一株產(chǎn)纖維素酶細(xì)菌的發(fā)酵產(chǎn)酶條件優(yōu)化
2020-06-21 15:35:06郭建華張春強(qiáng)郭宏文
江蘇農(nóng)業(yè)科學(xué) 2020年9期
郭建華 張春強(qiáng) 郭宏文
關(guān)鍵詞:產(chǎn)纖維素酶細(xì)菌;FPA酶;CMC酶;發(fā)酵條件優(yōu)化
能源、環(huán)境及工農(nóng)業(yè)原料生產(chǎn)等問題越來越影響著人們的日常生活,高產(chǎn)纖維素酶微生物的篩選對解決這些問題都十分重要[1-3]。大多數(shù)細(xì)菌所產(chǎn)纖維素酶與真菌來源的酶性質(zhì)不同,某些方面具有真菌酶不可替代的作用[4]。細(xì)菌纖維素酶在飼料工業(yè)、洗滌劑工業(yè)及紡織工業(yè)中具有廣泛的利用前景,因此在工業(yè)生產(chǎn)中的地位逐漸提高[5],其中產(chǎn)纖維素酶的芽孢桿菌成為近年來的研究熱點(diǎn)[6]。本試驗以從白酒酒醅中選育得到的產(chǎn)纖維素酶細(xì)菌菌株DM-4為試驗菌株,利用單因素試驗和響應(yīng)面試驗設(shè)計,優(yōu)化其發(fā)酵產(chǎn)酶條件,為進(jìn)一步的試驗研究準(zhǔn)備條件。
1 材料與方法
1.1 試驗菌株
菌株DM-4,由筆者所在實驗室從白酒酒醅中選育得到,經(jīng)鑒定該菌株為枯草芽孢桿菌(Bacillus subtilis)。
1.2 培養(yǎng)基
種子培養(yǎng)基:牛肉膏0.5%,蛋白胨1%,NaCl 0.5%,pH值7.5~7.6,121 ℃下滅菌30 min。發(fā)酵培養(yǎng)基:CMC-Na 10 g,蛋白胨2.5 g,酵母浸出汁 1.0 g,NaCl 2.5 g,KH2PO3 0.5 g,硫酸鎂0.1 g,瓊脂10 g,去離子水500 mL,121 ℃下滅菌30 min。
1.3 纖維素酶活力測定方法
待測發(fā)酵液倒入離心管中,5 000 r/min離心 10 min,取上清液測定酶活。濾紙酶(FPA酶)活力測定:根據(jù)國際理論應(yīng)用化學(xué)協(xié)會(IUPAC)的方法測定[7]。CMC酶活力測定見參考文獻(xiàn)[8]。
1.4 菌株液體發(fā)酵產(chǎn)酶條件的優(yōu)化
1.4.1 發(fā)酵時間的確定 發(fā)酵培養(yǎng)基內(nèi)接入種子后,從12 h開始,每12 h取樣測定酶活,直到72 h。
1.4.2 接種量的確定 按照1%、2%、3%、4%、5%和6%的接種量將種子液接入發(fā)酵培養(yǎng)基中,于37 ℃、搖瓶轉(zhuǎn)速200 r/min的條件下發(fā)酵36 h后測酶活力。
1.4.3 培養(yǎng)基成分的優(yōu)化
1.4.3.1 碳源的優(yōu)化 (1)碳源種類的優(yōu)化:以羧甲基纖維素鈉(CMC-Na)、麩皮(過80目篩)、玉米粉、葡萄糖、蔗糖、淀粉和微晶纖維素作為唯一碳源(2%)配制培養(yǎng)基,接入種子后發(fā)酵36 h后測酶活力。(2)碳源濃度的優(yōu)化:在優(yōu)化碳源種類后,配制碳源濃度為0.5%、1%、1.5%、2%、2.5%、3%、35%和4%的發(fā)酵培養(yǎng)基,接入突變菌株發(fā)酵36 h后測酶活力。
1.4.3.2 氮源的優(yōu)化 (1)氮源種類的優(yōu)化:優(yōu)化碳源后,以蛋白胨、豆餅粉(過80目篩)、大豆蛋白粉、硫酸銨、硝酸鉀和尿素作為唯一氮源(0.5%)配制發(fā)酵培養(yǎng)基,接入種子后發(fā)酵36 h后測酶活力。(2)氮源濃度的優(yōu)化:在優(yōu)化氮源種類后,配制優(yōu)化氮源濃度為0.1%、0.3%、0.5%、0.7%、0.9%和11%發(fā)酵培養(yǎng)基,接入種子后發(fā)酵36 h后測酶活力。
1.4.3.3 初始pH值的優(yōu)化 分別配制pH值4.5、5.0、5.5、6.0、6.5、7.0、7.5、8.0碳氮源優(yōu)化后發(fā)酵培養(yǎng)基,接入突變菌株發(fā)酵36 h后測酶活力。
1.4.3.4 磷酸鹽濃度優(yōu)化 配制K2HPO3濃度為01%、0.5%、0.9%、1.3%和1.7%的上述條件優(yōu)化后的發(fā)酵培養(yǎng)基,接入突變菌株發(fā)酵36 h后測酶活力。
1.4.4 響應(yīng)面設(shè)計試驗 在上述優(yōu)化基礎(chǔ)上,根據(jù)Box-Behuken設(shè)計方法,采用minitab 16構(gòu)建3因素(碳源濃度、氮源濃度和磷酸鹽濃度)3水平響應(yīng)面分析[9-10]。
1.4.5 統(tǒng)計分析 利用SPASS軟件實現(xiàn)統(tǒng)計分析及差異顯著性檢驗[11]。
2 結(jié)果與分析
2.1 發(fā)酵時間的確定
接入種子液后,從發(fā)酵12 h開始每隔12 h測定FPA酶活和CMC酶活,直到發(fā)酵后72 h,結(jié)果如圖1所示。發(fā)酵時間為36 h時,菌株產(chǎn)FPA酶和CMC酶都表現(xiàn)出了比較高的酶活力,繼續(xù)發(fā)酵到48 h會有少許增加,之后逐漸下降。經(jīng)差異顯著性檢驗,發(fā)酵36 h酶活與發(fā)酵48 h酶活差異不顯著(P>0.05),而與其他發(fā)酵時間的酶活差異極顯著(P<0.01),確定36 h為最優(yōu)發(fā)酵時間。
2.2 接種量的確定
不同接種量下菌株發(fā)酵產(chǎn)酶結(jié)果見圖2。隨接種量提高,菌株產(chǎn)酶活力不斷提高。接種量大于4%時,酶活提高量變小。經(jīng)差異顯著性檢驗,4%、5%和6%這3種接種量條件下的酶活差異不顯著(P>0.05),綜合考查,選擇4%的接種量為優(yōu)化條件。
2.3 初始pH值的優(yōu)化
菌株DM-4在不同初始pH值條件下發(fā)酵產(chǎn)纖維素酶活力結(jié)果見圖3。試驗結(jié)果顯示,初始pH值為5.5~6.0時,菌株DM-4產(chǎn)酶活性最高。顯著性檢驗結(jié)果表明,初始pH值5.5條件下菌株所產(chǎn)酶活與初始pH值6.0條件下差異不顯著(P>005),而與其他初始pH值條件下差異極顯著(P<0.01),因此,確定最優(yōu)產(chǎn)酶初始pH值為5.5~6.0。
2.4 培養(yǎng)基組分的優(yōu)化
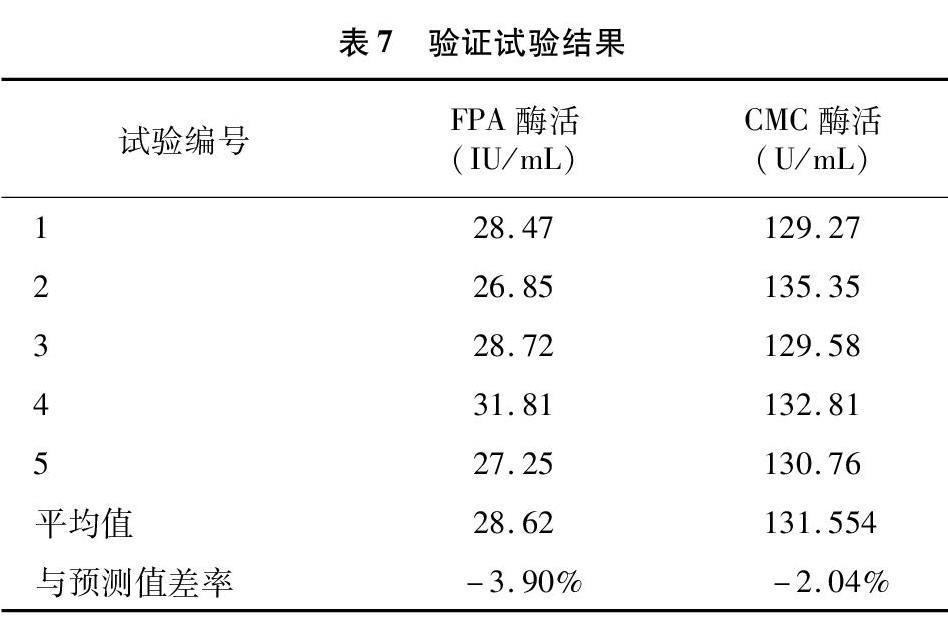
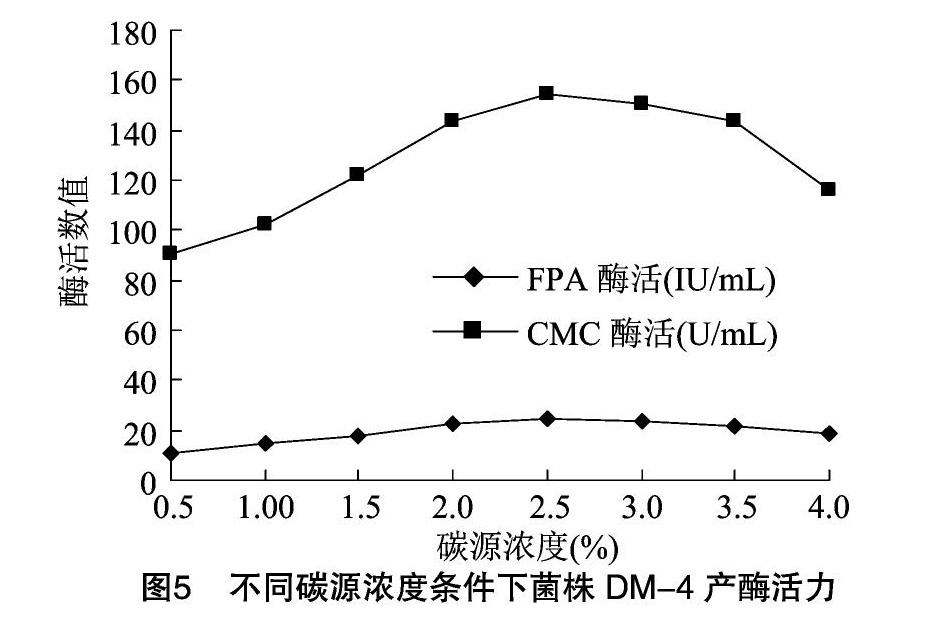
2.4.1 碳源的優(yōu)化 分別利用不同碳源作為唯一碳源配制培養(yǎng)基進(jìn)行發(fā)酵,結(jié)果見圖4。總體分析,含有纖維素物質(zhì)的碳源比不含纖維素物質(zhì)的碳源所產(chǎn)酶活高。其中以麩皮為唯一碳源時產(chǎn)酶能力最強(qiáng),與其他碳源比較差異極顯著(P<0.01),確定麩皮為最優(yōu)碳源。由圖5可知,當(dāng)麩皮濃度在 2.5% 以下時,隨著麩皮濃度升高,酶活升高,超過2.5%時,酶活反而下降。在碳源濃度為2.5%時菌株所產(chǎn)酶活與其他濃度下所產(chǎn)酶活比較具有顯著性差異。碳源濃度過低會造成營養(yǎng)不足,能源物質(zhì)少,細(xì)胞不能生成足夠的ATP滿足菌體的生長代謝,菌體生長不良,產(chǎn)酶活性低;而碳源濃度過高時,能源物質(zhì)充足,細(xì)胞生長快,同時發(fā)酵液黏度升高,溶氧不足,使細(xì)胞產(chǎn)酶量下降,降低活性。在本試驗條件下選擇麩皮濃度2.5%作為優(yōu)化碳源濃度。
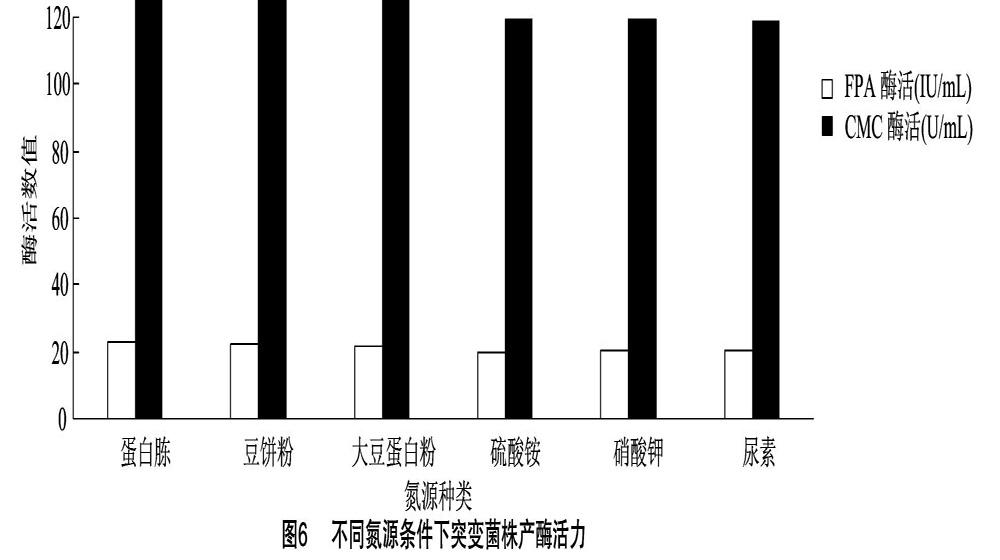
2.4.2 氮源的優(yōu)化 分別以不同物質(zhì)作為唯一氮源進(jìn)行發(fā)酵試驗,結(jié)果見圖6。總體而言,有機(jī)氮源產(chǎn)纖維酶酶活性要比無機(jī)氮源稍高。差異顯著性檢測表明,蛋白胨、豆餅粉和大豆蛋白粉作為氮源時,其酶活之間差異不顯著(P>005),而與硫酸銨、硝酸鉀和尿素作為氮源時的產(chǎn)酶活性差異極顯著(P<0.01)。由于試驗菌株是在酒醅中篩選出來的,經(jīng)常年不斷優(yōu)化,適應(yīng)了有機(jī)氮源的營養(yǎng),同時,有機(jī)氮源中所含的營養(yǎng),除氮源外還有其他營養(yǎng)成分,會促進(jìn)菌體產(chǎn)酶。綜合考慮,本試驗以蛋白胨作為最佳氮源。氮源濃度優(yōu)化試驗結(jié)果(圖7)顯示,隨氮源濃度增加,菌株產(chǎn)酶活力增加,當(dāng)?shù)礉舛葹?.9%時,菌株產(chǎn)酶活力最高,氮源濃度繼續(xù)提高至1.1%后,酶活力少許下降。氮源是細(xì)胞合成產(chǎn)物的重要物質(zhì),數(shù)量過少,產(chǎn)物合成不足,數(shù)量過多,有可能造成底物抑制,細(xì)胞合成酶的數(shù)量降低,從而使菌體產(chǎn)酶活性降低。通過0.9%濃度時的酶活力與其他氮源濃度下的酶活力差異性比較可知,氮源濃度0.9%時,F(xiàn)PA酶活和CMC酶活與其他濃度下的酶活具有顯著性差異(P<0.05),因此,選擇氮源濃度為0.9%作為優(yōu)化濃度。
2.4.3 磷酸鹽濃度的優(yōu)化 配制磷酸鹽濃度分別為0.1%、0.5%、0.9%、1.3%和1.7%的培養(yǎng)基,接入菌株后進(jìn)行發(fā)酵,36 h后測定纖維素酶活力,結(jié)果見圖8。當(dāng)磷酸鹽(KH2PO3)濃度為 0.5% 時,菌株所產(chǎn)酶活最高。濃度進(jìn)一步增加后,酶活反而下降。磷酸鹽濃度很小時,磷元素不足造成細(xì)胞代謝緩慢,磷酸鹽濃度過高,會引起細(xì)胞生長過快,導(dǎo)致營養(yǎng)物質(zhì)只合成細(xì)胞而不產(chǎn)酶。顯著性檢驗表明,在磷酸鹽濃度為0.5%的條件下,與其他濃度條件下的酶活差異極顯著(P<0.01),因此,確定磷酸鹽0.5%的濃度作為優(yōu)化結(jié)果。
2.3.5 響應(yīng)面試驗結(jié)果 選擇碳源(麩皮)濃度、氮源(蛋白胨)濃度和磷酸鹽(KH2PO3)濃度作為培養(yǎng)基組分優(yōu)化因子(分別為X1、X2、X3)以FPA酶(Y1)和CMC酶(Y2)活力分別作為響應(yīng)值設(shè)計試驗方案。試驗因子和水平見表1,試驗方案和結(jié)果見表2。
使用mintab 16進(jìn)行關(guān)于響應(yīng)值與因子麩皮濃度(X1)、蛋白胨濃度(X2)和磷酸鹽濃度(X3)的二次回歸分析,去掉不顯著因素,得到二次表達(dá)式:
Y1=29.81-X3-4.267 5X21-1.872 5X22-3.52X23-1.352 5X2X3;
Y2=134.14+2.355X1-1.383 8X3-6.913 7X21-3.091 2X22-5.166 3X23-1.837 5X2X3。
方程顯著性檢驗結(jié)果如表3至表6所示,2個回歸方程都具有顯著性(R21=0.936,R22=0.949,P<0.01),失擬項不顯著(P>0.05),剔除其他項后,保留的各項系數(shù)都具有顯著性(P<0.05)。根據(jù)顯著性檢驗結(jié)果可以得知,對于菌株產(chǎn)FPA酶活力來說,培養(yǎng)基中的麩皮濃度和蛋白胨濃度在二次水平上影響FPA酶的產(chǎn)生,磷酸鹽濃度在一次水平和二次水平上都對FPA酶的產(chǎn)生具有顯著性影響,麩皮濃度和蛋白濃度之間及麩皮與磷酸鹽濃度之間不具有交互作用,而蛋白胨和磷酸鹽之間對FPA酶的產(chǎn)生具有交互作用。對菌株所產(chǎn)CMC酶來說,麩皮濃度在一次水平上也會影響CMC酶活的生產(chǎn),其他與FPA酶一致,由此說明,菌株所產(chǎn)的CMC酶活力是FPA酶活力的重要組成部分。
利用mintab 16軟件繪出等值線圖及響應(yīng)面圖(圖9、圖10)。等值線的形狀反映因素間交互作用的大小,圖形傾斜表示交互作用顯著。結(jié)合方差分析和響應(yīng)面圖可知,蛋白質(zhì)濃度與磷酸鹽濃度交互作用具有顯著水平,而麩皮與蛋白胨、麩皮與磷酸鹽之間的交互作用不顯著。
FPA酶優(yōu)化后水平為:X1=0,X2=0.055 1,X3=-0.152 6;CMC酶優(yōu)化后因子水平為:X1=0.170 3,X2=0.037 8,X3=0.141 4。考慮2種酶取因子水平平均值:X1=0.085 15,X2=0.046 45,X3=-0.005 6。對應(yīng)的麩皮濃度為2.54%,蛋白胨為0.92%,磷酸鹽為0.497%(取0.50%)。在此條件下利用回歸表達(dá)式預(yù)測2種酶活力大小,F(xiàn)PA酶為29.78 IU/mL,CMC酶為134.29 U/mL。
2.4 優(yōu)化結(jié)果驗證
根據(jù)優(yōu)化結(jié)果,在麩皮濃度2.54%、蛋白胨濃度0.92%、磷酸鹽濃度0.50%、發(fā)酵初始pH值 5.5、接種量4%和發(fā)酵時間36 h的條件下對菌株進(jìn)行5次產(chǎn)酶試驗,檢測產(chǎn)酶活力,并與模型預(yù)測值比較,結(jié)果如表7所示。
驗證試驗顯示,在菌株所產(chǎn)纖維素酶活力與模型預(yù)測值接近,差率很小(<5%),由此可確定對菌株DM-4產(chǎn)酶條件的優(yōu)化模型是有效和可靠的。
3 結(jié)論
利用單因素試驗和響應(yīng)面試驗,確定產(chǎn)纖維酶菌株DM-4的產(chǎn)酶優(yōu)化條件為:發(fā)酵時間為36 h,接種量4%,培養(yǎng)基初始pH值5.5~6.0,培養(yǎng)基麩皮濃度2.54%,蛋白胨濃度0.92%,磷酸鹽濃度 0.5%。驗證試驗表明優(yōu)化模型是有效和可靠的。
參考文獻(xiàn):
[1]Peter M,Zarnea G,Adrina P,et al. Biodegradation and bioconversion of cellulose wastes using bacterialb and fungalcells immobilized hydrogels[J]. Resources Conservation and Recycling,1999,27(5):309-332.
[2]Rabinovich M L,Melnick M S,Bolobova A V. The structure and
mechanism of action of cellulolytic enzymes[J]. Biochemistry-Moscow,2002,67(8):850-871.
[3]Chinnarjan R,Thangiah N,Govindaswamy R V. Optimization of alkaline cellulase production by the marine-derived fungus Chaetomium sp. using agricultural and industrial wastes as substrates[J]. Botanica Marina,2010,53(3):275-282.
[4]Taechapoempol K,Sreethawong T,Rangsunvigit P,et al. Cellulase-Producing bacteria from Thai higher termites,Microcerotermes sp.:enzymatic activities and ionic liquid tolerance[J]. Applied Biochemistry and Biotechnology,2011,164(2):204-222.
[5]Deka D,Jawed M,Goyal A.Purification and characterization of an alkaline cellulase produced by Bacillus subtilis (AS3)[J]. Preparative Biochemistry & Biotechnology,2013,43(3):256-270.
[6]Choudhuryl B,Chauhan S,Singh S N,et al. Production of xylanase of Bacillus coagulans and its bleaching potential[J]. World Journal of Microbiology & Biotechnology,2006,22(3):283-288.
[7]Ghose T K. Measurement of cellulase activity[J]. Pure and Applied Chemistry,1987,59(2):257-268.
[8]Nwagu K E,Ominyi M C,Nwoba G E. Isolation,screening and measurement of amylase and cellulase activities of some microorganisms[J]. Continental Journal of Biological Sciences,2012,6(1):37.
[9]Wang G G. Adaptive response surface method using inherited Latin hypercube design points[J]. Journal of Mechanical Design,2003,125(2):210-220.
[10]Steinberg D M,Bursztyn D. Response surface methodology in biotechnology[J]. Quality Control & Applied Statistics,2011,22(2):109-110.
[11]Hoshino T,Hashimoto T,Shigemasu K. A bayesian significance test for the difference and linear combination of factor means between groups[J]. Japanese Journal of Educational Psychology,2001,49(1):31-40.李林紅,李孟涵. 農(nóng)產(chǎn)品圖文呈現(xiàn)與在線評論不一致性對銷量的影響——以蘋果為例[J].
