術學院畜牧獸醫學院,江蘇 句容 212400)
近幾年來,大規模的銀杏和杜仲經濟林栽培項目已經啟動,中國的銀杏葉產量大約每年有4萬噸,杜仲葉的產量也在3萬噸以上。如何將這些林產資源有效利用,成為實際生產中需要解決的問題。我國農村農業部公告第194號:自2020年1月1日起,畜禽類飼料全面禁抗,人們將視角轉向可飼用天然產物飼料添加劑的開發。銀杏葉、杜仲葉作為林產資源天然產物,它們除具有一般樹葉的營養外,還富含黃酮類化合物、綠原酸、萜內酯、聚戊烯醇等生物活性物質。微生物在植物資源發酵過程中起到分解轉化作用,在營養價值及生物活性方面,經過發酵的銀杏葉飼料添加劑均有大幅度的提高[1]。幼齡仔雞的免疫系統尚未發育成熟,機能不健全,對環境應激因素易感,益生素正逐漸成為增強禽類生產性能、改善腸道健康和功能的重要途徑[2]。預期通過營養調控措施進行腸道健康的管理越來越受到專家學者的認可。多項研究表明,使用銀杏葉、杜仲葉及其提取物作為禽畜及水產動物的飼料添加劑,能減少預防用藥,增加食用安全性[3]。國內外關于復合發酵銀杏-杜仲葉作為飼料添加劑對肉雞生長發育的研究報道尚比較缺乏[4]。
本研究采用產阮假絲酵母和黑曲霉菌復合發酵按一定比例復配的杜仲葉和銀杏葉,同時提升總黃酮和綠原酸等活性成分的含量,制備出一種生物飼料添加劑,選用AA肉雞作為研究對象,探討外源添加銀杏-杜仲葉混合發酵物(簡稱“FGE”)對肉雞的生長發育、腸道結構與消化功能的作用,以期為銀杏葉和杜仲葉作為飼料添加劑在家禽飼料中的使用提供理論依據。
1 材料與方法
1.1 試驗材料
1.1.1 原料
銀杏葉:南京林業大學銀杏園,10月樹上采摘,自然晾曬葉;杜仲葉:河南理工大學杜仲園,12月落葉收集,自然晾曬葉。以上材料均60 ℃烘干后、粉碎機磨成粉,40目過篩,裝入自封袋密封保存。
菌種:產朊假絲酵母(Candidautilis)和黑曲霉(Aspergikrsniger)均來自南京林業大學化學工程學院微生物菌種保藏室。于土豆葡萄糖斜面培養基 (PDA)上保存。
固體培養基:以銀杏-杜仲葉為主要發酵原料80%;適量添加麩皮,添加量為20%,含2.0%葡萄糖、3.0% (NH4)2SO4、2.0% KH2PO4、0.5% MgSO4·7H2O。水分含量60%。
所有培養基都需在121 ℃條件下滅菌30 min。在優化試驗過程中,培養基的組成會有所變化。
1.1.2 試驗動物
360只1日齡健康AA肉仔雞(購自山東煙臺蘇佳麗禽業有限公司),體重平均為(49.24±3.89)g,差異不顯著(P>0.05)。
1.2 試驗設計
1.2.1 發酵試驗設計
單因素試驗:采用250 mL錐形瓶作發酵容器,固體重量50 g,厚度1~2 cm,加60%的水,滅菌后接種產朊假絲酵母量1.0%(按培養基的干重比例配)。
銀杏-杜仲葉發酵選擇常規條件:銀杏葉與杜仲葉配比為2∶1,黑曲霉菌為2.0%(按培養基的干重比例配),發酵溫度為28 ℃,發酵時間為96 h。選取銀杏葉與杜仲葉配比1∶2、1∶1、2∶1、3∶1、4∶1,黑曲霉菌0.5%、1.0%、1.5%、2.0%、2.5%,發酵溫度為23、25、28、30、32 ℃,發酵時間為48、72、84、96、120 h作為工藝參數探討值,進行單因素試驗。在單因素試驗基礎上進行四因素三水平L9(34)正交試驗。每個處理做3個重復,每個重復做3次平行試驗,取平均值。
1.2.2 動物試驗設計
在確定最佳的銀杏葉與杜仲葉配比和最佳發酵工藝條件后,選用1日齡(1 d)健康、體重相近的AA肉仔雞360只(公雞苗,P>0.05),完全隨機設計分為6個處理組,每個處理組含6個重復(每個重復10只雞)。分為:基礎日糧對照組;未發酵銀杏-杜仲葉(NFGE組),分別在基礎日糧中添加NFGE前期0.3%,后期0.6%;4個FGE組(FGE1、FGE2、FGE3、FGE4),即在基礎日糧中添加FGE前期分別為0.2%、0.3%、0.4%和0.5%,后期分別為0.4%、0.6%、0.8%和1.0%。
試驗期42 d,分別在試驗的第1、21、42天的早晨07:00空腹稱重并結料,逐只將試雞進行稱重。稱重前1 d的晚上7:00斷料,但持續喂水,計算以重復為單位進行,記錄不同生長階段雞的采食量、體增重及料重比(F/G)。試驗基礎日糧采用玉米-豆粕型,其組成及營養水平見表1。日糧分前期(0~21 d)和后期(21~42 d)2個階段,在不同的日糧處理組中,采用重量替代法配制,使用麩皮來補齊試驗添加物的量。
表1 基礎日糧組成及營養水平
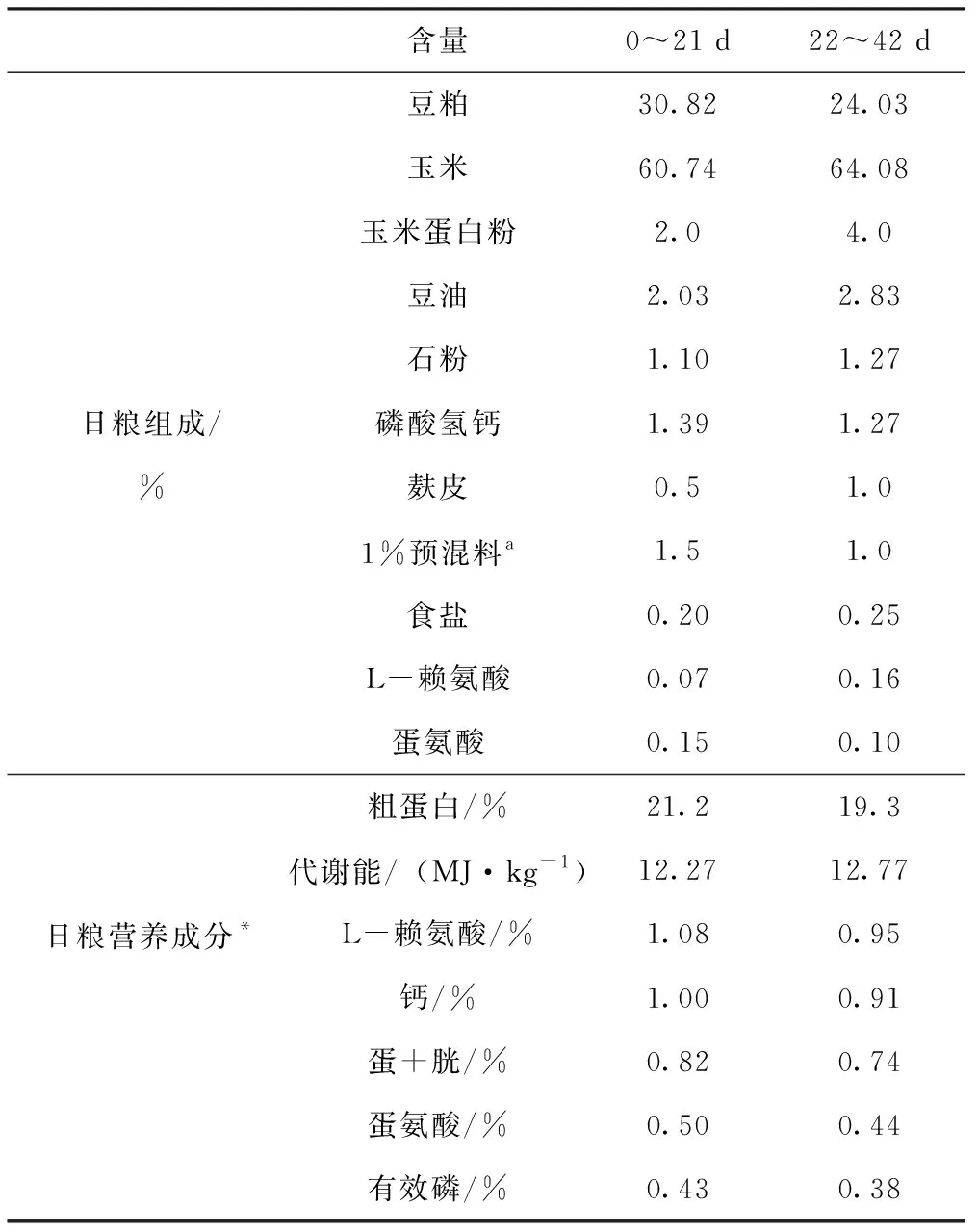
含量0~21 d22~42 d豆粕 30.8224.03玉米 60.7464.08玉米蛋白粉 2.04.0豆油 2.032.83石粉 1.101.27日糧組成/磷酸氫鈣1.391.27%麩皮 0.51.01%預混料a 1.51.0食鹽 0.200.25L-賴氨酸0.070.16蛋氨酸 0.150.10粗蛋白/%21.219.3代謝能/(MJ·kg-1)12.2712.77日糧營養成分*L-賴氨酸/%1.080.95鈣/%1.000.91蛋+胱/%0.820.74蛋氨酸/%0.500.44有效磷/%0.430.38
注:*為計算值。a:每kg日糧提供:鐵,60 mg;銅,7.5 mg;鋅,65 mg;錳,110 mg;碘,1.1 mg;硒,0.4 mg;維生素A,90 mg;維生素B1,2.2 mg;維生素B2,10 mg;維生素B3,50 mg;維生素B5,4 mg;維生素B6,10 mg;維生素B11,1 mg;維生素B12,1.013 mg;維生素D3,20 mg;維生素K,1.3 mg;膽堿,400 mg;維生素H,0.04 mg。
1.3 采樣與測定方法
1.3.1 提取方法
稱取干燥發酵產物5.0 g,在超聲波功率500 W條件下,固液比1∶15、乙醇濃度70%、提取溫度為50 ℃條件下,提取20 min,累計提取2次,合并提取液。
1.3.2 樣品的采集與制備
每個處理于每個重復中隨機抓取1只雞,頸靜脈采血制備血清,置-20 ℃冰箱待測,頸部放血致死。取新鮮的十二指腸、空腸、回腸組織,于每段組織截取1 cm左右的腸段,輕輕用生理鹽水沖洗掉腸道食糜,放入4%甲醛溶液中固定,備用于形態檢測分析分離胰腺、十二指腸、空腸和回腸,均要輕輕擠出腸道各段的食糜,再分別將各段腸管剪開,先輕輕用載玻片刮去各腸段表層的殘留食糜,再用適度刮取黏膜,將黏膜分別于-70 ℃的冰箱保存,備用。
1.3.3 活性成分測定
總黃酮濃度測定:以蘆丁為對照品制備溶液,505 nm波長制定標準曲線,獲得回歸方程:A=11.155 6C+0.006 1,R2=0.999 7。適用濃度0~48.0 mg/L。
綠原酸濃度的測定:以綠原酸為對照品制備溶液,328 nm波長制定標準曲線,獲得回歸方程:C=19.049 0A-0.074 4,R2=0.998 4。適用濃度0~38.4 mg/L。
總黃酮、綠原酸含量計算方法:分別在505 nm、328 nm下測定其吸光值,代入相應回歸方程,計算總黃酮、綠原酸濃度C1、C2,計算提取率Y1、Y2。
計算公式為:Y=100%C×N×V/m。式中:Y:發酵物總黃酮或綠原酸提取率;C:樣液中總黃酮或綠原酸濃度,mg/L;N:稀釋倍數;V:提取液體積,L;m:原料質量,mg。
1.3.4 蛋白質和氨基酸含量的測定
采用凱氏定氮法測定復合發酵物中粗蛋白含量(GB/T 14771-93)。發酵物樣品用6 mol/L的HCl在110 ℃水解22 h后測總氨基酸含量,用HP-1050型液相色譜儀進行測定[5]。
1.3.5 腸道形態結構與酶活性的測定
腸道形態采用對腸道切片進行HE染色,采集圖像,分析獲得。測定酶活性試劑盒購自南京建成生物工程研究所,按說明操作。
1個淀粉酶活力單位為:37 ℃條件下,每克組織或食糜或每毫克蛋白與底物作用,30 min,每水解10 mg淀粉,定義為1個淀粉酶活力單位。
1個脂肪酶活力單位為:每克組織及食糜(或每毫克蛋白)37 ℃下,于反應體系中和底物反應1 min,每消耗l μmol底物,定義為1個脂肪酶活力單位。
1個總蛋白酶單位為:在40 ℃和pH=6.0條件下,每克組織及食糜或每毫克蛋白每分鐘水解酪素產生1 μL酪氨酸,定義為1個總蛋白酶活力單位。
1.3.6 小腸黏膜堿性磷酸酶(AKP)活性的測定
采用南京建成生物工程研究所的試劑盒測定十二指腸、空腸和回腸黏膜中AKP活性。組織勻漿上清液中蛋白質含量采用文獻[6]方法進行測定。
1.3.7 血清D-木糖濃度和血清尿素氮的測定
均采用南京建成生物工程研究所試劑盒測定血清D-木糖濃度和血清尿素氮,按說明操作。
1.3.8 統計分析
采用SPSS 19.0軟件進行單因素方差分析 (One-way ANOVA);以IBM SPSS Statistics 19.0軟件采用獨立樣本t檢驗進行統計分析,差異顯著性水平設為P<0.05;采用Duncan氏多重比較法進行顯著性檢驗,差異顯著性水平設為P<0.05(或P<0.01)。數據采用“平均值±標準差”表示。
2 結果與分析
2.1 銀杏-杜仲葉發酵工藝參數研究
2.1.1 銀杏與杜仲葉配比對總黃酮和綠原酸含量的影響
如圖1,隨著銀杏葉質量比重的增加,總黃酮含量有明顯增加(P<0.05),綠原酸含量有所降低,在比例2∶1開始,綠原酸含量降低明顯。隨著杜仲葉比例的增加,綠原酸含量增加明顯(P<0.05),總黃酮含量呈降低趨勢,總黃酮和綠原酸總含量出現先增加后降低的情況。綜合考慮,正交試驗銀杏葉與杜仲葉配比為1∶1、2∶1、3∶1。
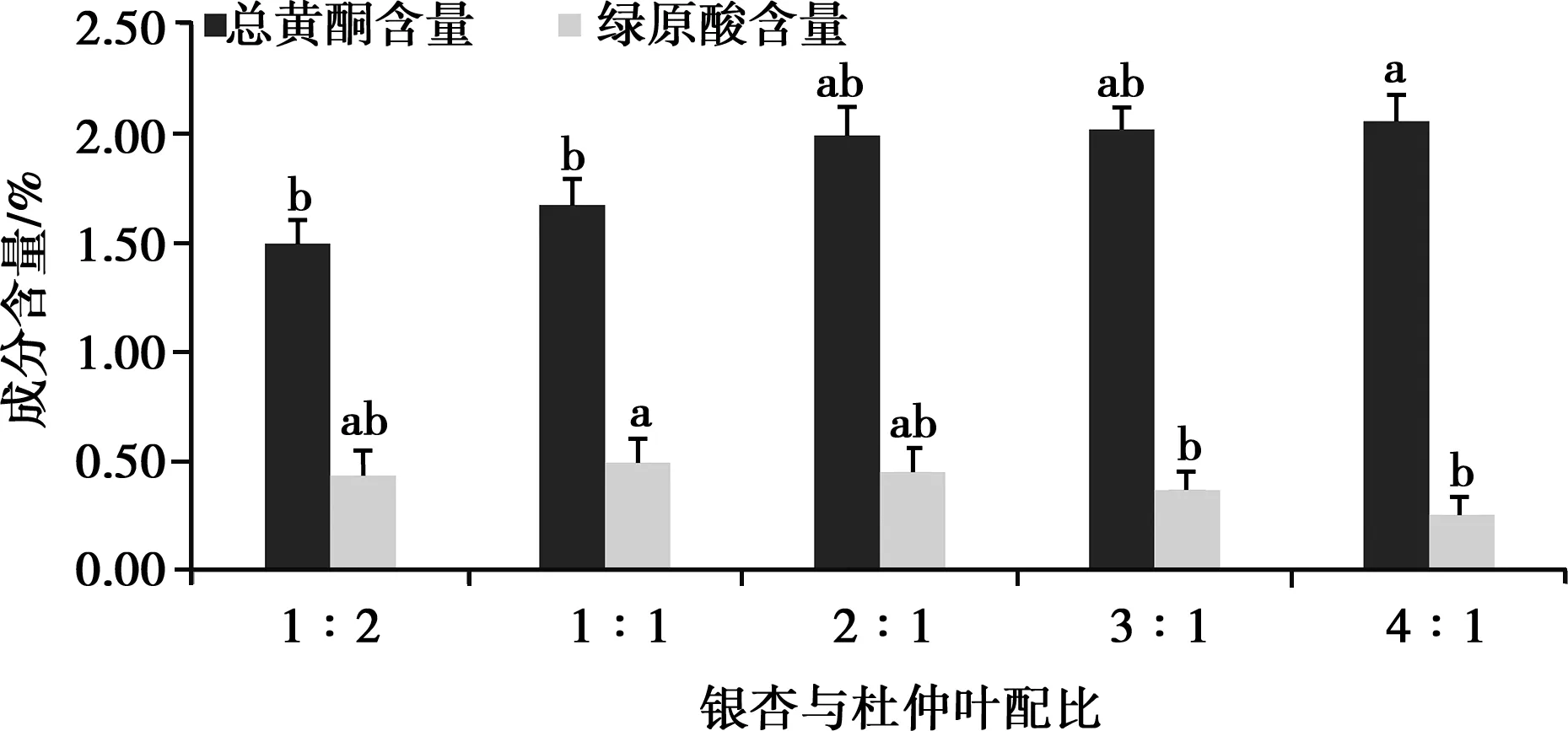
同指標相比,不同字母表示差異顯著(P<0.05),相同字母或無字母表示差異不顯著(P>0.05)。下同
圖1 銀杏葉與杜仲葉配比對總黃酮和綠原酸含量的影響
2.1.2 黑曲霉菌接種量對總黃酮和綠原酸含量的影響
如圖2,隨著黑曲霉菌接種量的增加,總黃酮和綠原酸的含量均出現明顯提升趨勢(P<0.05),當接種量達到2.0%時,總黃酮量增勢減緩。綜合考慮,正交試驗黑曲霉菌接種量設計為1.5%、2.0%、2.5%。
2.1.3 發酵溫度對總黃酮和綠原酸含量的影響
如圖3,隨著發酵溫度的升高,總黃酮和綠原酸的含量呈先升后降,根據變化趨勢,正交試驗發酵溫度設計為25、28、30 ℃。
2.1.4 發酵時間對總黃酮和綠原酸含量的影響
如圖4,隨著發酵時間的延長,總黃酮和綠原酸的含量均有明顯提升,總黃酮含量出現明顯下降,綜合考慮,正交試驗發酵時間設計為72、84、96 h。
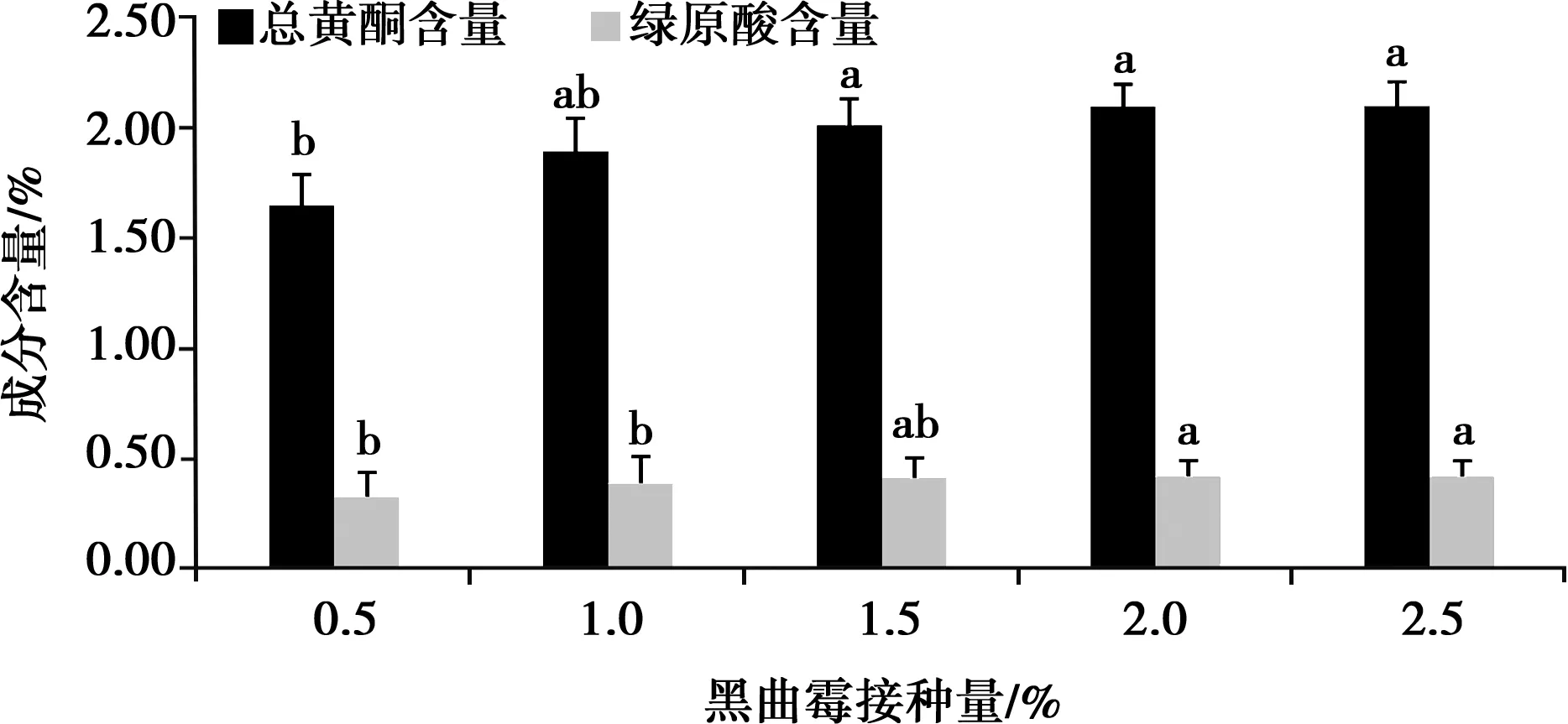
圖2 黑曲霉菌接種量對總黃酮和綠原酸含量的影響
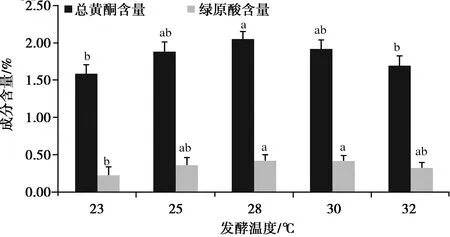
圖3 發酵溫度對總黃酮和綠原酸含量的影響
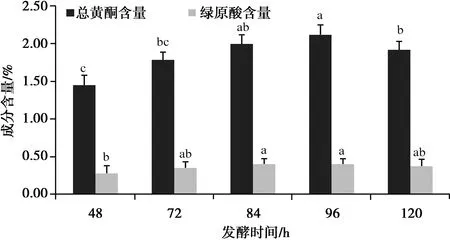
圖4 發酵時間對總黃酮和綠原酸含量的影響
2.1.5 銀杏-杜仲葉復合發酵工藝條件優化
根據單因素試驗的結果,確定銀杏葉與杜仲葉的質量比例(A)、黑曲霉菌接種量(B)、發酵溫度(C)、發酵時間(D)為試驗因素,如表2來設計正交試驗因素水平,優化銀杏-杜仲葉復合植物資源發酵工藝結果如表3。
表2 銀杏-杜仲葉復合發酵工藝正交試驗因素水平
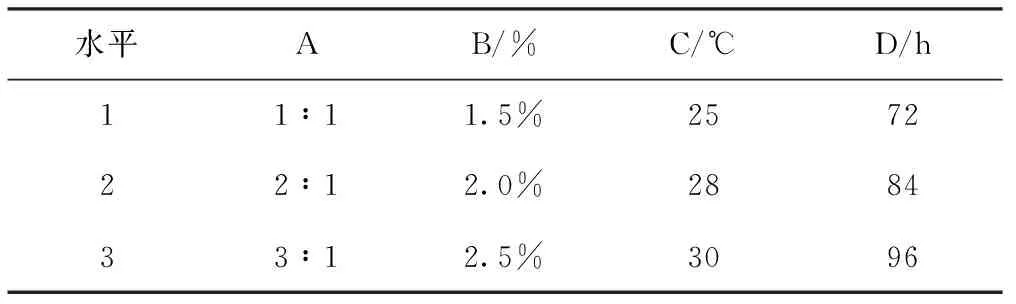
水平AB/%C/℃D/h11∶11.5%257222∶12.0%288433∶12.5%3096
表3 銀杏-杜仲葉復合發酵工藝正交試驗結果
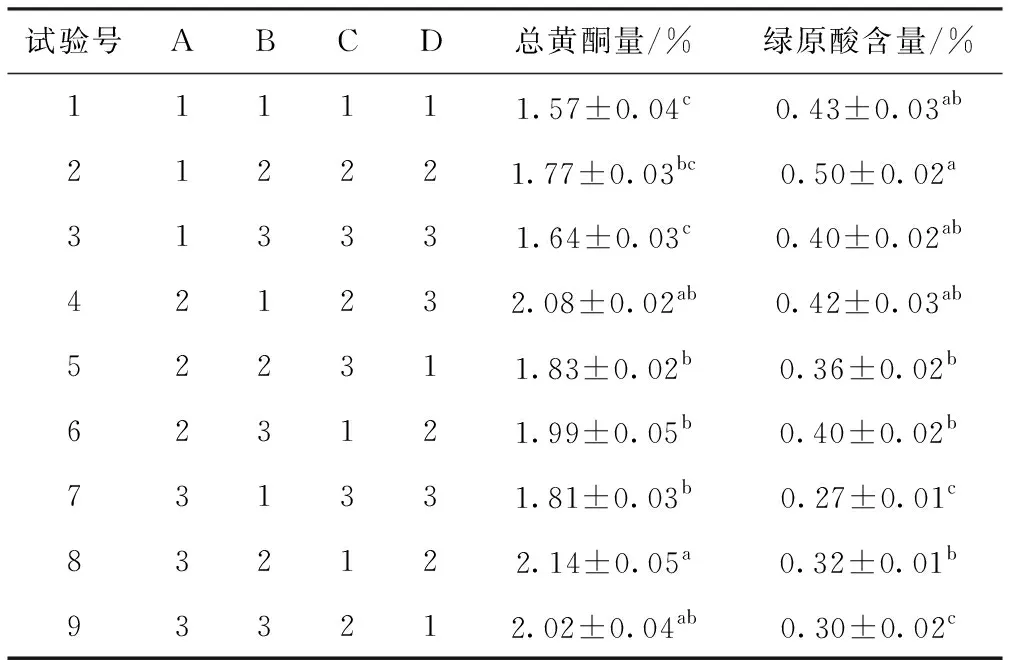
試驗號ABCD總黃酮量/%綠原酸含量/%111111.57±0.04c0.43±0.03ab212221.77±0.03bc0.50±0.02a313331.64±0.03c0.40±0.02ab421232.08±0.02ab0.42±0.03ab522311.83±0.02b0.36±0.02b623121.99±0.05b0.40±0.02b731331.81±0.03b0.27±0.01c832122.14±0.05a0.32±0.01b933212.02±0.04ab0.30±0.02c
注:同列數據肩標不同字母表示差異顯著(P<0.05),相同字母或無字母表示差異不顯著(P>0.05)。
表3數據所示,4個因素對總黃酮提取率和綠原酸含量影響均有顯著性差異,對總黃酮含量影響力依次為A>C>D>B,總黃酮含量最高的工藝組合為A3B2C1D2;對綠原酸提取率影響力依次為A>C>D>B,綠原酸含量最高的工藝組合為A1B2C2D2。分析單因素試驗趨勢和正交試驗結果,取A2B2C2D3和A2B2C3D3進行最優化工藝參數比較確定試驗,結果顯示總黃酮和綠原酸的含量分別為2.13%、0.50%和2.12%、0.43%。最終選定A2B2C2D3為最佳工藝條件,此條件下固態發酵產品總黃酮含量為2.13%,綠原酸的含量為0.50%。
2.1.6 銀杏-杜仲葉發酵產物的主要營養成分及活性成分分析
對選定條件下銀杏-杜仲葉發酵產品中主要營養成分和生物活性成分進行分析,經t檢驗分析如表4,發酵后相對應發酵前各項主要營養成分和活性成分均有顯著提高(P<0.05)。粗蛋白的含量從發酵前的12.41%增加到25.23%,提高了103.30%;總氨基酸、必需氨基酸含量分別從發酵前的9.76%和3.74%增加到17.49%和6.34%,分別提高了79.20%和69.52%;總黃酮含量發酵前的1.67%增加到2.13%,提高了27.55%;綠原酸含量由發酵前的0.42%增加到0.50%,提高了19.48%。

表4 銀杏-杜仲葉發酵產物主要營養成分及活性成分分析 %
注:肩標*表示與發酵前對應值差異顯著(P<0.05)。
2.2 動物試驗
2.2.1 FGE對肉雞生產性能的影響
由表5可知,在1~21 d的生長發育階段中,各組間的日增重和日采食量均無顯著差異(P>0.05),22~42 d和1~42 d的日增重無顯著差異(P>0.05);FGE2和FGE3組22~42 d的日采食量與對照組相比顯著降低(P<0.05);F/G在1~21 d各FGE組間無顯著差異(P>0.05),但與對照組和NFGE組相比,FGE2和FGE3組的F/G在22~42 d和1~42 d間有顯著降低(P<0.05)。
表5 日糧添加FGE對肉雞生長性能的影響
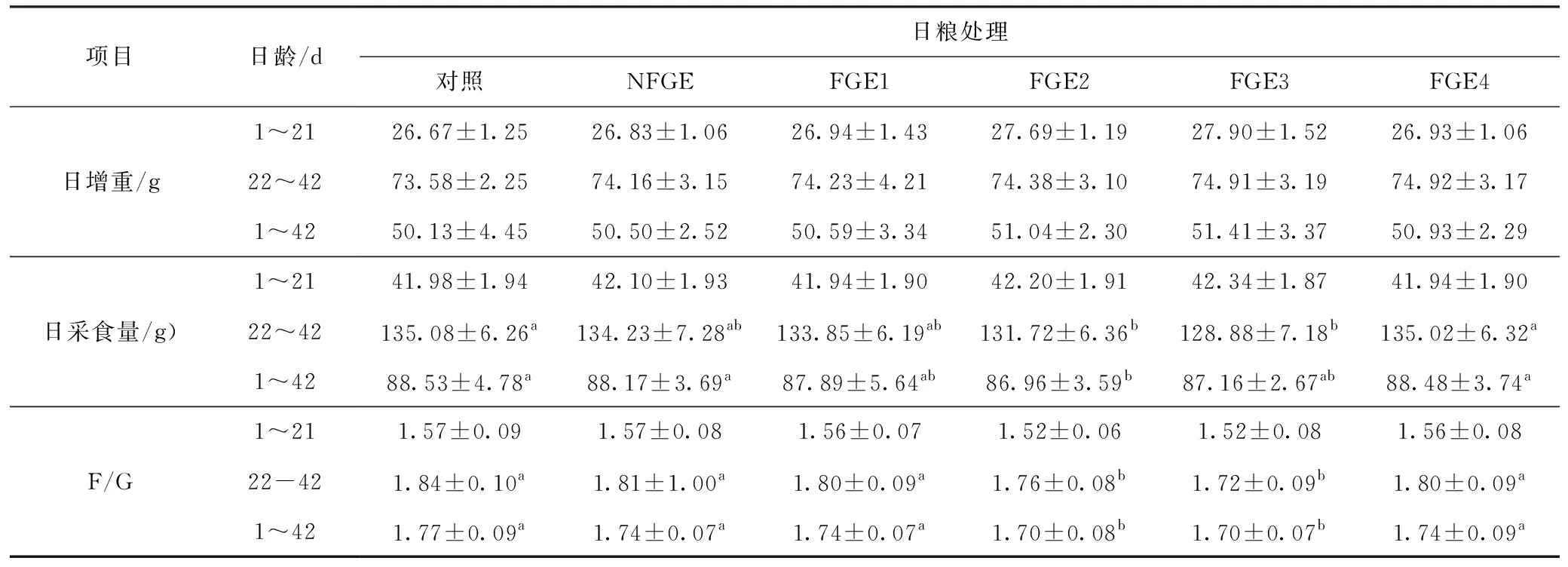
項目日齡/d日糧處理 對照NFGEFGE1FGE2FGE3FGE4日增重/g1~2126.67±1.2526.83±1.0626.94±1.4327.69±1.1927.90±1.5226.93±1.0622~4273.58±2.2574.16±3.1574.23±4.2174.38±3.1074.91±3.1974.92±3.171~4250.13±4.4550.50±2.5250.59±3.3451.04±2.3051.41±3.3750.93±2.29日采食量/g)1~2141.98±1.9442.10±1.9341.94±1.9042.20±1.9142.34±1.8741.94±1.9022~42135.08±6.26a134.23±7.28ab133.85±6.19ab131.72±6.36b128.88±7.18b135.02±6.32a1~4288.53±4.78a88.17±3.69a87.89±5.64ab86.96±3.59b87.16±2.67ab88.48±3.74aF/G1~211.57±0.091.57±0.081.56±0.071.52±0.061.52±0.081.56±0.0822-421.84±0.10a1.81±1.00a1.80±0.09a1.76±0.08b1.72±0.09b1.80±0.09a1~421.77±0.09a1.74±0.07a1.74±0.07a1.70±0.08b1.70±0.07b1.74±0.09a
注:同行數據肩標不同字母表示差異顯著(P<0.05),相同字母或無字母表示差異不顯著(P>0.05)。下同
2.2.2 FGE對肉雞小腸絨毛形態的影響
由表6可見,與對照組相比,日糧中添加FGE顯著增加了肉雞十二指腸絨毛高度(P<0.05)。與對照組和NFGE組相比,FGE3和FGE4組的空腸絨毛高度顯著增加(P<0.05),FGE2、FGE3、FGE4組空腸的隱窩深度顯著降低(P<0.05)。就回腸而言,與對照組和NFGE組相比,日糧中添加FGE顯著降低了肉雞回腸隱窩深度(P<0.05),顯著提高了絨毛高度與隱窩深度之比(P<0.05)。此外,總體而言,日糧中添加FGE對十二指腸絨毛高度,空腸絨毛高度及隱窩深度產生了線性和二次線性影響。
表6 FGE對肉雞小腸形態及發育的影響

部位指標日糧處理對照NFGEFGE1FGE2FGE3FGE4絨毛高度/μm1 558.00±145.61b1 633.00±156.35ab1 665.00±170.53a1 677.00±165.37a1 684.00±176.42a1 689.00±175.30a十二指腸隱窩深度/μm215.90±15.41214.87±13.46214.01±17.40213.09±19.74211.60±16.44209.19±18.59絨毛高度/隱窩深度7.25±0.357.60±0.397.78±0.427.87±0.478.05±0.488.10±0.53絨毛高度/μm1 141.00±46.78b1 162.00±48.94b1 220.00±57.94ab1 260.00±60.36ab1 287.00±40.59a1 295.00±53.73a空腸隱窩深度/μm192.74±8.54a189.56±9.47a178.62±7.39a172.13±10.48b175.52±8.97b172.67±9.20b絨毛高度/隱窩深度5.92±0.38b6.13±0.42ab6.83±0.54ab7.32±0.47a7.46±0.50a7.50±0.47a絨毛高度/μm819.00±28.43823.00±49.05839.00±50.42851.00±53.85860.00±40.47862.00±52.33回腸隱窩深度/μm109.48±5.67a106.26±6.63a103.53±7.32b101.75±6.94b100.49±7.06b99.61±6.44b絨毛高度/隱窩深度7.48±0.26b7.75±0.20b8.10±0.29a8.36±0.38a8.56±0.37a8.65±0.45a
2.2.3 FGE對肉雞消化酶活性的影響
與對照組相比,FGE2、FGE3和FGE4組的肉雞胰腺蛋白酶活性有顯著升高(P<0.05)(表7)。與對照組相比,FGE3和FGE4組十二指腸和空腸蛋白酶活性、十二指腸淀粉酶活性有顯著提高(P<0.05);日糧中添加FGE顯著增加了空腸淀粉酶的活性(P<0.05)。分析結果表明,隨著日糧中FGE添加水平的增加,對胰腺和十二指腸的蛋白酶活性有線性(P=0.036,P=0.040)影響。
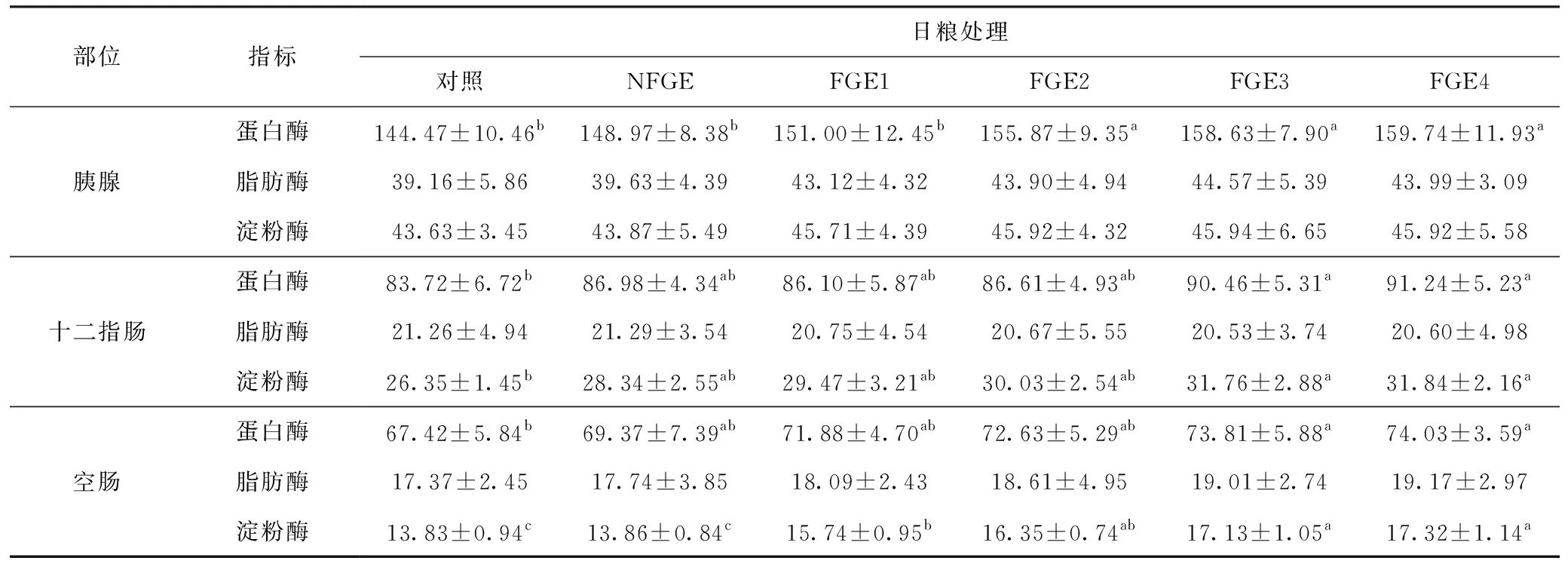
表7 FGE對肉雞胰腺和小腸內容物消化酶活性的影響 U·g-1
2.2.4 FGE對肉雞AKP酶活性的影響
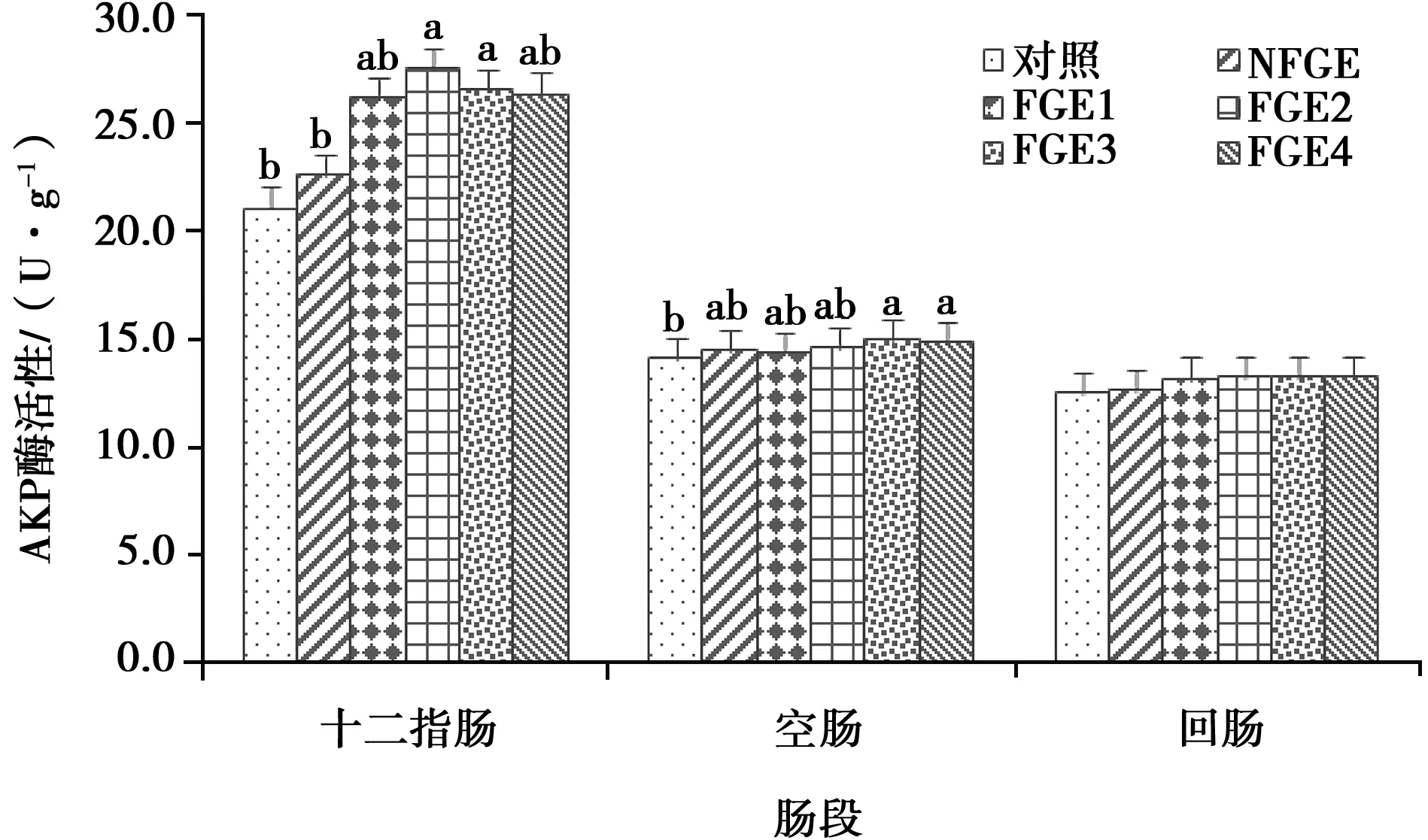
由圖5可見,與對照組和NFGE組相比,FGE2、FGE3組肉雞十二指腸AKP的活性顯著提高(P<0.05);此外,與對照組相比,FGE3和FGE4組的空腸AKP活性也有顯著提高(P<0.05)。
2.2.5 FGE對肉雞血清D-木糖和尿素氮水平的影響
如圖6所示,與對照組和NFGE組相比,日糧中添加銀杏葉發酵物對血清D-木糖水平均有提高,其中FGE2、FGE3組差異顯著(P<0.05)。添加FGE顯著降低血清尿素氮水平(P<0.05)。分析結果表明,隨著日糧中FGE添加水平的增加對血清D-木糖水平有線性(P=0.013)影響。
相同指標,不同字母表示差異顯著(P<0.05),相同字母或無字母表示差異不顯著(P>0.05)。下同
圖5 日糧添加FGE對肉雞小腸黏膜AKP活性的影響

圖6 FGE對肉雞血清D-木糖和尿素氮水平的影響
3 討論
3.1 雙菌發酵對銀杏葉-杜仲葉固態發酵的影響
本試驗采用了黑曲霉菌和產朊假絲酵母菌,混合發酵銀杏葉、杜仲葉原料,使其粗蛋白、氨基酸、總黃酮類和綠原酸含量皆得到提高,這和湯小朋[7]在研究單菌及混菌固態發酵木薯渣品質中認為混菌發酵能夠較好提高產品蛋白質含量的結論是一致的。雙菌發酵可以解除單菌發酵時微生物產生代謝產物累積導致的反饋遏制,如本研究中黑曲霉會產生大量的纖維素酶,分解纖維素產生大量還原糖,這些還原糖除了供應黑曲霉自身利用外,大部分將積累在培養基,從而阻遏黑曲霉繼續分泌纖維素酶[8];而產朊假絲酵母菌恰恰可以利用這些還原糖,從而使發酵持續進行,直至發酵更為徹底,而且還產生了大量的菌體蛋白[9]。這也可以解釋本研究中無論是單葉發酵還是復合葉發酵,最終產品的蛋白含量都有大幅提高。
3.2 發酵溫度和時間對銀杏葉-杜仲葉固態發酵的影響
固態發酵過程中,微生物生長會有大量的熱產生,引起物料溫度變化,需要有效控制發酵溫度。發酵時間也是影響發酵品質的重要因素,發酵過程中,產物濃度隨著發酵時間而變化。本試驗中銀杏葉的發酵溫度為28 ℃、發酵時間為72 h,杜仲葉的發酵溫度為30 ℃、發酵時間為96 h,銀杏-杜仲葉混合發酵的溫度為28 ℃、時間為96 h。和趙林果等[1]以銀杏葉為原料,采用黑曲霉為發酵菌種,生產飼料復合酶的研究:發酵溫度是28-30 ℃,發酵時間72 h的條件相近,低于陳旸等[10]采用植物乳桿菌發酵轉化人參皂苷發酵溫度35℃發酵溫度。低于侯衍英等[11]以紅曲霉雙向固態發酵丹參20 d的發酵時間。這和微生物的種類和具體發酵基質相關。
3.3 FGE對肉雞生產性能的影響
植物資源經過微生物發酵處理,發酵產物中富含各種酶、維生素和生長因子[12],發酵后銀杏黃酮甙元能夠更容易、更迅速地被動物的腸道消化吸收[13]。本研究中,AA肉雞日糧中添加發酵產物FGE顯著改善了F/G,原因可能是日糧中添加銀杏-杜仲葉發酵物改善了肉雞腸道的健康狀態,進而降低了肉雞的脂肪沉積。腹部沉積的脂肪作為家禽產業和家禽市場的一種浪費,被認為是一種額外的損失,本試驗揭示了日糧中添加FGE能夠降低腹部脂肪的沉積,進而改善了動物的飼料效率。
3.4 FGE對肉雞消化機能的影響
腸道是最重要的營養吸收位點,腸道黏膜的結構能夠在一定程度上反應腸道健康與否。食糜中存在的應激因子與黏膜表面近距離接觸,能夠快速改變腸道黏膜的結構。本試驗中,與對照組相比,飼喂FGE組肉雞的十二指腸和空腸的絨毛高度,空腸的絨毛高度與隱窩深度之比都有顯著增加。本試驗中所觀察到的生產性能的改善與腸道結構的改善是相吻合的。
在腸道中,隨著腸細胞的成熟并上移至絨毛頂端,AKP酶活性的表達增加。因此,AKP活性的增加表明腸道中較多數量的官能的和成熟的腸上皮細胞[14]。本試驗中,發酵物處理肉雞的AKP活性比對照組顯著提高,這可能暗示了銀杏-杜仲葉發酵物具有通過刺激AKP的活性而改善機體消化和吸收功能的能力。日糧中添加FGE對AKP活性的刺激作用也支持本試驗中生產性能的結果。
胰腺及腸道消化酶在腸道大分子的消化與吸收中起著關鍵的作用。本試驗中腸道蛋白酶和淀粉酶的活性增加與尿素氮分泌降低的結果相一致。Hong等[15]報道大分子的肽,如抗原蛋白,可能會被降解為小分子的肽。同時,抗營養因子-酶復合物的減少可能也會為營養物質的消化讓出更多的消化酶[16],最終導致對飼料更高效率的利用并改善肉雞的F/G。因此,肉雞日糧中添加FGE對腸道消化酶活性的改善作用可能與FGE中的蛋白質對酶的降解作用有關。
4 結論
銀杏-杜仲葉復合發酵的最佳工藝參數:初始含水量60%,培養基初始pH為5.0,接種產朊假絲酵母菌量為1.00%,銀杏葉與杜仲葉配比為2∶1,黑曲霉菌接種量為2.00%,發酵溫度為28 ℃,發酵時間為96 h,培養基通過生物轉化后營養豐富,總黃酮和綠原酸含量達到最優化,分別為2.13%和0.50%,比發酵前分別提高了27.55%和19.48%。發酵后蛋白含量提高103.30%,必需氨基酸含量提高69.52%。日糧中添加0.2%~0.5%(后期0.4%~1.0%)的FGE,能夠改善腸道結構,增強腸道消化吸收功能,明顯降低料重比,從而提升肉雞的生長性能。
猜你喜歡
四川蠶業(2021年2期)2021-03-09 03:15:32
四川蠶業(2021年3期)2021-02-12 02:38:46
中成藥(2018年11期)2018-11-24 02:57:00
中成藥(2017年8期)2017-11-22 03:19:40
中成藥(2017年10期)2017-11-16 00:50:13
中成藥(2017年4期)2017-05-17 06:09:50
哈爾濱醫藥(2016年1期)2017-01-15 13:43:16
天然產物研究與開發(2016年11期)2016-06-15 20:29:17
湖南師范大學自然科學學報(2015年1期)2015-02-27 14:50:04
安徽醫藥(2014年12期)2014-03-20 13:15:15
